
Mesele Yihune
Department of Biology, Debre Markos University, P.O. Box 269, Debre Markos, Ethiopia
Afework Bekele
Department of Zoological Sciences, Addis Ababa University, P.O. Box 1176, Addis Ababa, Ethiopia
International Journal of Zoological Research
Year: 2012 | Volume: 8 | Issue: 4 | Page No.: 137-149







ABSTRACT
Species composition, distribution and abundance of rodent community were described in the Simien Mountains National Park. The study was carried out between June, 2011 and May, 2012 during the wet and dry seasons. Six species of rodents (Lophuromys flavopunctatus, Arvicanthis abyssinicus, Stenocephalemys griseicauda, Otomys typus, Rhabdomys pumilio and Tachyoryctes splendens) and two species of insectivores (Crocidura baileyi and Crocidura thalia) were recorded. R. pumilio is the newly recorded species. The distribution of rodents and insectivores was significantly differed (χ2 = 132.3, df = 4, p<0.001) across studied habitats. There was also variation in species abundance in all habitats. S. griseicauda is the highest (45.8%) species recorded in all habitat types followed by A. abyssinicus (28%). Large numbers of individuals were trapped in Helichrysum-Festuca habitat. The mean number ± SE of individuals caught during the day and night were 3.18±0.05 and 1.91±0.72, respectively. However, there was no statistically significant difference (χ2 = 1.93, df = 1, p>0.05) between the overall number of species recorded during the day and night time. There was variation (χ2 = 25, df = 1, p<0.001) in the number of rodents and insectivores trapped during the wet and dry seasons. The average number ±SE of rodents and insectivores caught during the wet and dry seasons were 2.77±0.07 and 2.28±0.07, respectively. Human interference and grazing pressure influenced the rodent community in the study area. This should be curtailed so as to increase the species composition and abundance of rodents. Hence, this will help to sustain their ecological importance in the study area.
PDF Abstract XML References Citation
Received: July 25, 2012;
Accepted: September 06, 2012;
Published: December 17, 2012
How to cite this article
Mesele Yihune and Afework Bekele, 2012. Diversity, Distribution and Abundance of Rodent Community in the Afro-alpine Habitats of the Simien Mountains National Park, Ethiopia. International Journal of Zoological Research, 8: 137-149.
DOI: 10.3923/ijzr.2012.137.149
URL: https://scialert.net/abstract/?doi=ijzr.2012.137.149
DOI: 10.3923/ijzr.2012.137.149
URL: https://scialert.net/abstract/?doi=ijzr.2012.137.149
INTRODUCTION
Rodents are the most diverse group of mammals (Vaughan et al., 2000). They show great diversity in their ecology, morphology, physiology, behaviour and life history strategies (Nedbal et al., 1996). They are well adapted to a wide range of environments (Nowak, 1999). Rodents are also the most diverse and abundant among mammals in Africa (Bekele, 1996). There are 84 species of rodents in Ethiopia (Bekele and Leirs, 1997). Of the total rodent species of the country, 21% are endemic. The unique topography and wide range of climatic conditions are reasons for diverse biological resources in Ethiopia (Hillman, 1993). Rodents are better adapted to the afro-alpine environment than large mammals due to their ability to avoid temperature fluctuations by hiding themselves in their burrows (Marino, 2003). The presence of large body size is another adaptation for the afro-alpine rodents. Most (60%) of the rodent fauna of the country is confined to the highlands and there are at least 14 endemic species of rodents, all from the northwestern, central and southeastern highland plateaux (Yalden and Largen, 1992). Six endemic species belong to the afro-alpine moorland above 3000 m.a.s.l. (Dendromus lovati, Arvicanthis blicki, Stenocephalemys griseicauda, Stenocephalemys albipes, Praomys ruppi and Mus mahomet), four endemic species occur in mountain grassland (Tachyoryctes macrocephalus, Stenocephalemys albocaudata, Lophuromys melanonyx and Megadendromus nikolausi) and the other four endemics occur in mountain forest areas. Furthermore, 16 species of rodents which are not endemic live in the highlands. However, only some species like Tachyoryctes splendens, Otomys typus, Lophuromys flavopunctatus and Arvicanthis abyssinicus survive above 3000 m.a.s.l. (Yalden and Largen, 1992). Usually, night frosts, diurnal temperature fluctuations and strong irradiation are common in the afro-alpine areas. However, rodents can avoid extreme cold by staying underground so that they are inherently well adapted as the dominant herbivores of the afro-alpine ecosystem (Marino, 2003).
Rodents show habitat preferences and this mainly is dependent upon the vegetation type and life history strategies (Fitzherbert et al., 2007). There are rodents that can only survive in a narrow altitudinal range whereas others are altitude generalists (Mukinzi et al., 2005). Their distribution and abundance are influenced by environmental factors such as the nature and density of vegetation, climatic conditions, disease, predation and habitat exploitation by humans (Johnson and Horn, 2008). Absence of sufficient food and ground cover largely determine the number of individual rodents in a certain area. The loss of ground vegetation leads to loss of cover and food supply for small mammals thereby decreasing rodent diversity but increasing predation risk (Hoffmann and Zeller, 2005). On the other hand, habitat structure and predation risk affect species composition in various habitat types (Massawe et al., 2007). Bushy vegetation provides safe sites for germination and growth of herbaceous vegetation, hence enhancing the diversity of rodent food resources (Kerley et al., 2004). The density of rodents enhances in habitats such as bushland, grassland and forest in the presence of ample food (Lentic and Dickman, 2005). The diversity of species can be resulted from habitat variability due to the supply of different resources (Cramer and Willig, 2002).
In terms of evolution, rodents are quite young where the populations retain unexploited source of genetic variability (Vaughan et al., 2000). The number of rodents and their dynamics usually depend upon extensive vegetation cover (Hansson, 1999). Rodents form vital components of ecosystems (Avenant and Cavallini, 2007). They interact extensively with their environments and their activities have beneficial effects on other organisms in different ecosystems. They are important dietary components for many carnivorous small mammal, raptors and reptiles (Davies, 2002). They are vital food for the survival of the Ethiopian wolf (Canis simensis) (Sillero-Zubiri et al., 1995). On the other hand, they are host of disease causing parasites such that wild rodents can host Schistosoma mansoni (El-Naggar et al., 2011). Ecological studies of the rodent communities in Ethiopia are important due to their high levels of endemism. In addition, the Ethiopian wolf heavily depends on rodents for its survival (Malcolm and Sillero-Zubiri, 1997). Thus, studies of the rodent community are important to assess habitat quality for wolves and as a predictor wolf density. Hence, the present study covers species composition and abundance.
A few studies of rodent biology have been conducted in the southeastern highlands of the Bale Mountains (Lavrenchenko et al., 1995) and only few studies were conducted in the northwestern highlands of Simien Mountains (Hurni, 1986). In the Simien Mountains National Park, small mammals are of special interest for different reasons. Several species are endemic and occurring only in the highland areas of Ethiopia (Yalden and Largen, 1992). Therefore, the present study investigates diversity, distribution, abundance, habitat association and activity patters of rodents in the afro-alpine habitats of the Simien Mountains National Park.
MATERIALS AND METHODS
The study area: The Simien Mountains National Park (SMNP) represents one of the most scenic natural areas in the world. It is a part of the Simien Mountains located between 38000'-38012'E and 13012'-13019’N (Fig. 1). Originally, the area of SMNP used to be 136 km2 but recently the area has been extended to 412 km2. It contains broad undulating plateau and grassy plains. The area includes Mt. Ras Dejen which is the highest mountain in Ethiopia (4,543 m.a.s.l.). The massif was formed 25 million years ago. In the process, the igneous basalts have been eroded to form precipitous cliffs and deep gorges. Some cliffs reach 1,500 m.a.s.l. and extend horizontally long distances. The plateau is surrounded on the south and northeast by deep valleys of Tekeze River (Hurni and Ludi, 2000).
SMNP was nationally gazetted in 1969. Then, it was designated as a World Heritage Site in 1978 due to the presence of high number of endemic species, unique bio-physical features and its international significance. However, in 1996 it was listed as World Heritage in Danger. This is because of the declining number of Walia ibex (Capra walie) population, agricultural encroachment, loss of biodiversity and impact of road construction (Falch and Keiner, 2000). However, currently the wildlife is increasing from time to time due to effective and community based conservation. The climate of SMNP varies from area to area. Generally, the highlands have a relatively low temperature. They are cold in the early mornings of the dry season and receive annual rainfall of 1500 mm which occurs mainly from May to October (Hurni, 1986). The temperature of the area shows high diurnal variability. The annual minimum temperature ranges from -4-2.5°C in January and maximum temperature from 11-18°C in April.
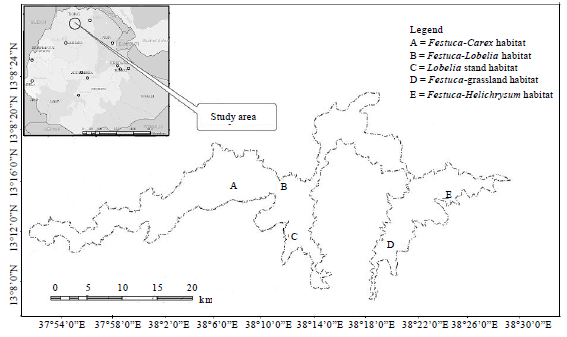 | |
| Fig. 1: | Map of Simien Mountains National Park and the study area |
The SMNP is part of the afro-alpine centre of plant diversity with high level of endemism. The park preserves a representative part of the Ethiopian Tropical seasonal highland biome and contains vegetation characteristics of each (Debonnet et al., 2006). It consists of a mixture of Afro-alpine woods, heath forest, high mountain vegetation, montane savanna and moorland (Hurni and Ludi, 2000). In the SMNP, 57 tree species and herbaceous plants have been recorded. There are over 20 endemic plant species within and in the buffer zone of the SMNP (Falch and Keiner, 2000). Numerous species of mammals and birds exist in SMNP (Hurni, 1986). In the SMNP, there are 22 large mammals, 13 small mammals and 180 bird species.
Trapping: A permanent 4900 m2 live trapping grid was established at five randomly selected habitat types (Festuca-Lobelia, Lobelia stand, Festuca grassland, Festuca-Carex and Festuca-Helichrysum habitats) for three consecutive days and nights. In each trapping site, a standard square (seven rows by seven columns) trapping grid was established during wet and dry seasons (Linzey and Kesner, 1997). A total of 49 Sherman traps (5.5x6.5x16 cm) were set per grid at every 10 m interval between points during both seasons. The traps were baited with peanut butter and barley flour. Traps were covered with leaves and grasses to prevent from cold weather consequently death of rodents and to avoid the damage of traps by other wild animals. The traps were checked twice a day early morning (between 06:00 and 07:00 h) and late afternoon (between 17:00 and 18:00 h). Animals caught from 06:00-18:00 h were recorded as day trappings and those caught from 18:00-06:00 h the next morning as night trappings. Each trapped animal was captured alive, identified, marked by toe clipping and released back to the site from where it was trapped (Linzey and Kesner, 1997; Clausnitzer, 2003). Information such as weight, sex, approximate age (juvenile, sub-adult and adult) based on their weight and coat colour (Bekele, 1996) and reproductive condition (for females: Imperforate or perforated vagina and for males: Scrotal or abdominal testes) were recorded. Furthermore, tail length, ear length, hind foot length and incisor type at lower jaw (grooved or ungrooved) was also recorded for identification purposes.
Statistical analysis: Data were analyzed using SPSS statistical software package version 17 and Microsoft Excel. Chi-square test was performed to test significant variation of the number of individuals caught across habitat types, seasons and time. One-way ANOVA was also used to test mean differences of seasonal distribution and variation in age classes.
RESULTS
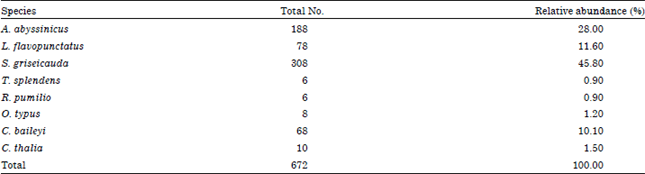
Diversity and abundance: A total of 672 individual rodents (order Rodentia: Family Muridae and Rhizomyidae) and insectivores (order Insectivora) were captured in five habitat types during wet and dry seasons. Rodent species which were identified and recorded were: Lophuromys flavopunctatus, Arvicanthis abyssinicus, Stenocephalemys griseicauda, Tachyoryctes splendens, Otomys typus and Rhabdomys pumilio. Two insectivore species, Crocidura baileyi and Crocidura thalia were also recorded. Rhabdomys pumilio is a new record for Simien Mountains National Park. The relative abundance of caught rodent and insectivore species include: A. abyssinicus (28%), L. flavopunctatus (11.6%), S. griseicauda (45.8%), T. splendens (0.9%), R. pumilio (0.9%), O. typus (1.2%), C. baileyi (10.1%) and C. thalia (1.5%) (Table 1).
Distribution, habitat association and activity patterns: The distribution of rodents and insectivores significantly differed (χ2 = 132.3, df = 4, p<0.001) across studied habitats throughout the study period. Variation in species composition and abundance was observed across all habitats. A. abyssinicus, S. griseicauda and C. baileyi were widely distributed in all habitats. S. griseicauda (n = 308) is the largest recorded rodent species from all habitat types followed by A. abyssinicus (n = 188). On the other hand, T. splendens (n = 6) and R. pumilio (n = 6) were the least recorded species in the study area. T. splendens was recorded form Lobelia stand and Festuca-Carex habitat whereas R. pumilio was only recorded from Festuca-Lobelia habitat (Table 2).
Most (33.9%) of the species were trapped from Helichrysum-Festuca habitat followed by Festuca grassland habitat (27.1%). Furthermore, 14.1 and 12.5% of the species were trapped from Festuca-Carex and Festuca-Lobelia habitats, respectively. The least number of species was recorded from Lobelia stand habitat (12.2%) (Fig. 2).
The number of different species of rodents and insectivores caught across all five habitat types had been further sub-divided in to those caught during the day and night. A total of 354 (52.7%) individuals were captured during the night time and 318 (47.3%) during day time. The mean number ±SE of individuals caught was 3.18±0.05 during the night and 1.91±0.72 during the day time. However, there was no statistically significant difference (χ2 = 1.93, df = 1, p>0.05) between the overall number of species recorded during the day and night time (Fig. 3).
There was a significant difference (χ2 = 306.5, df = 7, p<0.001) in the proportion of each species trapped during the day and night time. A. abyssinicus and L. flavopunctatus were primarily diurnal whereas S. griseicauda, C. baileyi, C. thalia and R. pumilio were nocturnal. However, equal numbers of O. typus individuals were trapped during day and night time.
| Table 1: | Number of rodent and insectivore species and their relative abundance in the study area |
 | |
| Table 2: | Number of rodent and insectivore species recorded in different habitats |
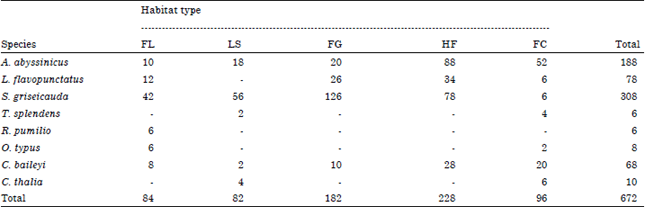 | |
| FL: Festuca-Lobelia, LS: Lobelia stand, FG: Festuca-grassland, HF: Helichrysum-Festuca, FC: Festuca-Carex grassland | |
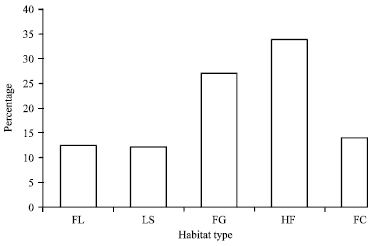 | |
| Fig. 2: | Percentage of species trapped in different habitats, FL: Festuca-Lobelia, LS: Lobelia stand, FG: Festuca-grassland, HF: Helichrysum-Festuca and FC: Festuca-Carex grassland |
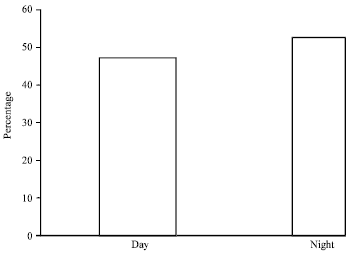 | |
| Fig. 3: | Percentage of rodents and insectivores caught during day and night time |
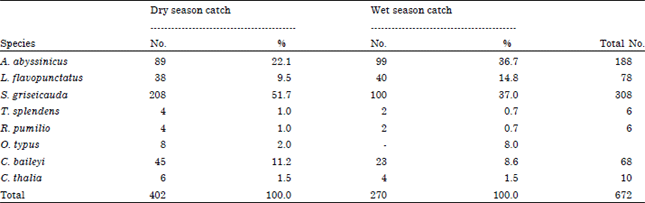
Seasonal variation: Species caught significantly differed (χ2 = 25, df = 1, p<0.001) in their number during the wet and dry seasons. During the dry season, 402 individuals were trapped whereas during the wet season 270 individuals were caught. S. griseicauda was the highest number of species caught during the dry (n = 208) and wet (n = 100) seasons followed by A. abyssinicus during the dry (n = 89) and wet (n = 99) seasons. All species were caught during both seasons except O. typus. O. typus was not trapped during the wet season (Table 3). The mean number ±SE of rodents and insectivores caught were 2.77±0.07 and 2.28±0.07 during the dry and wet seasons, respectively.
The distribution of rodents and insectivores significantly differed (F670 1 = 30.49, p<0.001) across studied habitats during wet and dry seasons. The highest number of individuals was caught in Helichrysum-Festuca habitat during both the wet (n = 77, 28.5%) and dry (n = 151, 37.6%) seasons (Fig. 4).
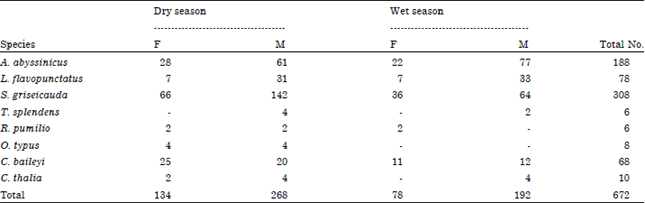
Sex and age structure: The distribution of sex is significantly different among individuals (χ2 = 91.5, df = p<0.001) and across seasons (χ2 = 25.9, df = 1, p<0.001). Most (68.5%) of the trapped individuals were males and the remaining 31.5% were females. S. griseicauda consisted of the highest number of female (n = 66) and male (n = 142) individual in the dry season. On the other hand, the highest number of caught species was male A. abyssinicus (n = 77) followed by male S. griseicauda (n = 64) during the wet season. Female individuals of T. splendens were not caught during both seasons (Table 4).
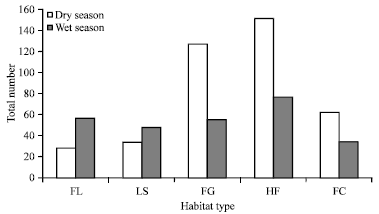 | |
| Fig. 4: | Total number of rodents and insectivores across all habitats caught during wet and dry seasons, FL: Festuca-Lobelia, LS: Lobelia stand, FG: Festuca-grassland, HF: Helichrysum-Festuca and FC: Festuca-Carex grassland |
| Table 3: | Number and relative percentage of rodents and insectivores caught during the wet and dry seasons |
 | |
| Table 4: | Sex distribution of rodents and insectivores across seasons |
 | |
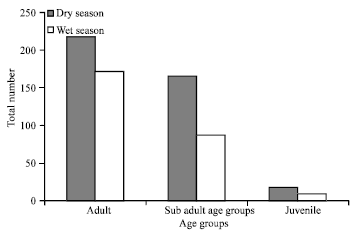 | |
| Fig. 5: | Age distribution of trapped rodents and insectivores during the dry and wet seasons |
Of the total number of species caught, 58% (n = 390) were adults, 37.8% (n = 254) were sub-adults and 4.2% (n = 28) were juveniles. There is a significant variation (F 670 1 = 5.12, p<0.05) among species caught in terms of their age class across seasons. Adults were largely caught in both dry (n = 218) and wet (n = 172) seasons. In contrast, juveniles were the least caught groups during the dry (n = 18) and wet (n = 10) seasons (Fig. 5).
DISCUSSION
Diversity and abundance: Different rodent species are confined in the highlands of Ethiopia (Yalden and Largen, 1992). According to the present study, six species of rodents (A. abyssinicus-unstriped grass rat, L. flavopunctatus-harsh-furred rat, S. griseicauda-narrow headed rat, O. typus-swamp rat, T. splendens-common mole rat and R. pumilio-striped grass mouse) and two species of insectivores (C. baileyi and C. thalia-white toothed shrews) have been recorded in the SMNP. However (Guttinger et al., 1998) recorded six species of rodents (A. abyssinicus, L. flavopunctatus, S. griseicauda, T. splendens, O. typus and Pelomys harringtoni) and one species of shrew (C. baileyi). Pelomys harringtoni had not been recorded in our study whereas R. pumilio was not recorded before in the SMNP. On the other hand (Busby et al., 2006) recorded three species of rodents (A. abyssinicus, L. flavopunctatus and O. typus) and one insectivore species (C. thalia). S. griseicauda is the most abundant (45.8%) rodent species in the study area. Similarly, a study conducted in Guassa area of Menz indicated that S. griseicauda was the most abundant species in all habitat types (Tefera, 2001).
Distribution, habitat association and activity pattern: The distribution of rodents varied across habitats throughout the study area. Although, S. griseicauda was distributed in all habitat types, it was highly abundant in Festuca grassland habitat. A similar result was observed in Guassa area of Menz (Tefera, 2001). A. abyssinicus is one of the endemic species of Ethiopian plateau (Yalden et al., 1976). A. abyssinicus was distributed in all habitat types but its abundance was high in Helichrysum-Festuca habitat. According to Yalden et al. (1976), L. flavopunctatus is a widely distributed species in Ethiopia. Kityo and Clausnitzer (2001) added that the species has a wide range of distribution along the different montane and highland habitats of East Africa. But, this species has moderate ecological tolerance confined only to three or four habitats of forests (Happold, 1974). L. flavopunctatus is also highly abundant in Helichrysum-Festuca habitat. However, it was not trapped in Lobelia stand habitat. Lophuromys spp. shows a habitat preference of long grass cover and is known to be intolerant of grazing (Marino, 2003). O. typus was trapped from Festuca-Lobelia and Festuca-Carex habitats. It was not caught in any other habitats. Similarly, O. typus, was common in Festuca, Lobelia, Carex habitat of the Simien Mountains National Park (Guttinger et al., 1998).
The present study added R. pumilio as the newly recorded species of rodents in the SMNP although, some scientists reported as Rhabdomys is missing in Ethiopia (Kityo and Clausnitzer, 2001). It was caught only in Festuca-Lobelia habitat during day time. R. pumilio cannot cope with hot climatic condition so it is restricted to high altitude area in East Africa (Delany and Happold, 1979). This restriction to the grassy afro-alpine area is related to its adaptive physiology (Kityo and Clausnitzer, 2001). Generally, the distribution of rodent species in the study area was presumably related to vegetation structure, availability of food and human and livestock impact in the study area. The grazing by livestock had a significant impact in the rodent community. Grazing has a number of detrimental effects which directly impact on rodent communities including food competition, trampling of rodent holes, reduced vegetation height and associated reductions in predation cover (Guttinger et al., 1998).
Habitat association is important to determine how various species respond to environmental heterogeneity (Martin, 1998). Habitat selection may have evolved as a result of past competitive effects but, only weak interspecific competition perhaps necessary to maintain habitat selection. Hence, the coexistence of species can be determined by resource availability and by means of which species utilizes which habitat and interact with others (Schoener, 1983). The abundance of rodents varied across all habitat types in the present study. There was high number of species in Helichrysum-Festuca followed by Festuca grassland habitats. This is probably because such habitats are good in providing shelter due to their dense cover. The least number of rodents was captured in Lobelia stand habitat. This may be related to the poor cover of the habitat exposing rodents to predators and other factors that decrease their population (Iyawe, 1988) indicated that habitat selection of different species of small mammals mainly relies on the vegetation of the given habitat. This idea is also supported by Taylor and Green (1976) where cover removal reduced most species of rodent population. Whenever there is enough cover and adequate food, the abundance of rodent population is high.
There was no statistically significant difference (p>0.05) between the overall number of individuals caught during the day and night time. However, there was a significant difference (p<0.001) in the proportion of each species trapped during the day and night time. Although the number of rodents trapped was higher during the night time than day time, the largest proportion of caught species was S. griseicauda (n = 256). Because of this reason, most rodents caught during the present study were diurnal. Similarly, a study conducted in Bale Mountains National Park (Sillero-Zubiri et al., 1995) and Guassa area of Menz (Tefera, 2001) also endorsed this idea in that most rodents in afro-alpine habitats show a diurnal activity pattern. The extreme low temperatures experienced at high altitude could impose limitation on the activity patterns of rodents. According to the present study, A. abyssinicus and L. flavopunctatus are diurnal rodents although some individuals are caught during night time. On the other hand, S. griseicauda is strictly nocturnal as very few individuals are trapped during the day time. Similar result was recorded in Guassa area of Menz (Tefera, 2001). According to Tefera (2001), thermoregulatory adaptation of S. griseicauda could be achieved by lowering its metabolic rate. In addition to this, environmental constraints, competition and predation may be crucial in determining the structure of rodent community.
Seasonal distribution: In the present study, there was variation (p<0.001) in the number of rodents and insectivores caught during the wet and dry seasons (Table 3). All the species trapped in this study were included during the dry season. However, O. typus was not trapped during the wet season. This variation may be caused by seasonal variation in vegetation structure, ground cover and other related environmental variables. Decreasing in food resources and vegetation cover mainly resulted from variation in habitat structures (Juch, 2000). Hence, the overall species composition of small mammals is affected. Habitat changes might be brought about by different factors. Joubert and Ryan (1999) revealed that diversity, distribution and abundance of small mammals are affected by wild ungulates and livestock due to over grazing. This might cause seasonal variation in the existence of a species in a given habitat. The largest proportion of individuals trapped in Helichrysum-Festuca habitat during both wet (28.5%) and dry (37.6%) seasons (Fig. 4). This is probably because such habitat provides a good shelter for them.
Sex and age structure: The distribution of sex among individuals trapped significantly differed (p<0.001). In this study, 68.5% of the total caught individuals were males whereas the remaining 31.5% were females. Nearly similar results were reported by Shenkut et al. (2006) in Alleltu Woreda, Ethiopia. Furthermore, this result is supported by the study conducted in southwest Ethiopia (Bayessa, 2010). The significant variation in the proportion of female and male population presumably was the result of differences in the mobility of males. Males generally make wider field excursion than females as reported by Odhiambo and Oguge (2003) in Kenyan Rift Valley. On the other hand, the responsibility of females in nursing their litters might have hindered their movement from one area to the other, consequently limiting the chance of entering the traps.
All age groups (adults, sub-adults and juveniles) were included in all trapping sessions. However, there was a significant variation (p<0.05) among individuals caught in terms of their age groups across seasons. Most (58%) of the individuals caught were adults. The least caught individuals were juveniles (4.2%). This could probably be due to the presence of large home ranges for adults and sub-adults to alleviate the cost of competition with juveniles. This is supported by (Gebresilassie et al., 2006) in that adult and sub-adult individuals have wider home range than young rodents. The capture rates of all age groups were higher during the dry season than during the wet season. This is because, during the dry season there is higher movement of rodents outside their burrow than during the wet season.
The Simien Mountains National Park is largely affected by human population pressure and livestock impact. This determines the rodent population in the study area. In addition, rodent populations are also affected by climatic factors according to the previous studies. Rodents have ecological importance in the study area. One of their ecological importance is that they are used as the main prey source for the Ethiopian wolf. Thus, affecting the population of rodents can determine the fate of Ethiopian wolf population in the area. Therefore, the effect of livestock grazing and humans on the rodent population in the Park should be curtailed. The present study is only conducted in the afro-alpine habitats of the Simien Mountains National Park. Hence further investigation need to be carried out in the lowland areas of the Park as well to get a complete picture.
ACKNOWLEDGMENTS
We sincerely thank Addis Ababa University for funding this study. We are also grateful to the Ethiopian Wildlife Conservation Authority for providing permit.
REFERENCES
- Bekele, A. and H. Leirs, 1997. Population ecology of rodents of maize fields and grassland in central Ethiopia. Belg. J. Zool., 127: 39-48.
Direct Link - Bekele, A., 1996. Population dynamics of the Ethiopian endemic rodent, Praomys albipes in the Menagesha State Forest. J. Zool., 238: 1-12.
CrossRef - Cramer, M.J. and M.R. Willig, 2002. Habitat heterogeneity, habitat associations and rodent species diversity in a Sand-Shinnery-Oak Landscape. J. Mammal., 83: 743-753.
Direct Link - El-Naggar, S.A., I.M. Al-Sharkawi and G.A. Madkour, 2011. Susceptibility of some wild rodents widely distributed in Egyptian Foci to Schistosoma mansoni infection under laboratory conditions. Int. J. Zool. Res., 7: 358-368.
CrossRefDirect Link - Fitzherbert, E., T. Gardner, T. Caro and P. Jenkins, 2007. Habitat preferences of small mammals in the Katavi ecosystem of western Tanzania. Afr. J. Ecol., 45: 249-257.
CrossRefDirect Link - Gebresilassie, W., A. Bekele, G. Belay and M. Balakrishnan, 2006. Home range and reproduction of rodents in Maynugus irrigation field, northern Ethiopia. SINET Ethiopia J. Sci., 29: 57-62.
Direct Link - Guttinger, R., L. Leumann, M. Getahun, J. Simmen and M. Wust, 1998. The Actual Situation of Several Small Mammal Species in the Park Area. In: A Survey on the Flora and Fauna of the Simien Mountains National Park, Ethiopia, Nievergelt, B., T. Good and R. Guttinger (Eds.). EWNHS and Group for Wildlife and Conservation Biology, University of Zurich, Switzerland, pp: 64-73.
- Hansson, L., 1999. Intraspecific variation in dynamics: Small rodents between food and predation in changing landscapes. Oikos, 86: 159-169.
Direct Link - Iyawe, J.G., 1988. Distribution of small rodents and shrews in a lowland rain forest zone of Nigeria, with observations on their reproductive biology. Afr. J. Ecol., 26: 189-195.
CrossRef - Johnson, M.D. and C.M. Horn, 2008. Effects of rotational grazing on rodents and raptors in a Coastal Grassland. Western N. Amer. Nat., 68: 444-452.
Direct Link - Juch, P., 2000. Habitat preference of small mammals on Mount Elgon, Uganda. Bull. Soc. Nat. Luxemb., 100: 83-96.
Direct Link - Kerley, G.I.H., W.G. Whitford and F.R. Kay, 2004. Effects of pocket gophers on desert soils and vegetation. J. Arid Environ., 58: 155-166.
Direct Link - Kityo, R. and V. Clausnitzer, 2001. Altitudinal distribution of rodents (Muridae and Gliridae) on Mt Elgon, Uganda. Trop. Zool., 14: 95-118.
Direct Link - Lentic, M. and C.R. Dickman, 2005. The responses of small mammals to patches regenerating after fire and rainfall in the Simpson Desert, central Australia. Aust. Ecol., 30: 24-39.
CrossRef - Linzey, A.V. and M.H. Kesner, 1997. Small mammals of a woodland savannah ecosystem in Zimbabwe. I. Density and habitat occupancy patterns. J. Zool. Lond., 243: 137-152.
CrossRef - Martin, T.E., 1998. Are microhabitat preference of coexisting species under selection and adaptive? Ecology, 79: 656-670.
Direct Link - Massawe, A.W., W. Rwamugira, H. Leirs, R.H. Makundi and L.S. Mulungu, 2007. Do farming practices influence population dynamics of rodents: A case study of the multimammate field rats, Mastomys natalensis, in Tanzania. Afr. J. Ecol., 45: 293-301.
Direct Link - Mukinzi, I., P.G.B. Katuala, J. Kennis, M. Gambalemoke and N. Kadange et al., 2005. Preliminary data on the biodiversity of rodents and insectivores (Mammalia) in the periphery of Kisangani (D.R. Congo). Belg. J. Zool., 135: 133-140.
Direct Link - Odhiambo, C.O. and N.O. Oguge, 2003. Patterns of Rodent Pest Distribution in Maize Cropping System in the Kenyan Rift Valley. In: Rats, Mice and People: Rodent Biology and Management, Singleton, G.R., L.A. Hinds, C.J. Krebs and D.M. Spratt (Eds.). Australian Centre for International Agricultural Research, Canberra, Australia pp: 217-220.
- Schoener, T.W., 1983. Field experiments on interspecific competition. Am. Nat., 122: 240-285.
Direct Link - Sillero-Zubiri, C., F.M. Tattersall and D.W. Macdonald, 1995. Bale Mountains rodent communities and their relevance to the Ethiopian wolf (Canis Simensis). Afr. J. Ecol., 33: 301-320.
CrossRef - Taylor, K.D. and M.G. Green, 1976. The influences of rainfall on diet and reproduction in four African rodent species. J. Zool., 180: 367-389.
CrossRef - Yalden, D.W. and M.J. Largen, 1992. The endemic mammals of Ethiopia. Mammal Rev., 22: 115-150.
CrossRefDirect Link