
B.M. Md-Zain
School of Environmental and Natural Resource Sciences, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
C.E. Ch`ng
School of Environmental and Natural Resource Sciences, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
International Journal of Zoological Research
Year: 2011 | Volume: 7 | Issue: 1 | Page No.: 59-67







ABSTRACT
This study describes the activity budget of a group of Cantor’ dusky leaf monkeys (Trachypithecus obscurus) in Penang Botanical Garden, Malaysia. The main objective is to investigate the daily activity budget of T. obscurus in the field. Scanning sampling was employed as the observation method from June 2006 to December 2006. Behavioral observation of 18 individuals were conducted for 8 hours per day, from 0700 h to 1630 h. On average, T. obscurus spent most of their time on feeding (40.02%), followed by the resting (23.96%), moving (22.51%), autogrooming (6.52%), vocalization (3.89%), allogrooming (1.33%), allomothering (0.97%) and the least in playing (0.80%). Chi-square test showed that there is a significant difference among every behavior category of Cantor’ dusky leaf monkeys. The habitat and canopy used of Cantor’ dusky leaf monkeys in Penang Botanical Garden were also described.
PDF Abstract XML References Citation
Received: April 20, 2010;
Accepted: June 15, 2010;
Published: August 21, 2010
How to cite this article
B.M. Md-Zain and C.E. Ch`ng, 2011. The Activity Patterns of a Group of Cantor’ Dusky Leaf Monkeys (Trachypithecus
obscurus halonifer). International Journal of Zoological Research, 7: 59-67.
DOI: 10.3923/ijzr.2011.59.67
URL: https://scialert.net/abstract/?doi=ijzr.2011.59.67
DOI: 10.3923/ijzr.2011.59.67
URL: https://scialert.net/abstract/?doi=ijzr.2011.59.67
INTRODUCTION
Trachypithecus obscurus is a member of subfamily Colobinae (Groves, 2001, 2005) which constitutes of eleven subspecies (Brandon-Jones et al., 2004): T. o. obscurus, T. o. corax, T. o. flavicauda, T. o. halonifer, T. o. carbo, T. o. styx, T. o. phayrei, T. o. shanicus, T. o. smithi, T. o. seimundi and T. o. sanctorum. Geographical distribution of T. obscurus ranges from Peninsular Malaysia to India (Brandon-Jones et al., 2004). T. obscurus is commonly called as dusky leaf monkey or spectacled leaf monkey due to its dietary habit and white mascara patch over the eye respectively. In Malaysia, it is locally called as cengkong, lotong mata puteh or lotong bercelak regarding to its unique physical characters (Marsh and Wilson, 1981). T. o. obscurus (Reid’s dusky leaf monkey) is the major subspecies that exists in Peninsular Malaysia, covering from the north part of Malaysia up until Thailand (Brandon-Jones et al., 2004). T. o. styx (Perhentian dusky leaf monkey) occurs in Perhentian Island and probably the adjacent coast (Brandon-Jones et al., 2004). T. o. halonifer (Cantor’s dusky leaf monkey) is another subspecies that can be traced in Dayang Bunting Island, Langkawi Island and Penang Island (Brandon-Jones et al., 2004).
Previous primate research in Malaysia mostly involved molecular aspects for both human primate (Shahrom et al., 2005; Lim et al., 2010) and non-human primates (Jalil et al., 2008; Md-Zain et al., 2008a; 2010). In addition, molecular systematic studies on Malaysian primates were also focused at the genus Trachypithecus of both T. obscurus and T. cristatus (Ernie et al., 2005; Md-Zain et al., 2005; 2008a). Other molecular studies from neighbouring country (Thailand) also incorporated samples of T. obscurus (Chaveerach et al., 2007; Tanee et al., 2009). Recently, many primatologists have concentrated on primate ecology and behavior both in the field (Ampeng and Md-Zain, 2007) and captivity (Arnold and Barton, 2001; Md-Zain et al., 2008b) with little emphasis put on dusky leaf monkeys. The field observation of dusky leaf monkey is relatively scarce due to their shyness, less predictable habits, greater stealth and tendency to disperse (Chivers, 1980). In this study, the field observation of Cantor’s dusky leaf monkeys was carried out quantitatively to identify their daily activity patterns. It is hoped that this study can add more information as the other previous behavioral studies had done especially in understanding the behavior of the Malaysian primates (Alonso-Spilsbury et al., 2006; Siddique, 2008; Zainudin et al., 2009; Zakaria et al., 2009).
MATERIALS AND METHODS
This research was conducted in 2006 at Penang Botanical Garden, Malaysia (Fig. 1). The study site is a historical public botanical garden established in 1884 by Charles Curtis, who enthused to bear the seeds of exotic plants from distant lands. Penang Botanical Garden occuppied an area of 30 hectares is inhabiting by fine collection of exotic flora and fauna, especially the long-tailed macaques (Macaca fascicularis) and Cantor’s dusky leaf langurs. The habituated long-tailed macaque of the garden has earned it another name as the Monkey Garden.
Dusky leaf monkeys are found to exist and forage at seven different sites where five of the sites are located within the garden while another two are at the adjacent areas (Fig. 1). The study group of dusky leaf monkeys at Penang Botanical Garden is composed by 18 individuals that comprises of an adult male, five adult females, 11 sub-adult females and one infant (Table 1).
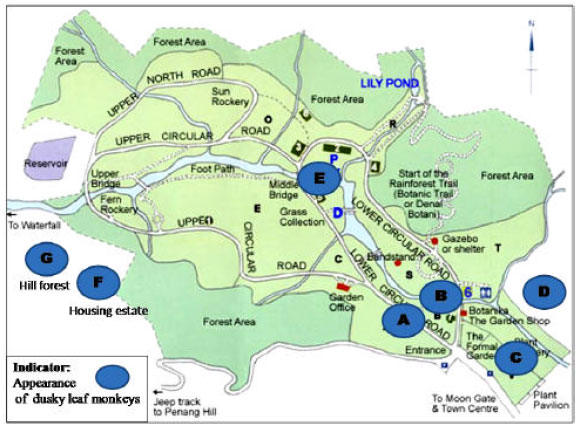 | |
| Fig. 1: | Map of Penang Botanical Garden indicating the appearance sites of dusky leaf monkeys |
| Table 1: | Categories and description of study subjects |
 | |
This subject group is considered as habituated in the term of not running away or accustomed to the presence of the observer (Curtin, 1980).
Preliminary observation was conducted in order to be familiar with subjects and to determine types of activity patterns of the subjects (Altmann, 1974; Martin and Bateson, 1993). Preliminary observation was conducted for four days from June 16-21 2006. Scanning sampling was employed in the intensive observation for 18 days from June 2006 to December 2006. The sample session was scheduled from 0700 to 1630 h, 8 h per day excluding an hour and 30 min break. Every sampling subsession was 15 min with 10 min confined to scan sampling while another 5 min was used to describe and make brief conclusion of each subsession. Chi-square test was applied to analyze the conducted behavior data set. This nonparametric test is suitable to analysis the significance of activity budget which does not follow the normal distribution (Martin and Bateson, 1993).
RESULTS AND DISCUSSION
Profiles on activity pattern: Cantor’s Dusky leaf monkey is known as a social primate. They had been conducting their daily activity within a social group. Nevertheless, they seldom get involve in social interaction with other members during observation, where they foraged and fed individually within the group. The greatest proportion of activity budget of Cantors dusky leaf monkey was devoted to feeding (40.02%), followed by resting (23.96%) and moving (22.51%). The indirect social behaviors, autogrooming (6.52%) and vocalization (3.89%) contributed to a relatively large portion of time compared to the other social behaviors, allogrooming (1.33%), allomothering (0.97%) and playing (0.80%) (Fig. 2). Every daily activity of the dusky leaf monkey has significant different (χ2 = 39380.37; df = 7, p = 0.05) meaning that there were differences occurred during the daily activity (Table 2). The same behavior patterns shown by Geoffroy’s marmoset indicated that more time in feeding and foraging than to social behavior (Passamani, 1998). Present results however contradict with these previous studies which found out that many colobines spend most of their time in resting rather than feeding: T. leucocephalus (Li and Rogers, 2004), T. francoisi (Zhou et al., 2007), Colobus guereza (Fashing, 2001) and C. vellerosus (Teichroeb et al., 2003). However, our results are in line (Zhou et al., 2007) as they found out that François’ langurs spent little time on social activities (grooming and playing). Leaf monkeys will adjust their activity budgets in response to the change of ecological conditions particularly when food resources are scarce (Zhou et al., 2007). Thus, in this study, we found that Cantor’s dusky leaf monkeys increased their feeding time may due to maintain a certain daily energy intake.
The daily activity pattern of this study has been recalculated as an average daily percentage allocated to each activity. As the observation was conducted during two different periods (June 2006 and December 2006), the daily behavior allocation was presumed to be affected by some ecological settings or life span. Feeding behavior was the major behavior of the daily activity budget, by occupying more than 30% of the daily observation time.
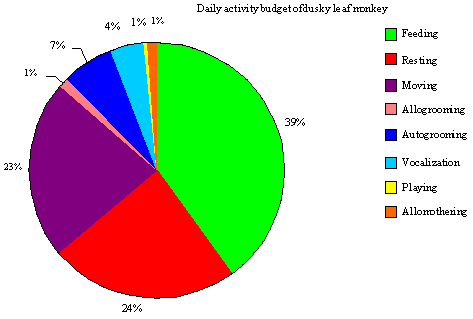 | |
| Fig. 2: | Percentage of daily activity budget of dusky leaf monkeys |
| Table 2: | Percentage and Chi-square value of dusky leaf monkey’s activity budget at penang botanical garden |
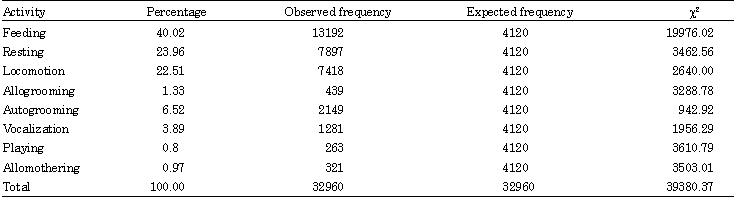 | |
| χ2: 39380.37; p<0.05; degrees of freedom = 7 | |
As no nutrition test has been conducted on the food items consumed by the langurs, there is no significant evidence showing that either the highest feeding activity was contributed by the poor nutrient (Li and Rogers, 2004) or the abundance of the food (Passamani, 1998). The feeding allocation was relatively higher from 12 to 16 December as young shoot of rain tree (food source) was sprouting within that period. The feeding time of Rhinopithecus bieti has changed parallel with the seasonality (Ding and Zhao, 2004). Yet, there was no obvious seasonality change at Penang Botanical Garden. Dusky leaf monkeys were seen to consume food more on leaves as many Asian colobines (Dela, 2007; Solanki et al., 2008; Zhou et al., 2009).
The abundant and spatial patterning of food resources affect the ranging and activity budgets of Old World Monkeys (Kinnaird and O’Brien, 2000; Fashing et al., 2007). In this study, the percentage of resting behavior was recorded to be lower when the other motion-based behavior increased. The most obvious negative correlation was between feeding and resting behavior (Kurup and Kumar, 1993) and between playing and resting (Li and Rogers, 2004). However, it has still employed the second largest portion of the activity budget because food resources of dusky leaf monkeys in Penang Botanical Garden are abundance. Results showed that frequency of movement was distributed evenly during the whole observation period with 20% per day. In daily percentage allocation, movement has no correlation with feeding behavior. It could be explained that the langurs’ movements were not restricted in foraging or resource directed but has been contributed by other non-foraged movements, such as traveling (Kramer, 2001). Fashing et al. (2007) found out that C. angolensis spent markedly less time resting, spent more time feeding and moving and travelled much longer distances that is due to substantial energetic costs to living in large groups.
In this study, Cantor’s dusky leaf monkeys allocated less time on social activities (grooming, playing and allomothering). Allogrooming indicated the negative correlation with the feeding behavior from 12 to 16 December 2006. The langurs have reduced their social interaction when they were active in feeding. Autogrooming and vocalization have shown no trend in daily activity allocation. Autogrooming is presumed to be a non-functional behavior in this study as it did not show hygiene (Hutchins and Barash, 1976) or mutualistic social interaction (Goosen, 1974) during the observation. Somehow, it was recorded to be in higher frequency during rainy day. Vocalization has a relatively higher frequency during playing session. Playing behavior was also distributed evenly with an extremely low percentage. However, it has recorded relatively larger fraction (>2%) for 2 days of observation. Apparently, allomothering was affected by the age and development of the infant. Its percentage only increased during observation in December 2006, when the infant has grown up and is capable to wander around with its mother. It was a contrast from the previous study which indicated that the allomothering will decrease or cease with the growth of the infant (Eimerl and DeVore, 1980). Overall, every behavior category showed its unique daily trend during the whole observation. The fluctuation of the dusky leaf monkey daily behavior was not obvious.
Habitat use: The habitat used was evaluated by the amount of times visited by the Cantor’s dusky leaf monkeys. The same method was used by Kaplin (2001) to determine the habitat use of guenons. Habitat quality may be a factor causing different activity budget between populations of same species (Li and Rogers, 2004). Behavior could be affected by the different habitat use, such as dispersal of resources and habitat-related adaptations (Singh and Vinathe, 1990). A more widely ranging or diverse pattern would allow the group to monitor the environment for resource availability (Garber, 1993).
Site A is the entrance of the Penang Botanical Garden which planted with a rain tree (Fig. 1). Throughout the observation, site A was visited most frequently by langurs (Table 3). The prominent big tree is the favorite food source of dusky leaf monkeys. This situation relates to previous finding in Blue monkey group that was recorded to visit habitats with abundant favourite food (Kaplin, 2001).
| Table 3: | Frequency of appearance of dusky leaf monkey at different sites within penang botanical garden |
 | |
Their ranging behavior and distance are the response to the availability of specific resources and dietary diversity (Kaplin, 2001). Young shoot or leaves with high nutrient have attracted langurs to visit early in the morning (Ding and Zhao, 2004). Cercopithecus aethiops were found to practice optimal foraging theory by spending longer duration of feeding bouts in higher-quality patches (Whitten, 1998). However, Cantor’s dusky leaf monkeys did not totally practiced the optimal foraging theory because their foraging area has occupied a large patch of garden. They need to employ more energy to forage in such a large area. This is true as the temporal and spatial distribution of food resources may be the most important determinant of leaf monkey ranging behaviour (Tan et al., 2007).
Cantor’s dusky leaf monkeys seem to exhibit the same ranging habit. Site B and F are occupied with medium-sized trees with adjacent buildings. A souvenir shop and rural housing area are located at the central of site B and F, respectively. However, langur only visited once at the housing area during the observation (Table 3). Overall, the langur group did not utilize many sites in Penang Botanical Garden evenly, but had been visiting certain site regularly with certain essential factor. Presbytis potenziani has occasionally utilized the gardens and secondary forest besides the predominant primary forest. It is a common trend in Mentawai primates that exists among the forest communities, human settlements and commercial logging areas (Fuentes, 1996).
Canopy use: Southeast Asian Presbytis and Trachypithecus tend to use the middle and upper strata of the primary or tall forests (with canopy > 20 m) (Curtin, 1980; Bennett and Davies, 1994). The same observation was found at Penang Botanical Garden while most of the activities of dusky leaf monkeys were concentrated at higher strata of canopy (site A, C, E and G). These sites are filled with the highest layer of canopy by tall and broad canopy. The feeding pattern is an indicator of the abundance and the distribution of the food (Passamani, 1998). Indeed, most of the langur’s food sources comes from this layer, such as rain tree (site A), nutmeg (site C), bamboo (site E) and hill forest (site G). Normally, dusky leaf monkeys were fed on the tree-crowns of tall trees, such as figs which have many seeds especially during the fruit scarcity period (Stevenson, 2004).
Cantor’s dusky leaf monkeys were observed to utilize every layers of canopy in range as long as there are supporting substrates ready for them. This finding supports previous finding by Ampeng (2006) on Presbytis melalophos chrysomelas which utilized every part of canopy layer in Samunsam Wildlife Sanctuary, Sarawak. The incomplete habituation of P. potenziani has restricted them from using the primary supports at <15-10 m at dipterocarps forest (Fuentes, 1996). Apparently, Cantor’s dusky leaf monkeys group at Penang Botanical Garden has no limitation in utilizing any layer of canopy. They even found out to the range at the housing area (site F) and cross the electricity cable to site G. From this point, the subjects are assumed to be habituated to the activity’s sites.
ACKNOWLEDGMENTS
We are deeply indebted to several institutions that provided necessary facilities and assistance including Universiti Kebangsaan Malaysia, Department of Wildlife and National Parks and Penang Botanical Garden management. We wish to thank Farhana Shukor and anonymous reviewers for their comments on the manuscript. This research was made possible under grants IRPA 0802020019 EA301, ST-019-2002 and UKM-KRIB-16/2008.
REFERENCES
- Alonso-Spilsbury, M., I. Escobar-Ibarra, L. Mayagoitia, R. Ramirez-Necoechea and D.Mota-Rojas, 2006. Social and sexual behavior of two newly formed pairs of mexican gray wolf in captivity. Int. J. Zool. Res., 2: 334-343.
CrossRefDirect Link - Altmann, J., 1974. Observational study of behavior: Sampling methods. Behaviour, 49: 227-267.
CrossRefDirect Link - Arnold, K. and R.A. Barton, 2001. Postconflict behavior of spectacled leaf monkeys Int. J. Primatol., 22: 243-266.
CrossRef - Brandon-Jones, D., A.A. Eudey, T. Geissmann, C.P. Groves and D.J. Melnick et al., 2004. Asian primate classification. Int. J. Primatol., 25: 97-164.
Direct Link - Chaveerach, A., T. Tanee, N. Sattayasai, A. Tanomtong, S.A. Suarez and S. Nuchadomrong, 2007. Genetic relationships of langur species using AFLP markers. Pak. J. Biol. Sci., 10: 1445-1451.
CrossRefPubMedDirect Link - Dela, J.D.S., 2007. Seasonal food use strategies of Semnopithecus vetulus nestor, at Panadura and Piliyandala, Sri Lanka. Int. J. Primatol., 28: 607-626.
CrossRef - Ding, W. and Q.K. Zhao, 2004. Rhinopithecus bieti at Tacheng, Yunnan: Diet and daytime activities. Int. J. Primatol., 25: 583-598.
CrossRef - Fashing, P.J., 2001. Activity and ranging patterns of guerezas in the Kakamega forest: Intergroup variation and implications for intragroup feeding competition. Int. J. Primatol., 22: 549-577.
CrossRef - Fashing, P.J., E.S. Dierenfeld and C.B. Mowry, 2007. Influence of plant and soil chemistry on food selection, ranging patterns and biomass of Colobus guereza in Kakamega forest, Kenya. Int. J. Primatol., 28: 673-703.
CrossRef - Fuentes, A., 1996. Feeding and ranging in the Mentawai Island Langur (Presbytis potenziani). Int. J. Primatol., 17: 525-548.
CrossRef - Garber, P.A., 1993. Seasonal patterns of diet and ranging in two species of tamarin monkeys: Stability versus variability. Int. J. Primatol., 14: 145-166.
CrossRef - Groves, C., 2001. Primate Taxonomy. Smithsonian Institution Press, Washington, ISBN-10: 156098872X.
Direct Link - Hutchins, M. and D.P. Barash, 1976. Grooming in primates: Implications for its utilitarian function. Primates, 17: 145-150.
CrossRef - Jalil, M.F., J. Cable, J. Sinyor, I. Lackman-Ancrenaz, M. Ancrenaz, M.W. Bruford and B. Goossens, 2008. Riverine effects on mitochondrial structure of Bornean orang-utans (Pongo pygmaeus) at two spatial scales. Mol. Ecol., 17: 2898-2909.
CrossRef - Kaplin, B.A., 2001. Ranging behavior of two species of Guenons (Cercopithecus lhoesti and C. mitis doggetti) in the nyungwe forest reserve, Rwanda. Int. J. Primatol., 22: 521-548.
CrossRef - Kinnaird, M.F. and T.G. O'Brien, 2000. Comparative Movement Patterns of Two Semiterrestrial Cercopithecine Primates: The Tana River Crested Mangabey and the Sulawesi Crested Black Macaque. In: On the Move: How and Why Animals Travel in Groups, Boinski, S. and P.A. Garber (Eds.). University of Chicago Press, Chicago, pp: 327-350.
- Kurup, G.U. and A. Kumar, 1993. Time budget and activity patterns of the Lion-Tailed Macaque (Macaca silenus). Int. J. Primatol., 14: 27-39.
CrossRef - Li, Z. and E. Rogers, 2004. Habitat quality and activity budgets of white-headed langurs in fusui, China. Int. J. Primatol., 25: 41-54.
CrossRef - Lim, L.S., K.C. Ang, M.C. Mahani, A.W. Shahrom and B.M. Md-Zain, 2010. Mitochondrial DNA polymorphism and phylogenetic relationships of proto malays in peninsular Malaysia. J. Biol. Sci., 10: 71-83.
CrossRefDirect Link - Md-Zain, B.M., J.C. Morales, M.N. Hassan, A. Jasmi, M. Lakim, J. Supriatna and D.J. Melnick, 2008. Is Presbytis a distinct monophyletic genus: Inferences from mitochondrial DNA sequences. Asian Primates J., 1: 26-36.
Direct Link - Md-Zain, B.M., M.Y. Yen and I.A. Ghani, 2008. Daily activity budgets and enrichment activity effect on Chimpanzees (Pan troglodytes) in captivity. Sains Malaysiana, 37: 15-19.
Direct Link - Md-Zain, B.M., S.J. Lee, M. Lakim, A. Ampeng and M.C. Mahani, 2010. Phylogenetic position of Tarsius bancanus based on partial cytochrome b DNA sequences. J. Biol. Sci., 10: 348-354.
CrossRefDirect Link - Passamani, M., 1998. Activity budget of geoffroy's marmoset (Callithrix geoffroyi) in an Atlantic forest in Southeastern Brazil. Am. J. Primatol., 46: 333-340.
Direct Link - Siddique, Y.S., 2008. Breeding behavior of Copsychus saularis in Indian-sub-continent: A personal experience. Int. J. Zool. Res., 4: 135-137.
CrossRefDirect Link - Singh, M. and S. Vinathe, 1990. Inter-population differences in the time budgets of Bonnet Monkeys (Macaca radiata). Primates, 31: 589-596.
CrossRefDirect Link - Solanki, G.S., A. Kumar and B.K. Sharma, 2008. Feeding ecology of Trachypithecus pileatus in India. Int. J. Primatol., 29: 173-182.
CrossRefDirect Link - Stevenson, P.R., 2004. Fruit choice by woolly monkeys in tinigua national park, Colombia. Int. J. Primatol., 25: 367-381.
CrossRef - Tan, C.L., S. Guo and B. Li, 2007. Population structure and ranging patterns of Rhinopithecus roxellana in zhouzhi national nature reserve, Shaanxi, China. Int. J. Primatol., 28: 577-591.
CrossRef - Teichroeb, J.A., T.L. Saj, J.D. Paterson and P. Sicotte, 2003. Effect of group size on activity budgets of Colobus vellerosus in Ghana. Int. J. Primatol., 24: 743-758.
CrossRef - Whitten, P.L., 1998. Effects of patch quality and feeding subgroup size on feeding success in vervet monkeys (Cercopithecus aethiops). Behaviour, 105: 35-52.
Direct Link - Zakaria, M., M.N. Rajpar and A.S. Sajap, 2009. Species diversity and feeding guilds of birds in paya indah wetland reserve, Peninsular Malaysia. Int. J. Zool. Res., 5: 86-100.
CrossRefDirect Link - Zhou, Q., C. Huang, Y. Li and X. Cai, 2007. Ranging behavior of the francois langur (Trachypithecus francoisi) in the Fusui nature reserve, China. Primates, 48: 320-323.
CrossRef - Zhou, Q., Z. Huang, X. Wei, F. Wei and C. Huang, 2009. Factors influencing interannual and intersite variability in the diet of Trachypithecus francoisi. Int. J. Primatol., 30: 583-599.
CrossRef - Tanee, T., A. Chaveerach, R. Sudmoon, A. Tanomtong and P. Mokkamul, 2009. Genetic relationships of cercopithecidae in thailand as inferred from rDNA ITS regions. J. Biol. Sci., 9: 51-56.
CrossRefDirect Link