
S.A. Omer
Zoological Society of London, Conservation Programmes, Regent�s Park, London, NW1 4RY, United Kingdom
A. Apio
Department of Wildlife Management and Nature Conservation, Faculty of Veterinary Medicine, Umutara Polytechnic, P.O. Box 57, Nyagatare, Rwanda
T. Wronski
King Khalid Wildlife Research Centre, Saudi Wildlife Commission, P.O. Box 61681, Riyadh 11575, Kingdom of Saudi Arabia
O.B. Mohammed
King Khalid Wildlife Research Centre, Saudi Wildlife Commission, P.O. Box 61681, Riyadh 11575, Kingdom of Saudi Arabia
International Journal of Zoological Research
Year: 2011 | Volume: 7 | Issue: 1 | Page No.: 85-92







ABSTRACT
The aim of this study was to describe a new species of Eimeria from Farasan gazelle which has not been described before. Faecal examination of 24 Farasan gazelles, Gazella gazelle farasani collected on Frasan Kebir (Farasan Islands, Saudi Arabia) yielded oocysts of an undescribed coccidian parasite, Eimeria farasanii n. sp. Eleven out of the 24 faecal samples (45.8%) were infected with the parasite. Sporulated oocysts of Eimeria farasanii n. sp., are ellipsoidal, flattened at micropylar end, measured 20.8±1.8x19.5±1.6 μm (19-25x18-23 μm), length/width ratio 1.07±0.05 (1.04-1.15), with smooth, double-layered oocyst wall, the inner yellow, the outer bluish green, with micropyle 3.6 μm but without a micropylar cap. Sporocyst elongate, measured 8.2±1.1x3.2±0.7 μm (7-10x2-5 μm) length/width ratio 2.6±0.5 (2-4). Stieda body present, substieda body absent. Sporocyst residuum present, consists of diffuse, coarse, refractile granules. Sporozoites elongate, measured 6.5±1.2 μm (6-8 μm), each with a small refractile body at wide end. Sporulation time 7-8 days at 25±2°C. In the present investigation Eimeria farasanii n. sp., was described from Farasan gazelle based on the morphological differences with previously described species from the genus Gazella.
PDF Abstract XML References Citation
Received: March 12, 2010;
Accepted: May 20, 2010;
Published: August 21, 2010
How to cite this article
S.A. Omer, A. Apio, T. Wronski and O.B. Mohammed, 2011. A New Coccidian Parasite (Eimeria farasanii n. sp.) Indicates Parasite-Host Specificity in Endemic Farasan Gazelle. International Journal of Zoological Research, 7: 85-92.
DOI: 10.3923/ijzr.2011.85.92
URL: https://scialert.net/abstract/?doi=ijzr.2011.85.92
DOI: 10.3923/ijzr.2011.85.92
URL: https://scialert.net/abstract/?doi=ijzr.2011.85.92
INTRODUCTION
Eimeria infections are common in both domestic and wild artiodactyls in Saudi Arabia (Ghandour, 1988; Alyousif et al., 1992; Kasim and Al-Shawa, 1985a, b; Mohammed, 1997; Toulah, 2007). Several species were reported from camels (Kawashmeh and El-Bihari, 1983; Kasim et al., 1985; Hussein et al., 1987), sheep (Kasim and Al-Shawa, 1985a; Toulah, 2007), cattle (Kasim and Al-Shawa, 1985b), the Arabian oryx (Kasim and Al-Shawa, 1988), the Arabian sand gazelle (Hussein and Mohammed, 1992) and the Mountain gazelle (Mohammed and Hussein, 1992). Antelopes indigenous to Saudi Arabia include the Arabian oryx (Oryx leucoryx), the Sand gazelle (Gazella subgutturosa marica) and the Mountain gazelle (Gazella gazella ssp.). The latter species was reintroduced from a captive stock held at King Khalid Wildlife Research Centre (KKWRC) of the Saudi Wildlife Commission (SWC) into two protected areas in the Kingdom, i.e., Ibex Reserve and Uruq Bani Ma-Arid (Dunham, 1995, 1997; Dunham et al., 1993). Other populations are persisting in small numbers in several remote areas along the Mountain ranges of western Saudi Arabia (Magin, 1996; Thouless et al., 1997). The Farasan gazelle (Gazella gazella farasani, Thouless and Al-Bassri, 1991) is endemic to the Farasan Archipelago, off-shore the Red Sea coast of Saudi Arabia. The population is still believed to number around 1100 animals (Cunningham and Wronski, 2009). Although, the IUCN Red List classification of vulnerable is based on a population estimated to number <1000 mature individuals (Mallon and Kingswood, 2001), the population of this insular gazelle is characterised by an acute restriction in the number of locations (three islands) and is thus threatened by human nuisance, by stochastic events and devastating disease outbreaks (Flamand et al., 1988; Dunham et al., 2001). During the course of a systematic survey of gastro-intestinal tract parasitic infections in goats and Farasan gazelles and a possible cross-infection between domestic life stock and wild gazelles (Apio and Omer, unpublished data), oocysts of an unknown Eimeria species were detected in the faeces of gazelles.
This parasite is different from that infecting main-land mountain gazelles (Eimeria idmii; Mohammed and Hussein, 1992) and parasite-host specificity seems to be in place between endemic Gazella gazella farasani and the newly described Eimeria species. Data from the literature suggest that the phylogenetic backgrounds, body sizes and morphology and feeding behaviors of animal species serve to promote parasite-host specificity. Differences in these factors among coexisting animal species appear to result in hosts with parasite faunas characteristic of their species. There is laboratory and field evidence that suggests parasites may constitute barriers to sympatry of host species with similar parasite susceptibilities. Taxonomically related host species are susceptible to infection by the same parasite species (Awachie, 1972; Baer, 1961; Blackmore and Owen, 1968; Cameron, 1964; Jenkin, 1963; Segun, 1971; Vinson and Iwantsch, 1980). Evolution of host specificity along host taxonomic lines may result from selection favouring parasites whose antigens more closely resemble those of the host (Dineen, 1963; Jenkin, 1963) parasites capable of coating themselves with host antigen (Smithers et al., 1969); parasites that respond to the host by producing host-like substances (Capron et al., 1968); parasites that depress host immune responsiveness (Cypess et al., 1973; Faubert, 1976) or in other ways control the development of a host defensive response (Vinson and Iwantsch, 1980).
In the present study we show a case of parasite-host specificity by describing a new parasite species which is different from that infecting the taxonomically similar host (Gazella gazella sp.) inhabiting the near Arabian mainland.
MATERIALS AND METHODS
Sampling was carried out during a four-week field study (during October 2009 and February 2010) on Farasan Kebir, the largest island of the Farasan Archipelago harbouring the endemic subspecies of Gazella gazella farasani. A total of 24 fresh faecal samples from gazelles in Farasan Island were collected from the ground into wide-mouth, screw-capped plastic containers immediately after the animal had defecated. A few samples were collected during a systematic search for localised defecation sites in the study area. The numbers of gazelles in the Farasan Island was so small and it was extremely difficult to see a herd of few gazelles and they dispersed singly in wide areas. Samples from such latrines were fresh but could not be assigned to a sex or age class. Faecal samples were preserved in 2.5% (w/v) potassium dichromate (K2Cr2O2) and transferred to the laboratory at King Khalid Wildlife Research Centre (KKWRC). In order to obtain measurements and microphotographs, faecal cultures were examined daily using the floatation method over saturated sodium chloride solution as described by Mohammed and Hussein (1992). Cover slides were searched for unsporulated and sporulated oocysts using a magnification x40 objective lens.
To obtain measurements and microphotographs of sporulated oocysts (sporocysts), three grams of fresh faecal pellets were ground to fine particles using pestle and mortar and mixed with 2.5% (w/v) aqueous solution of potassium dichromate, strained with a fine-mesh wire strainer and suspended in shallow layers of the solution in Petri dishes at room temperature (25±2°C) for sporulation. Measurements of sporulated oocyst were taken using a calibrated ocular micrometer and photomicrographs were made using a Nikon camera (Nikon Company, Japan) attached to a Nikon microscope (Nikon Company, Japan). All measurements were depicted in micrometers (μm, Mean±SD). The description of the new Eimeria sp., was based on measurements of sporulated oocysts, presence or absence of micropyle, micropylar caps, oocyst shape and wall color.
RESULTS AND DISCUSSION
Eleven out of 24 sampled Farasan gazelles (45.8%) were infected with a single species of Eimeria. The number of oocysts per individual sample was only few and it was not easy to quantify the infection using modified McMaster technique unlike coccidian parasites detected in goats. This species of Eimeria was different to those found in domestic goats inhabiting and browsing in the same area as the wild gazelles. The observed Eimeria species is also different from that found in mainland mountain gazelles of Saudi Arabia (Gazella gazella sp.) and from Eimeria species found in other members of the genus Gazell1a (Table 1). Hence, we postulate the detected parasite represents a new species of Eimeria.
Eimeria farasanii n. sp.
Description: Unsporulated oocysts spherical to subspherical, slightly flattened at micropylar end. Oocyst wall 2.9±0.8 (2-4) thick, smooth, double-layered, each of about the same thickness, outer layer bluish green, inner, slightly thicker, layer yellow, with narrow micropyle measuring 3.6±1.1 (3-5) but without micropylar cap. Sporulated oocysts (N = 100) measured, 20.8±1.8x19.5±1.6 (19-25x18-23), length/width ratio 1.07±0.05 (1.04- 1.15). Oocyst residuum and oocyst polar granule are both absent. Each sporulated oocyst with four sporocysts which are elongate, (N = 150) measured 8.2±1.1x3.2±0.7 (7-10x2-5); length/width ratio 2.6±0.5 (2-4). Stieda body present, substieda body absent. Sporocyst residuum present, consists of diffuse, coarse, refractile granules. Sporozoites elongate (N = 50), 6.5±1.2 (6-8), each with a small refractile body at wide end.
Type host: The Farasan gazelle, Gazella gazella farasani (Thouless and Al Bassri, 1991).
Type locality: Miharraq, Farasan Kebir, Farasan Islands, Saudi Arabia.
Prevalence: Found in 11 of 24 Farasan gazelles (45.8%).
Site of infection: Unknown, oocysts recovered from feces. Low level infection was noticed in gazelles and this coccidian parasite was probably of low pathogenicity.
Sporulation: Exogenous, within 7-8 days at 25±2°C in 2.5% K2Cr2O7.
| Table 1: | Morphometric comparison between Eimeria farasanii n. sp. and Eimeria species described from other gazelle species |
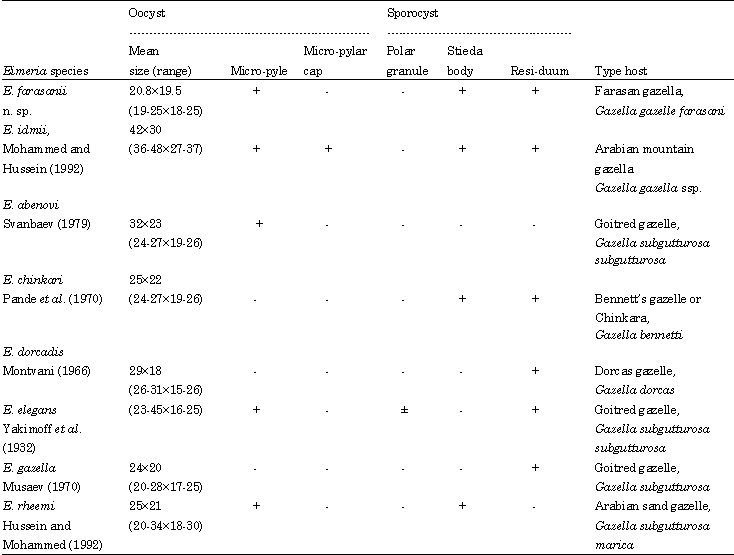 | |
Types: Phototypes and preserved materials in authors’ collection at KKWRC.
Etymology: The specific name is derived from the subspecific name of the type host Farasan Gazelle (Gazella gazella farasani) which inhabits Farasan Islands (Fig. 1, 2).
Levine and Ivens (1986) added another 5 species of Eimeria from other members of the genus Gazella Blainville, 1816. Later, Mohammed and Hussein (1992) and Hussein and Mohammed (1992) added two new eimerain species found in the Arabian sand gazelle and in the Arabian mountain gazelles. Eimeria farasanii n. sp., showed pronounced differences to its closely related relative Eimeria idmii. It has a very small oocyst with no micropylar cap and appears to be the smallest Eimeria species among antelope eimerians (Mohammed and Hussein, 1992). From E. chinkari, E. dorcadis and E. gazella by the presence of the micropyle and from E. abenovi and E. elegans it can be separated by the presence of the sporocyst residuum. The shape of E. farasanii n. sp., as well as the oocyst measurements were closer to E. rheemi reported from the Arabian sand gazelle, however, E. rheemi lacks the sporocyst residuum. Kasim et al. (1991) redesrcibed E. gazella and found out that the oocysts of E. gazella investigated contained numerous small polar granules and a sporocyst residuum. The original description of E. gazella was from the Goitred gazelle (Gazella subgutturosa) and detecting it in Gazella gazella arabica would raise a question of whether E. gazella can infect both Gazella gazella sp. as well as Gazella subgutturosa,Wronski et al. (2010) reviewed the taxonomy of the mountain gazelle in the Arabian Peninsula including gazelles from Farasan Islands.
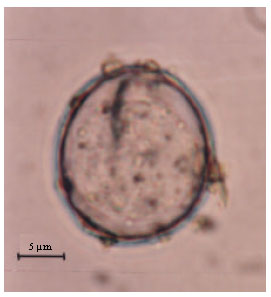 | |
| Fig. 1: | A photomicrograph of unsporulated oocyst of Eimeria farasanii n. sp. from Farasan gazelle (Gazella gazella farasani), scale bar = 5 μm |
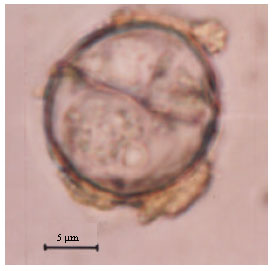 | |
| Fig. 2: | A photomicrograph of sporulated oocyst of Eimeria farasanii n. sp. from Farasan gazelle (Gazella gazella farasani). The mirocpyle with no micropylar cap, sporocyst with steida body and residuum. Scale bar = 5 μm |
Farasan Mountain gazelles are distinguishable from both mainland populations and the type specimen of G. arabica, putatively collected at Farasan in the 19th century (Groves, 1983; Thouless and Al Bassri, 1991). It remains unknown if Kasim et al. (1991) have really examined specimens from Gazella gazella arabica or not.
The fact that Eimeria farasanii n. sp. can be thoroughly separated from all other gazelle and antelope eimerians inhabiting the Arabian Peninsula indicates intra-specific parasite-host specificity.
Parasites depend to a large extent on the evolutionary processes of their hosts, meaning that parasites tend to evolve in parallel with their host species. When two host species have evolved from a common ancestor, the parasites that were originally present in that common ancestor evolve with the two new hosts. So, the modern descendants of the original host ancestor will have parasites descended from those originally present in the ancestral host and the phylogenetic tree of a group of parasites frequently corresponds to the phylogenetic tree of their hosts (Baverstock and Johnson, 1990; Tenter, 1995). Based on phenotypic differences in skull morphometry and fur coloration the subspecies Gazella gazella farasani was described (Thouless and Al-Basri, 1991). Although analysis of mitochondrial DNA indicate that several immigration (or introduction) events form the basis for today’s Farasan gazelle population it was hypothesized that during the last ice age (i.e., app. 16,000 years before today) a land bridge between Farasan Islands and the Arabian Peninsula enabled mainland Mountain gazelles to immigrate to the archipelago (Thouless and Al-Basri, 1991; Bailey et al., 2007).
ACKNOWLEDGMENTS
Our gratitude extends to H.H. Prince Bandar bin Saud bin Mohammed Al Saud, Secretary General, Saudi Wildlife Commission (SWC), for his continued support towards conservation efforts in Saudi Arabia. Our appreciation goes to Thomas M. Butynski (Director KKWRC) for commenting on an earlier draft of this study.
REFERENCES
- Alyousif, M.S., A.A. Kasim and Y.R. Al-Shawa, 1992. Coccidia of the domestic goat (Capra hircus) in Saudi Arabia. Int. J. Parasitol., 22: 807-811.
PubMedDirect Link - Baer, J.G., 1961. Host reactions in young birds to naturally occurring super infections with Porocaecum ensicaudatum. J. Helminthol., 35: 1-4.
CrossRefDirect Link - Baverstock, P.R. and M. Johnson, 1990. Ribosomal RNA nucleotide sequence: A comparison of newer methods used for its determination and its use in phylogenetic analysis. Aust. Syst. Bot., 3: 101-110.
CrossRefDirect Link - Cameron, T.W.M., 1964. Host specificity and the evolution of helminthic parasites. Adv. Parasitol., 2: 1-34.
PubMed - Cypess, R.H., A.S. Lubiniecki, W.M. and A. Hammon, 1973. Immunosuppression and increased susceptibility to Japanese encephalitis virus Trichinella spiralis-infected mice. Proc. Exp. Biol. Med., 143: 473-496.
PubMed - Dineen, J.K., 1963. Antigenic relationship between host and parasite. Nature, 197: 471-472.
CrossRefDirect Link - Dunham, K.M., T.B. Kichenside, N. Lindsay, F.E. Rierkerk and D.T. Williamson, 1993. The reintroduction of mountain gazelle, Gazella gazella, in Saudi Arabia. Int. Zoo Yb., 32: 107-116.
CrossRefDirect Link - Faubert, G.M., 1976. Depression of the plaque-forming cells to sheep red blood cells by the new-born larvae of Trichinella spiralis. Immunology, 30: 485-489.
Direct Link - Groves, C.P., 1983. Notes on the gazelles IV: The Arabian gazelles collected by Hemprich and Ehrenberg. Zeitschrift f�r S�ugetierkunde, 48: 371-381.
Direct Link - Hussein, H.S. and O.B. Mohammed, 1992. Eimeria rheemi sp. n. (Apicomplexa: Eimeriidae) from Saudi Arabian Sand Gazelle, Gazella subgutturosa marica (Artiodactyla: Bovidae) in Saudi Arabia. J. Helminthol. Soc. Washington, 59: 190-194.
Direct Link - Hussein, H.S., A.A. Kasim and Y.R. Al-Shawa, 1987. The prevalence and pathology of Eimeria infections in camels in Saudi Arabia. J. Comp. Pathol., 97: 293-297.
PubMed - Kasim, A.A. Y.R. Al-Shawa, 1985. Prevalence of Eimeria in feces of cattle in Saudi Arabia. Vet. Parasitol., 17: 95-99.
PubMedDirect Link - Kasim, A.A., M.S. Al-Yousif and Y.R. Al-Shawa, 1991. Redescription of Eimeria gazella Musaev, 1970 (Apicomplexa: Eimeriidae) from the Arabian gazelle, Gazella gazella arabica (Artiodactyla: Bovidae) in Saudi Arabia. Parasitologia, 33: 107-109.
PubMed - Kasim, A.A., S.H. Hussein and Y.R. Al-Shawa, 1985. Coccidia in camels (Camelus dromedarius) in Saudi Arabia. J. Protozool., 32: 202-203.
CrossRefPubMedDirect Link - Segun, A.O., 1971. Acephaline gregarines of British earthworms-their possible host specificity. Parasitology, 62: 389-396.
CrossRefDirect Link - Smithers, S.R., R.J. Terry and D.J. Hockley, 1969. Host antigens in shistosomiasis. Proc. R. Soc. Ser. B., 171: 483-494.
Direct Link - Tenter, A.M., 1995. Current research on Sarcocystis species of domestic animals. Int. J. Parasitol., 25: 1311-1330.
CrossRef - Thouless, C.R. and K. Al Bassri, 1991. Taxonomic status of the Farasan island gazelle. J. Zool. Lond., 223: 151-159.
CrossRefDirect Link - Toulah, F.H., 2007. Prevalence and comparative morphological studies of four Eimeria sp. of sheep in Jeddah area, Saudi Arabia. J. Biol. Sci., 7: 413-416.
Direct Link - Vinson, S.B. and G.F. Iwantsch, 1980. Host suitability for insect parasitoids. Annu. Rev. Entomol., 25: 397-419.
CrossRef - Wronski, T., T.J. Wacher, R.L. Hammond, B. Winney and K. Hundertmark et al., 2010. Two reciprocally monophyletic mtDNA lineages elucidate the taxonomic status of Mountain gazelles (Gazella gazella). Syst. Biodiver., 8: 1-10.
CrossRefDirect Link - Yakimoff, W.L., W.F. Gousseff and E.F. Rastegaieff, 1932. Die coocidiose der wilden kleinen Wiederkauer. Parasitol. Res., 5: 85-93.
CrossRefDirect Link - Mohammed, O.B. and H.S. Hussein, 1992. Idmii eimeria n. sp. (Apicomplexa: Eimeriidae) from the arabian mountain gazelle, Gazella gazella in Saudi Arabia. J. Helminth. Soc. Washington, 59: 120-124.
Direct Link