
S. Sujatha
St. Xaviers College (Autonomous), CPRC, Palayamkottai, Tirunelveli, India
S. Jeyaparvathy
St. Xaviers College (Autonomous), CPRC, Palayamkottai, Tirunelveli, India
M. Jasmine Priya
St. Xaviers College (Autonomous), CPRC, Palayamkottai, Tirunelveli, India
International Journal of Zoological Research
Year: 2011 | Volume: 7 | Issue: 4 | Page No.: 310-319







ABSTRACT
The objective of this study was to determine the bacterial pathogen and gene content of the Etroplus suratensis (pathogenic shrimp). PCR-based subtractive hybridization method detected with the help of Escherichia coli on shrimp liver and granulose cells for efficiently detecting the DNAs and applies in to liver pathogen. Seventeen DNAs specific to a mono-key colonizing strain (J140) were obtained by subtractive hybridization against an unrelated strain whose genome has been fully sequenced (ABO18799). Among the seventeen different clones 14 were unique and other three numbers were each represented at twice. Nine of the clones DNAs were found by sequencing to be absent or very divergent from those in the same direction on disappeared blank region of this liver and granulosa cells. Nine others had no database match with proteins of assigned function. PCR tests of 13 unrelated E.coli strains by using primer specific for 12 subtracted clones and complementary southern hybridizations indicated that these DNAs are highly polymorphic in the E. coli population with each strain yielding a different pattern of gene-specific PCR amplification. When described about this Polymorphic immune response gene, it helps to identify the previously unknown virulent genes in the pathogens and it provides a new insight of microbial genetic diversity and evolution.
PDF Abstract XML References Citation
Received: February 07, 2011;
Accepted: May 25, 2011;
Published: September 26, 2011
How to cite this article
S. Sujatha, S. Jeyaparvathy and M. Jasmine Priya, 2011. Biotechnological Based Subtractive Hybridization and Variations of Gene Content in an Organ of Pathogenic Shrimp. International Journal of Zoological Research, 7: 310-319.
DOI: 10.3923/ijzr.2011.310.319
URL: https://scialert.net/abstract/?doi=ijzr.2011.310.319
DOI: 10.3923/ijzr.2011.310.319
URL: https://scialert.net/abstract/?doi=ijzr.2011.310.319
INTRODUCTION
Fishes are the diverse group of animals, highly specialized for their aquatic existence and comprising almost half the number of the vertebrate species in existence today (Hedge et al., 2000). Fishes were in an intimate contact with their environment which can contain very high concentrations of bacteria and viruses (Shike et al., 2002). Very recently, Rane and Barve (2011) described standardization and optimization of gene content and molecular genetic analysis of D-loop region in an animal natural fiber. Genes that are present in certain isolates of a given bacterial species and absent are substantially difficult in others can be of great interest biologically. Some are strain specific traits such as immune responsive drug resistance (Bayne et al., 2001). Later, Pan et al. (2005) studied the immune-relevant factors responsible for the virus resistance in the WSSV-resistant shrimp, a suppression subtractive hybridization method was employed to identify differentially expressed genes and their expression profile in the hepatopancreas of the virus-resistant penaeid shrimp. Thirty-five genes were identified from more than 400 clones, of which eight are found for the first time in penaeid shrimp.
Many of the genes or DNA segments specific to individual strains were found by the special phenotypes they confer (Waldor and Mekalamos, 1996; Gorisman and Ochman, 1996). It was also reported that the detailed physical mapping 10 or comparisons of sequence data from different isolates of taxes (Himmelreich et al., 1997; Bobe and Goetz, 2000a, b; Kanamori, 2000). During the subtractive hybridization 13 allows strain specific DNAs to be selected directly and is attractive because it eliminates the need to score any particular phenotype or to do extensive mapping or sequencing at the outset. However, the subtractive methods developed to date have been unwieldy technically and tend to be quite narrowly selective (Douglas et al., 2003; Thangaraj et al., 2011). Very recently, James et al. (2010) published a subset of strain specific DNAs are generally obtained and DNA segments with potentially interesting mixes of pathogenic strain-specific and common sequences are generally excluded (Tinsley and Nassif, 1996).
The vertebrate immune system is composed of two types of immunity, innate and adaptive. This kind of immunity emerged early in vertebrate evolution at some time during the division of the jawless lamprey and the cartilaginous fishes (Fujiki et al., 2000). An adaptive immune response is called for when the innate host defenses are defeated or by passed and when the elimination of a new infection is unsuccessful (Bayne et al., 2001). There are many commonalities in the immune response of different vertebrate classes. In Teleost fish, however some unique features, such as temperature dependent acquired immunity and some limitations on antibody diversity (Kaattari and Piganelli, 1996). Hence, the innate immune functions may play a more important role in these animals than in the more derived homeothermic invertebrates such as fish and amphibians.
The present study is the first approach to the study of immune response of bacterial infection in a teleost fish of Etroplus. This has been employed Suppression Subtractive Hybridization (SSH) a new method based upon the Polymerase Chain Reaction (PCR) and requires far smaller amount of starting materials than traditional methods. There are number of techniques available to isolate regulated genes (Van Hal et al., 2000; Aziz et al., 2010). The choice of technique was somewhat dependent on the desired result and how much RNA can initially be obtained for subtraction. However, the more recent approaches such as SSH and DD-PCR are clearly more efficient and sensitive than the original techniques involving cDNA subtraction techniques. The objective of this study was to determine the bacterial pathogen and its gene content of the Etroplus suratensis (pathogenic shrimp).
MATERIALS AND METHODS
This experiment was carried out during the year of March 2006 to February 2007. The experimental Shrimp juveniles weighing approximately 5 g were collected from the backwaters of Tamilnadu and Kerala. The tank was filled with water and maintained with adequate aeration. The fishes were fed with pelted feed. The water in the holding tank was exchanged daily and the water quality parameters were monitored weekly.
Collection of bacterial pathogens: Fish bacterial pathogens used for the experiments were obtained from the Microbiology Laboratory. Isolates such as Vibrio esturiances and Aeromonas hydrophila characterized earlier were used. In order to prove the pathogenecity profile of bacterial isolates and verify the Koch’s postulates, experimental challenge studies were conducted using apparently healthy fishes.
General methods: The Esherichia coli and H. pylori strains used in this study are listed in Table 1. STD methods (Tiffoche et al., 1993; Gerwick et al., 2002) were used for the growth and DNA extraction from E. coli.
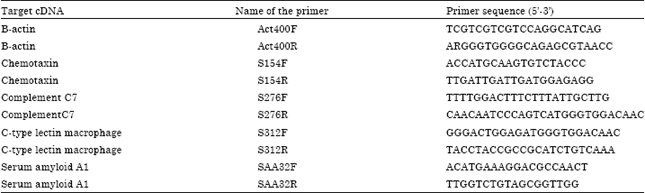
Oligonucleotides: E. coli strains-specific oligonucleotide primers used for PCR listed in Table 2. The following gel purified oligonucleotides were used for subtractive hybridizations. Adaptor-1 5’GATTATGCACTGTGTCTACCCGAGCTCCGCCCGCGGGCCCGACCA-3’ and 3’CCGGGCAGGA-5’; Adaptor2. 5’-GATTATTGCTGAGTGAGTGATATCCCGTCGCACCAGGCCCGGCTCCA-3’ and 3’-CGGCCGAGGT-5’; p1: 5’- GATTATGCTGAGTATGATCCCG-3’ (This P1 PCR primer matches the long strands of adaptors 1 and 2 at their 5’ ends.) NPI 5’- AGCTCGCGGCGGGCCCGTCCA-3’-3’; NP2 5’-GGCACCACGCCGGCGTGCA-3’. (These NP1 and NP2 nested PCR primer match the internal portions of the long strand of adaptors one and two, respectively).
Overview of subtractive hybridizations: This method involved the digestion of DNAs from the strain of interest and a reference (tester and driver, respectively) with restriction endonucleases such as Rsa1 to generate DNA fragment populations with median sizes of about 0.5 kb. Two different PCR adapters that can join only to 5’ ends of target DNAs because of their own 5’ ends lack of phosphate groups were ligated to different aliquots of tester DNA. These ligated DNAs were denatured then mixed with an excess of driver DNA (that has no adapters) and allowed to anneal. The two DNA pools were mixed together also add with additional denatured driver DNA for further bind tester sequences that are also present in the driver genome.
| Table 1: | E. coli strains and used in these studies |
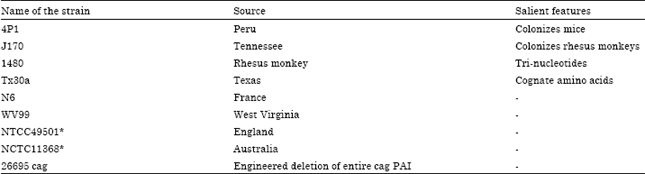 | |
| Each strain, except Tx30a WV99, 26695 cag contains the cag PAI in strain Hp1 contains an internal deletion (D. Kersulyte and D.EB, unpublished work). *NTCC, National Collection of Type Cultures | |
| Table 2: | Name of the experimentally designed primer and primer sequence |
 | |
Remaining complementary single strands of tester DNA are allowed to anneal and the adaptor sequences are copied on to their 3’ ends. Then PCR was used to obtain the exponential amplification of tester DNAs with the same adaptor at each end is suppressed because self-annealing of inverted repeat adaptors (Fig. 1) which inhibits the binding of PCR primers. Tester DNAs with an adaptor at only one end undergo linear but not exponential, amplification. This method offers several substantial advantages over earlier bacterial genome subtractive methods (i) less DNA was needed; (ii) multiple rounds of hybridization and physical removal of tester-driver DNA complexes as in ref.13 and are not needed. Earlier, Tinsley and Nassif (1996) stated that there was no need for complicated adaptor removal and re-addition with additional rounds of PCR amplification by difference analysis method.
Driver and tester DNA preparations: Two micrograms of tester and DNAs were each digested to completion with 20 units of AluI (made up where) for 16 h in 200 μL reaction volumes, extracted with phenol and precipitated with ethanol and resuspended in 10 mM Tris HCl, pH 7.5 at a final concentration of 200 ng μL-1. Two aliquots of tester DNA (120 ng each) were ligated separately to 2 μL of the two adapters each in a total volume of 10 μL of (2 μmM final concentration) at 16°C at overnight, using 1 μL of T4 DNA ligase in the buffer supplied by manufacturer. After ligation, 1 μL of 0.2 M EDTA was added and the samples were heated at 70°C for 5 min to inactive the ligase and then stored at -20°C.
Subtractive hybridization: Three micro liters of driver DNA (600 ng) was added to 1 μL (12 ng) of each of the adaptor-ligated tester DNAs (50:1 ratio). One microliter of 5 x hybridization buffer (2.5 M NaCl/250 mM Hepes, pH 8.3/1 mM EDTA) was added to each tube the solutions were overlaid with mineral oil and the DNAs were denatured (1.5 min, 90°C) and allowed to anneal for 1.5 h at 60°C. After this first hybridization, the two samples (first with adaptor 1, the second with adaptor 2) were combined, 300 ng more of heat-denatured driver DNA was added in 3 μL of 1 x hybridization buffer and the sample was allowed to hybridized for an additional 14 h at 65°C (without intermediate denaturation). This final 13 μL reaction was diluted 200 μL with dilution buffer (50 mM NaCl/5 mM EDTA), heated at 65°C for 10 min and stored at -20°C until use in PCR.
Two sequential PCRs were carried out. The first PCR contained 1 μL of genomic DNA prepared as described, 2 μL of PCR primer P1 (10 μM) and 47 μL of a PCR master mix prepared using the advantage cDNA PCR core kit (total volume 50 μL). This first PCR was incubated at 72°C, 2 min and then subjected in to 25 cycles of 95°C, 30 sec; 72°C for 1.5 min. The amplified products were then diluted 20 fold in 10 mM Tris HCl pH 7.5 and 1 μL of each diluted samples was used in the second PCR with 2 μL of PCR primers with NPI and NP2 in 10 μM each and advantage cDNA PCR core kit (total volume, 50 μL) for 10 cycles of 90°C, 30 sec with 68°C, 30 sec and 72°C 1.5 min.
Construction of SSH libraries: Total tissue extracted RNA was used to synthesized cDNA for SSH according to the manufacturer instructions. Briefly cDNA has been synthesized from challenged fish (tester-cDNA) and unchallenged fish (driver-cDNA) then RNA treated with an oligo (dT) containing primer. Tester and driver-cDNA were separately digested with Real shorter blunt ended molecules. Two ester populations were ligated to two different adapters. The tester cDNA was hybridized with an excess of driver-cDNA to remove common cDNA transcripts, generating the (Single Subtractive Hybridization) SSH+library. The tester cDNA were hybridized with water instead of driver cDNA to provide unsubstracted cDNA it act as a control.
Preparation of driver cDNA probes: DIG labeled probes were made by PCR labeling according to the manufacturer’s instructions (Roche Diagnostics) using primers listed in Table 1 also with tester-cDNA as a template. Driver-cDNA probes were made by PCR labeling according to the manufacturer’s protocol using 5’ PCR primer (5’-AAGCAGTGGTATCAACGCAGAGT-3’) SmartTM cDNA library construction Kit.
RESULTS
The present study is ascertaining to know the Etroplus host and its bacterial pathogenesis through amenable genetic analysis. An infection disease is the manifestation through amenable genetic analysis. An infection disease is the manifestation of a dynamic series of event that occur between the host and pathogen that are defined by the interaction of pathogen-expressed virulent factors and the surveillance and defense systems of the host. Expression of both host and pathogen components is highly co-ordinated and the stimulus for any given response by the pathogen.
Subtractive hybridization: Tested of this bacterial genome by subtractive hybridization method (Fig. 1) using isogenic E. coli strains that differed only by the presence or absence of the 37 kh cag PAI. Which were equivalents about at 2% of the E. coli genome: strain 1480 wild type (cag+) was used as tester and Cag (C) (Calcium Channel Gene) (Huntington Diseased Gene) was used as a driver. Subtracted DNAs were cloned, the inserts were PCR amplified and products were spotted on hybridization filters. Clones specific to the cag PAI were then identified by probing these filters with labeled genomic DNAs from the cag tester strain and the cage driver and also with the set of DNA fragments that span the entire cag PAI (data not shown).
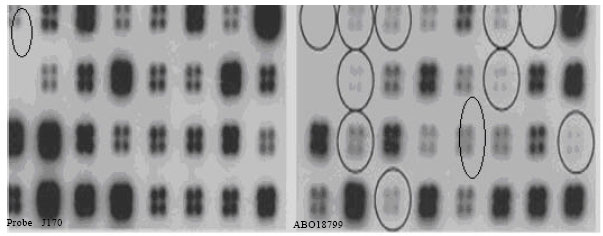 | |
| Fig. 1: | Representative dot-blot hybridization of products of subtractive hybridization by using genomic DNAs of E. coli strains J170 as tester and ABO18799 as driver. Cloned subtractive hybridization products were PCR-amplified using primers NPI and NP2 and spotted on Hybond N+ filters (Materials and Methods) and probe with labeled genomic DNAs, as indicated. The clones shown by DNA sequencing to have J170 DNAs either not present in the ABO18799 genome or substantially different from sequence in ABO18799 (as summarized in Table 3) are circled |
The hybridization results indicated that more than 90% of clones had derived from the cage PAI and 14 of them were one pass sequenced. This result indicates that many different DNAs specific to a given strain can be obtained by the subtractive hybridization method.
Two unrelated E. coli strains were tested next for differences in gene content, by using a strain well suited for rhesus monkey colonization (J 170) as tester and the strain whose genome was sequenced (26695) as driver. Each strain is plasmid free and it contains the cag PAI and the ice A1 gene. DNAs that recovered after subtraction were cloned. PCR amplified and arrayed for hybridization with labeled tester and driver DNAs as above. About 27 of 52 clones tested by hybridization were judged to contain strain J170 specific sequences, although the differences among clones were apparently less than in the first trial with isogenic cage of E. coli strains as will be explained. In contrast none of the 8 random clones (generated without subtractive hybridization) appeared to be specific (J170, 1480, 266695). Fourteen were unique three others were each represented twice which indicated that 17 different clones had been obtained. Nine of the cloned DNAs were found by sequencing to be absent or very divergent from those in the same direction on disappeared blank region of this liver and granulose cell.
Protein homologies: Eleven of the seventeen clones exhibited significant protein homologies with other databases entries, as summarized in Table 3. Five clones had homologies to part of putative restriction-modification (R-M) proteins. Two matched overlapping portions of type I specificity subunits (HsdS) shown in Table 3 but differed from one another size and sequence and thus must derive from different hsd5 genes three putative hsd 5 genes were found in the 16953 genome sequence ref. 29. Two clones had homology to Bcg-1 (Table 3) a two- subunit enzyme that is unusual in cleaving DNA at fixed distance on each side of the recognition site. Although quite difference BcgI homologs are encoded in strains 166943 (HP 1471; ref) PCR tests mapped these two types of homologs to the same locus. However the DNA sequence of the HP 1208 gene for J166 is known and does not contain the sequence found in clone F3. Thus the clones represent the different locus. The fifth clone exhibited protein homology to a type of II DNA methyltransferase. Two other clones exchibited homology to proteins that might be metabolic as well as immunoresponsive function. Five clones exhibited protein level homology to particular gene for hypothetical proteins of unknown homologs in other microbes. The remaining seven clones did not exchibit significant functions, two of which had equivalent function unknown protein homology to entries in current databases.
| Table 3: | Features of E. coli specific clones with significant database matches |
 | |
DISCUSSION
The present study is ascertaining to known the Etroplus host and its bacterial pathogenesis through amenable genetic analysis. Though, Fujiki et al. (2000) suggested a manifestation of the infectious diseases were a dynamic series of events mainly that occurred between the host and pathogen it might be defined as an interaction of pathogens expressed through virulence factors also depends upon the surveillance and resistant power of the host. Currently, Sriputhorn and Sanoamuang (2011) stated that the expression of both host and pathogen components is highly co-ordinated and the stimulus for any given response by the pathogen is often a prior change in defense gene expression by the host. This dynamic interplay determines the character, course and outcome of the pathogenic process. Much remains to be elucidated about host-pathogen interactions at the molecular level and the most informative model systems are likely to be those in which both the pathogen and host. Mainly, they were amenable to genetic systems for a wide variety of pathogenic microorganisms; rather less progress has been made in the host organisms that have traditionally been used to model infectious disease (Alonson and Leong, 2002). The present findings of this result also agreed by Bayne and Gerwick (2001) on immunological efficiency reflected gene content in another variety of the fish.
Recently an exciting approach has been to make use of the abilities of certain bacterial pathogens to infect nonmammalian model organisms with well-defined genetic systems, including the invertebrates Drosophilla melanogaster and Caenorhabdities elagans and the plant Arabidopsis thaliana. The ability of these models to mainly provide the infection of mammals derived from the fact that many fundamental host defense strategies. Velculescu et al. (1995) and Jorgensen et al. (2000) explained the evolutionarily ancient and are conserved diverse taxa are the microbial virulent factors, since they were used for evade from those defensive process. However, these model organisms lack defense systems that play important roles in mammalian host-pathogen interactions, including leukocytes, innate cellular immunity and adaptive immune systems.
Several technical aspects of this bacterial genome subtraction protocol merit attention. First the recovery of strain specific DNAs with unrelated strains (J140VS1780) was lower than that with isogenic control strains (cag+ and cag). This may reflect the base substitution and restriction fragment length differences between tester and driver DNAs which lower the efficiency with which driver DNAs titrate homologous tester sequences. Second half of J140-specific clone contained patches of sequence that were matched to those strain 1780. Such DNAs tend to be lost in other subtractive hybridization protocols (Brown and Curtiss, 1996; Tinsley and Nassif, 1996), although many of them could be important phenotypically (Coffey et al., 1995; Seifert, 1996). Their recovery here may reflect incomplete pairing with driver DNA which allows annealing with complementary tester DNA strands and thereby amplification (Fig. 1). In most of the cases, the nature and sizes of the DNA segments remain to be defined. Some will certainly represent members of divergent gene families that are mostly carried on E. coli strains (Savan and Sakai, 2002). Others may represent only a single gene that are strain specific in their distributions. A third important class may derive from much larger strain specific in their distribution of DNA segments such as PAIs and may contain new genes affecting colonization or diseases.
The present study also has demonstrated up-regulation of complement C7 and DRTP transcription by both RT-PCR and virtual Northern analysis. Many clones related to C7 complement and DRTP transcripts were sequenced in this SL5 library. A similar result was reported in rainbow trout liver library in response to Vibrio injestion with highest number of clones being for DRTP (n = 12) (Bayne and Gerwick, 2001; Ellis, 2001). The derived C7 and DRTP aminoacid sequences are 100% (AF281336). The derived C7 and DRTP aminoacid sequences are 100% (AF281336) and (AF81355) identical to rainbow trout sequences, respectively. Complement C7 is the part of the assembly of the late complement components to form a membrane attack complex (Janeway et al., 2001). The DRTP shares 34% sequences homology with CD59 which is complement inhibitor whose function is to protect against the lytic effect of the complement membrane attack complex (Lee and Goetz, 1998). The increase transcription of complement C7 and DRTP may balance one another and restore homeostasis during the APR to bacterial infection. Interestingly number of researchers discovered another detoxification related EST derived aminoacid sequence showed 26% identity to a novel soluble saxitoxin and tetrodotoxin binding plasma protein of puffer fish (Yotsu-Yamashita et al., 2001; Bayne and Gerwick, 2001) and common carp by Agouz and Anwer (2011).
Since the exact functions of both O-methyl transferases of SBP in fish are unknown, it will be very important to elucidate their role of infection distinguished by Hedge et al. (2000). Pathogens are often first recognized by virtue of their carbohydrate surfaces and a number of animal lectins are up regulated in infected salmon. Tachylectin was up regulated during infection in all three tissues tested (Aranishi and Mano, 2000). To date this fibrinogen like lectin has only been discovered in the Japanese horseshoe crab (Tachypleus tridentatus).
These results been showed that when the infection raised by any one of the pathogenic organisms the immune modulation of gene detected through the subtractive hybridization. Lectins from the C-type lectin super family were also represented in the subtracted libraries and three distinct C-type lectins were found to be up regulated in response to infection (Lund and Olafsen, 1998). While, C-type lectin 2-1 was originally isolated from carp and found to be constitutively expressed in liver and up-regulated in liver during the APR (Bayne et al., 2001; Alonson and Leong, 2002) and it specify an interesting difference in tissue specific modulation of expression between these two species. However, such mixtures of matching and divergent sequences are also potentially important evolutionarily in facilitating the formation of new genotypes by recombination during mixed infection and in thereby sometimes fascilitating quite dramatic changes in bacterial phenotype. Beyond these organisms that colonize inconstant and potentially hostile niches, such gene exchange can often be more potent than de novo mutation as adaptive mechanisms.
ACKNOWLEDGMENT
The authors gratefully acknowledged to our Malankara Catholic College Correspondent Fr. Prem Kumar (M.S.W) given encouragement and support for preparation of this research manuscript. We wish to extend the thanks to colleagues of S. Muthukumar, Dr. Francis Borgio, P.S. Sumi, Mary Sujin and Mary Jeeva for sample collection and all supports of this research study. The second author thankful to the Women Scientist Scheme (WOS-A), DST for financial assistance.
REFERENCES
- Agouz, H.M. and W. Anwer, 2011. Effect of biogen® and Myco-ad® on the growth performance of common carp (Cyprinus carpio) fed a mycotoxin contaminated aquafeed. J. Fish. Aquat. Sci., 6: 334-345.
CrossRefDirect Link - Alonson, M. and J. Leong, 2002. Suppressive subtraction libraries to identify interferon-inducible genes in fish. Mar. Biotechnol., 4: 74-80.
PubMed - Aranishi, F. and N. Mano, 2000. Antibacterial cathepsins in different types of ambicoloured Japanese flounder skin. Fish Shellfish Immunol., 10: 87-89.
PubMed - Aziz, D., S.S. Siraj, A. Arshad, S.M.N. Amin and S.A. Harmin, 2010. Population characterization of planktonic shrimp, Acetes japonicus (Decapoda: Sergestidae) using RAPD technique. J. Biol. Sci., 10: 355-361.
CrossRefDirect Link - Bayne, C.J. and L. Gerwick, 2001. The acute phase response and innate immunity of fish. Dev. Comp. Immunol., 25: 725-743.
CrossRef - Bobe, J. and F.W. Goetz, 2000. A tumor necrosis factor decoy receptor homologue is up-regulated in the brook trout (Salvelinus fontinalis) ovary at the completion of ovulation. Biol. Reporod., 62: 420-426.
PubMed - Bobe, J. and F.W. Goetz, 2000. Cysteine protease inhibitor is specifically expressed in pre- and early-vitellogenic oocytes from the brook trout periovulatory ovary. Mol. Reprod. Dev., 60: 312-318.
PubMed - Himmelreich, R., H. Plagens, H. Hilbert, B. Reiner and R. Herrmann, 1997. Comparative analysis of the genomes of the bacteria Mycoplasma Pneumoniae and Mycoplasma Genitalium. Nucl. Acids Res., 25: 701-712.
CrossRefDirect Link - Brown, P.K. and R. Curtiss, 1996. Unique chromosomal regions associated with virulence of an avian pathogenic Escherichia coli strain. Proc. Natl. Acad. Sci. USA., 93: 11149-11154.
Direct Link - Coffey, T.J., C.G. Dowson, M. Daniels and B.G. Spratt, 1995. Genetics and molecular biology of β-lactam-resistant Pneumococci. Microb. Drug Resist., 1: 29-34.
CrossRef - Ellis, A.E., 2001. Innate host defense mechanisms of fish against viruses and bacteria. Dev. Comp. Immunol., 25: 827-839.
CrossRefDirect Link - Gerwick, L., R. Steinhauer, S. Lapatra, T. Sandell, J. Ortuno, N. Hajiseyedjavadi and C.J. Bayne, 2002. The acute phase response of rainbow trout (Oncorhynchus mykiss) plasma proteins to viral, bacterial and fungal inflammatory agents. Fish Shellfish Immunol., 12: 229-242.
CrossRefDirect Link - Gorisman, E.A. and H. Ochman, 1996. Pathogenicity islands: Bacterial evolution in quantum leaps. Cell, 87: 791-794.
PubMed - Hedge, P., R. Qi, K. Abernathy, C. Gasy and S. Dharap et al., 2000. A concise guide to cDNA microarray analysis. Biotechniques, 29: 548-550.
PubMedDirect Link - Jorgensen, J.B., H. Lunde, L. Jensen, A.S. Whitehead and B. Robertson, 2000. Serum amyloid A transcription in Atlantic salmon hepatocytes is enhanced by stimulation with macrophage factors, recombinant human IL-1β, IL-6 and TNFα or bacterial lipopolysaccharide. Dev. Comp. Immunol., 24: 553-563.
CrossRef - Kanamori, A., 2000. Systematic identification of genes expressed during early oogenesis in medaka. Mol. Reprod. Dev., 55: 31-36.
CrossRef - Lund, V. and J.A. Olafsen, 1998. A comparative study of pentraxin-like proteins in different fish species. Dev. Comp. Immunol., 22: 185-194.
CrossRef - Pan, D., N. He, Z. Yang, H. Liu and X. Xu, 2005. Differential gene expression profile in hepatopancreas of WSSV-resistant shrimp (Penaeus japonicus) by suppression subtractive hybridization. Dev. Comp. Immunol., 29: 103-112.
PubMed - Rane, P.P. and S.S. Barve, 2011. Standardization and optimization of mtDNA isolation and molecular genetic analysis of D-loop region in animal natural fibres. Int. J. Zool. Res., 7: 190-200.
CrossRefDirect Link - Savan, R. and M. Sakai, 2002. Analysis of Expressed Sequence Tags (EST) obtained from common carp, Cyprinus carpio L., head kidney cells after stimulation by two mitogens, lipopolysaccharide and concanavalin-A. Comp. Biochem. Physiol. B, 131: 71-82.
CrossRef - Seifert, H.S., 1996. Questions about Gonococcal pilus phase and antigenic variation. Mol. Microbiol., 21: 433-440.
CrossRef - Shike, H., X. Lauth, M.E. Westerman, V.E. Ostland and J.M. Carlberg et al., 2002. Bass hepcidin is a novel antimicrobial peptide induced by bacterial challenge. Eur. J. Biochem., 269: 2232-2237.
CrossRefDirect Link - Sriputhorn, K. and L. Sanoamuang, 2011. Fairy shrimp (Streptocephalus sirindhornae) as live feed improve growth and carotenoid contents of giant freshwater prawn Macrobrachium rosenbergii. Int. J. Zool. Res., 7: 138-146.
CrossRefDirect Link - Tiffoche, C., A. Chesnel, P. Jego and J.P. Le Pennec, 1993. Isolation and characterization of a cDNA clone encoding Pleurodeles lectin. Eur. J. Biochem., 213: 901-907.
CrossRefDirect Link - Tinsley, C.R. and X. Nassif, 1996. Analysis of the genetic differences between Neisseria meningitidis and Neisseria gonorrhoeae: Two closely related bacteria expressing two different pathogenicities. Proc. Natl. Acad. Sci. USA., 93: 11109-11114.
Direct Link - Van Hal, N.LW., O. Vorst, A.M.M.L. van Houwelingen, E.J. Kok and A. Peijnenburg et al., 2000. The application of DNA microarrays in gene expression analysis. J. Biotechnol., 78: 271-280.
CrossRef - Velculescu, V.E., L. Zhang, B. Vogelstein and K.W. Kinzler, 1995. Serial analysis of gene expression. Science, 270: 484-487.
CrossRefDirect Link - Waldor, M.K. and J.J. Mekalamos, 1996. Lysogenic conversion by a filamentous phage encoding cholera toxin. Science, 212: 1910-1914.
PubMed - Yotsu-Yamashita, M., A. Sugimoto, T. Terakawa, Y. Shoji, T. Miyazawa and T. Yasumoto, 2001. Purification, characterization and cDNA cloning of a novel soluble saxitoxin and tetrodotoxin binding protein from plasma of the puffer fish, Fugu pardalis. Eur. J. Biochem., 268: 5937-5946.
PubMed - James, R., N. Thampuran, K.V. Lalitha, L.A. Rajan and T.C. Joseph, 2010. Differential gene expression profile of the hepatopancreas of white spot syndrome virus infected Fenneropenaeus indicus by suppression subtractive hybridization. Fish Shellfish Immunol., 29: 884-889.
CrossRef - Thangaraj, M., V. Prem, T. Ramesh and A.P. Lipton, 2011. RAPD fingerprinting and demonstration of genetic variation in three pathogens isolated from mangrove environment. Asian J. Biotechnol., 3: 269-274.
CrossRefDirect Link