
Priyanka P. Rane
KETs V.G. Vaze College of Arts, Science and Commerce, Mithagar Road, Mulund (E), 81, Mumbai, India
S. S. Barve
KET`s V.G. Vaze College of Arts, Science and Commerce, Mithagar Road, Mulund (E), 81, Mumbai, India
International Journal of Zoological Research
Year: 2011 | Volume: 7 | Issue: 2 | Page No.: 190-200







ABSTRACT
Increase in demand for animal natural fibres in recent years for the production of high quality textile products has resulted in the adulteration and false declaration of these fibres causing heavy financial loss. Fibres are expensive due to limited feedstock and less fibre production. To keep up with the demand these fibres are adulterated with less expensive fibres viz., wool to give special effect to the fabric. To control false declaration, there is a need for fibre identification and to ascertain blend composition. Though Scanning Electron Microscopy is generally used for fibre analysis but this method is time consuming, expensive and the reliability of results depend on the expertise of the microscopist. Hence, there is a need for reliable and economical method to characterize these fibres and to study composition of each animal fibre in blends. The aim of the present study was mitochondrial DNA extraction from animal natural fibres in untreated and blends. The modified protocol includes addition of Proteinase K, Dithioerythritol individually in each tube and final extraction with phenol: chloroform: isoamyl alcohol, amplification of D-loop region using species specific and mammalian specific primers. We observed that with species specific primers, it was possible to study inter species variation but the blends could be detected if there was prior knowledge about the fibres in blends. With mammalian specific primers we could study blends and differentiate between fibres from sheep breeds but inter species variation was difficult. It can be concluded that mtDNA analysis can be used to differentiate animal fibres and control adulteration.
PDF Abstract XML References Citation
How to cite this article
Priyanka P. Rane and S. S. Barve, 2011. Standardization and Optimization of mtDNA Isolation and Molecular Genetic Analysis of D-loop Region in Animal Natural Fibres. International Journal of Zoological Research, 7: 190-200.
DOI: 10.3923/ijzr.2011.190.200
URL: https://scialert.net/abstract/?doi=ijzr.2011.190.200
DOI: 10.3923/ijzr.2011.190.200
URL: https://scialert.net/abstract/?doi=ijzr.2011.190.200
INTRODUCTION
Animal hair fibres are based on cross-linked proteins known as keratins (Hamlyn, 1997). Animal hair has three region viz. root, shaft and tip. Hair shaft consists of an outer cuticle surrounding a central cortex which may contain a central medulla (Harkey, 1993). DNA is present in all fibre cell types (Cortical cells-ortho and para; cuticle cells-exo, endo and membranous epicuticle) and remains intact as nuclear remnants after the fibre has keratinized (Maclaren and Milligan, 1981; Nelson et al., 1996). Hair contains both nuclear DNA (when root present) and mitochondrial DNA (Sahajpal et al., 2009). During the process of keratinisation the cells become metabolically inactive; however remnants of the nuclear material containing DNA remain trapped inside the fibre. The recalcitrant and waterproof nature of fibres helps to protect the DNA from chemical and biological degradation (Hamlyn, 1997).
Mitochondrial DNA (mtDNA) is located in the mitochondria that are present in the cells of the fibre. mtDNA is circular, is maternally inherited and present in 1,000-10,000 copies per mammalian cell. mtDNA is composed of two regions viz. control (noncoding) and coding region. The control region consists of two variable segments, hypervariable region 1 (HV1) and hypervariable region 2 (HV2). The length of control region/D-Loop (displacement -loop) region is different which leads to difference in size of mtDNA molecules. The length can vary both between individuals and between different mtDNA molecules within an individual. It is an extremely useful genetic marker when only a minute amount of DNA is available (Angleby, 2005). It is the only nucleic acid that can be recovered from extremely small or very old or degraded samples. mtDNA extraction is frequently performed on difficult samples such as of tissues, hair, bones and teeth. These tissues do not have sufficient nuclear DNA for analysis, but rich source of mtDNA (Carlson et al., 2004).
Hair fibres of Cashmere goat, Angora goat, Camel, Angora rabbit fur are called as speciality fibres or luxury fibres and identified as cashmere wool, mohair, camel hair Angora rabbit fur respectively. These fibres are precious animal fibres and of commercial importance other than sheep wool. They are known as luxury fibres due to their softness, luster and scarcity relative to other fibres (Hamlyn, 1997). Fibre identification was mainly done by Light microscopy, Scanning Electron Microscopy (SEM) but the method was less reliable due to human errors. The method was time consuming, much costly and the reliability and authenticity of results depended on the expertise of the microscopist and the degree of fiber processing (Hocker, 1990). However microscopic studies suffer from observational biases and are ineffective at low sample sizes and require corroboration with other studies (Hamlyn et al., 1995). Due to these reasons and similarity in fineness (microns) it is difficult to differentiate them microscopically (Fig. 1a-c) and often leading to adulteration of these fibres with wool. Wortmann and Wortmann, 1988 showed that microscopically Cashmere and Wool showed surface similarities creating problem in differentiation while studying Cashmere/Wool blends. According to Hocker (1990) differentiation of Mohair and Wool was a major problem; also there was problem in identification of cashmere and its differentiation from yak hair and Camel from other cameloids.
Speciality fibres are of great importance in textile industry and its demand is also increasing but due to restricted availability and high prices, adulteration and false declaration are common (Kerkhoff et al., 2009). Qualitative and quantitative methods are required for the characterization of speciality animal fibres (raw and processed), not only to satisfy legal requirements concerning textile labeling but to assist processors to define and control blend composition (Nelson et al., 1990).
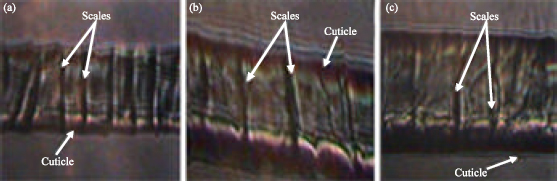 | |
| Fig. 1: | CCD micrographs of Hair fibres of (a) Ovis aries, (Wool), (b) Capra hircus laniger (Cashmere) and (c) Capra hircus aegagrus (Mohair) under 40x magnification. Figures show the scale pattern of the animal fibres and the cuticle |
Mitochondrial DNA analysis is a reliable and authentic method to satisfy all the limitations faced till today during analysis of both processed as well as raw fibres. Kerkhoff et al. (2009) studied genomic DNA of speciality fibres for differentiation of species with species specific primers and Ka-Man et al. (2009) showed the usefulness of universal primers for mammals for identification of different species by studying their mitochondrial DNA control region. In the present study we report here the extraction of mitochondrial DNA from hair shafts of animal natural fibres (individual species as well as blends of samples), amplification of the D-Loop region of the mtDNA using species specific as well as mammalian specific primers (universal primer for mammals) and to study the possibility of differentiation of animals as individual species as well as in blends.
DNA analysis is a useful tool in forensic analysis and identification (Carlson et al., 2004), speciality fibre identification for textile labeling (Suppiger, 2004; Hamlyn, 1997; Kerkhoff et al., 2009; Kalbe et al., 1988), dealing wildlife offence (Sahajpal, 2006; Sahajpal et al., 2009), ancient DNA analysis from extinct species (Gilbert et al., 2007), genetic diversity (Hofreiter et al., 2002), phylogenetic analysis (Grzybowski et al., 2003).
MATERIALS AND METHODS
The present study was conducted from January 2008 to June 2010.
Sample preparation: Hair samples were collected from the animals mentioned in Table 1 during their molting and shearing period (wool: sheared once a year in spring, cashmere: shed in spring, mohair: shorn twice a year in spring and fall, Angora fur: molt every 4 months, camel: shed in late spring to early summer) at random from the farms and traders in India wherein the history of the animal (i.e., nutrition, health, age, sex etc.) and the fibre (i.e., period of storage after shearing/moulting) was not known except that all the fibres collected were in raw state, i.e., without any prior chemical treatment. Hair sample was rinsed with anionic surface active detergent to remove suint, dirt and excretory matter, air dried completely and then used for extraction.
Extraction of mtDNA from animal hair: mtDNA was extracted by following the method of Gilbert et al. (2007) with modifications. 0.3 g of each sample was weighed. A blend of all samples was prepared in ratio 5: 1: 1: 1: 1: 1 (Australian wool in major quantity in comparison to other speciality fibres as this is the commercial trend followed in preparing blends). Hair fibres were cut in fine pieces with sterile scissors and washed with Isopropanol for ½ h. The hairs were air-dried and extraction buffer containing 10 mM Tris Cl pH 8.0, 2.5 mM EDTA pH 8.0, 10 mM NaCl, 5 mM CaCl2, 2% SDS (w/v), was added to each tube. Two hundred microliter of 40 mM Dithioerythritol (DTE) (Sisco Research Laboratories, SRL) and 160 μL Proteinase K (20 mg mL-1) (Himedia) was individually added to each tube and incubated at 56°C for 24 h on a shaker at 60 rpm. Final extraction was done with equal volumes of phenol: chloroform: isoamyl alcohol (25: 24: 1) and centrifuged at 14,000 rpm for 10 min at 30°C.
| Table 1: | List of animals from which hair samples were obtained |
 | |
Aqueous layer was collected carefully in a clean glass tube so that it was free of hair fibres and twice extracted with equal volumes of chloroform to remove residual phenol followed by centrifugation at 14,000 rpm for 10 min at 30°C. To the aqueous layer 0.5 volumes of chilled Isopropanol was added and mixed gently but thoroughly and placed in the freezer overnight. DNA precipitated as scattered thin fragments. Spin at 14,000 rpm for 10 min at 4°C. Pellet was washed with 70% ethanol followed by centrifugation at 14,000 rpm for 5 min at 4°C. Supernatant was slowly decanted without disturbing the pellet. Pellet was allowed to dry overnight at room temperature. Pellet was dissolved completely in 0.1 X TE buffer and stored at -40°C.
Purity and Electrophoresis of mtDNA: Extracted DNA was electrophoresed on 1.2% Agarose gel with internal Ethidium bromide staining at 50V for 1½ h. The purity of the extracted mtDNA was also checked by measuring absorbance on Varian Cary 50 UV spectrophotometer at 260 and 280 nm. Purity was analyzed by absorbance ratios, i.e., A 260/280 nm.
Mitochondrial DNA D-Loop amplification: D-Loop region of the extracted mtDNA was amplified using species specific primers (sheep, goat, camel and rabbit) and mammalian specific primers specific to mtDNA D-Loop region as given in Table 2. Primers were synthesized from Merck Specialities Private Limited (Bangalore Genei), India. All primers were reconstituted in sterile deionized water as 100 μM stock and were stored at -20°C.
Polymerase Chain Reaction (PCR) reaction was carried out with the final volume of 25 μL which contained 2.5 μL of DNA template, 2.5 μL 1X assay buffer containing MgCl2, 1 μL of 10 mM dNTPs mix, 2 μL of each primer and 0.6 μL of 1.2 U Taq DNA polymerase containing 20 mM Tris HCl (pH 8.0), 100 mM KCl, 0.1 M EDTA, 1 M DTT, 0.5% Tween-20 and 50% glycerol and final volume was adjusted with deionized water. PCR amplification was set up in the 2720 Thermal Cycler, Applied Biosystems� USA and cycle profile for the control region (D-Loop) were initial denaturation at 95°C (5 min), followed by 40 cycles of 0.15 sec at 95°C (denaturation), 1 min at 60°C (annealing) and 2 min at 72°C (extension). Final extension at 72°C (7 min) and hold temperature was maintained at 4°C. After amplification, PCR products were stored at 4°C till electrophoresis. PCR products were mixed with 2.5 mL of 10X loading dye (0.25% bromophenol blue, 0.25% xylene cyanol and 40% sucrose, w/v) and electrophorosed in 2% Agarose gel with internal ethidium bromide staining (10 μg mL-1) in 1X TAE buffer at 100V for half an hour.
| Table 2: | Sequence of species specific and mammalian specific primers specific to mtDNA D-loop |
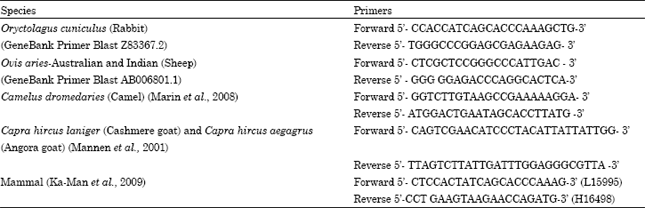 | |
A 100 bp DNA ladder was used as a standard molecular weight marker.
RESULTS AND DISCUSSION
mtDNA analysis: Hair fibres used for extraction were collected in raw state during the moulting and shearing periods and stored for long time at room temperature in clean dry place. Schuster and Miller (2007) reported that hair samples from Mammoth stored at room temperature for 200 years allowed complete analysis of its mitochondrial genome. Hair fibres were washed with anionic surface active detergent to remove external dirt, suint, excretory matter etc. Raw wool often contains wool grease, suint, dirt and vegetable matter (Teasdale, 1988; Rippon, 1992). It was observed that hair shafts can be readily decontaminated from external sources of contamination, due to their highly keratinized structure and relative lack of porosity in comparison to more conventional sources of a DNA such as bone or tooth (Gilbert et al., 2007). DNA is protected by keratin in hair (Smith, 2007) and even washing in a solution that kills and washes off external DNA has no effect on internal DNA (Schuster and Miller, 2007).
Hair contains both nuclear (when root present) and mitochondrial DNA (Sahajpal et al., 2009). Since hair fibres taken for analysis had no roots so mtDNA analysis was undertaken. Extraction of mtDNA from hair is exceptionally difficult (Carlson et al., 2004). The epicuticle of wool consists of lipids, proteins and/or carbohydrates (Hocker, 2002), is strongly hydrophobic and forms a resistant barrier (Lofts and Truter, 1969; Pailthorpe, 1992) to attack from alkalis, oxidizing agents and proteolytic enzymes. This makes the penetration of extraction buffer into the fibre extremely difficult. To isolate good and pure mtDNA from hair fibres we made few modifications in the extraction procedure of Gilbert et al. (2007). (1) Sample volume is critical when isolating mtDNA from hair shafts. With lesser quantity of fibres (0.05-0.2 g) no pellet formation was observed and increase in quantity (0.5 g and above) increased the relative amount of extraction buffer and other chemicals, thus increasing the chances of contamination. So, 0.3 g sample was taken for extraction. (2) Hair fibres were treated with Isopropanol for cleaning the surface to remove surface lipids increasing wettability of fiber and to remove other contaminants present and for fibre swelling to enhance the penetration of extraction buffer into the fibre. Zhang et al. (2009) used ethanol to clean the hairs free of fecal matter. Polar solvents such as alcohols cause swelling of the fibre and also act as surface-active agent to facilitate surface tension (Tímar-Balazsy and Eastop, 1998). (3) DTE an epimer of DTT was used as reducing agent to cleave disulphide bonds of proteins instead of Dithiothreitol (DTT). (4) Incubation temperature was optimized at 56°C as the activity of Proteinase K increased on raising the temperature from 37 to 50-60°C. (5) 160 μL 20 mg mL-1 Proteinase K was used instead of 10% Proteinase K which is used to digest proteins. (6) Proteinase K and Dithioerythritol (DTE) were added individually to each tube which gave better DNA yield than with addition in extraction buffer. Sahajpal et al. (2009), Miscicka-Sliwka et al. (1997), Zhang et al. (2009) and Kolomyjec (2007) used DTT for extraction of mtDNA from hair samples of wildlife species, human, tiger, respectively. (7) We used Phenol: chloroform: isoamyl alcohol (25:24:1) followed by repeated chloroform washings (Miscicka-Sliwka et al., 1997; Carlson et al., 2004) and followed by alcoholic precipitation instead of using microconcentrators. It was observed that Phenol: chloroform: isoamyl alcohol removed maximum amount of the proteineaceous material and hair residues than just phenol: chloroform or just phenol. Use of DNA microconcentrators was avoided to develop a cost effective method of extraction. Hocker (1990), Nelson et al. (1996) and Kerkhoff et al. (2009) used phenol: chloroform (1:1) for final extraction, Gilbert et al. (2007) used phenol followed by chloroform washing and Kolomyjec (2007) used chloroform: isoamyl alcohol (24: 1). 8. Centrifugation period and speed was optimized at 14,000 rpm for 5-10 min. There was no pellet formation below 12,500 rpm. Use of ultracentrifugation enabled us maximum recovery of pellet. 9. Isopropanol was used to precipitate DNA and pellet washed with 70% (v/v) ethanol and redissolved in TE buffer (Hamlyn et al., 1996).
We were not able to visualize DNA on 1.2% Agarose gel this was in agreement with the findings of Sahajpal, 2006 according to which the extracted DNA from hair could not be visualized due to low concentration. Swift (1977) reported that wool fibre is generally small in diameter (10-30 μm) and has a single layer of cuticle cells compared to human hair which is 60-100 μm in diameter and has 6-10 layers of overlapping cuticle cells. According to Miscicka-Sliwka et al. (1997) human hair shaft contain 1 to 1.5 ng only. This shows that DNA extracted from speciality fibres will be less than 1 to 1.5 ng. Studies by Kolomyjec (2007) and Oliveira et al. (2007) also showed that it was not possible to observe DNA on Agarose gel without amplification. Hence we checked the purity of the extracted DNA using UV Spectrophotometer. We found that ratio of A 260/280 nm was 1.85 indicating that there was no RNA or Protein contamination associated with DNA.
D-loop analysis: For amplification of extracted mtDNA D-loop region, species specific primers (goat, sheep, rabbit and camel) (NCBI primers blast Z83367.2; NCBI primers blast AB006801.1; Mannen et al. (2001) and Ka-Man et al. (2009) and mammalian specific primers were designed (Marin et al., 2008). To get sharp and reproducible amplicons we also optimized various PCR conditions such as annealing temperature, amount of DNA, concentration of Taq polymerase etc. When 0.6 U of Taq Polymerase was used gave feeble bands and 1.2 U gave sharp band with 1 and 2.5 μL of DNA template. Though at 1 μL of template DNA, amplicons were observed but were not reproducible. On the other hand with 2.5 μL DNA sharp bands were obtained. Hence for further experiments 1.2 U Taq DNA polymerase and 2.5 μL DNA template was used. Similarly when 55°C was used as annealing temperature did not give sharp amplicons but at 60°C gave sharp amplicons (Fig. 2). On electrophoresis of amplified product we observed that amplicons were off very low molecular weight hence electrophoresis was carried out for shorter duration.
D-Loop region of the extracted mtDNA from individual samples and blends was amplified with species specific primers (sheep, goat, camel and rabbit) and universal primer for mammal using the standardized protocol. The amplicons were subjected to electrophoresis for 1 h showed following results.
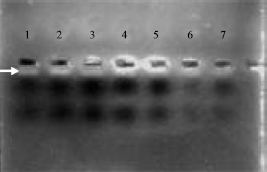 | |
| Fig. 2: | Electrophoresis of amplified product after half an hour using 0.6U Taq polymerase and 55°C annealing temperature. Lane 1: Ovis aries (Australian), lane 2: Ovis aries (Indian), lane 3: Capra hircus laniger, lane 4: Capra hircus aegagrus, lane 5: Oryctolagus cuniculus, lane 6: Camelus dromedaries and lane 7: Blends of all sample |
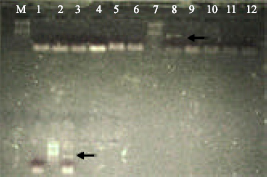 | |
| Fig. 3: | Electrophoresis of amplification with species specific primer (Lane 1 to 6) and mammalian specific primer (Lane 7 to 13). Species specific primers: Lane 1: Oryctolagus cuniculus, Lane 2: Ovis aries (Australian), Lane 3: Ovis aries (Indian), Lane 4: Camelus dromedaries, Lane 5: Capra hircus laniger, Lane 6: Capra hircus aegagrus, Mammalian specific primers: Lane 7: Ovis aries (Australian), Lane 8: Ovis aries (Indian), Lane 9: Capra hircus laniger, Lane 10: Capra hircus aegagrus, Lane 11: Oryctolagus cuniculus and Lane 12: Camelus dromedaries |
Figure 3 (Lane 1-6) showed the results for amplification using species specific primers. A single amplicon was produced for Oryctolagus cuniculus (Lane 1) using rabbit specific primers, for Ovis aries (Australian and Indian, respectively) (Lane 2, 3) using sheep specific primer, for Camelus dromedaries (Lane 4) using camel specific primers. A single amplicon of slightly high molecular weight than that for Ovis was produced for Capra hircus laniger and Capra hircus aegagrus (Lane 5, 6), respectively. It was observed that with species specific primers inter species variation could be studied between Ovis and Capra species but intra species (between Cashmere and mohair, sheep breeds) variation was difficult to study. Blends could be studied only if there was a prior knowledge of species present in mixture (Ka-Man et al., 2009). Kerkhoff et al. (2009) showed that discrimination of different breeds of goat (cashmere and mohair) was not possible with amplification using species specific primer as this was only possible with RAPD, AFLP. But we have used hair shafts without roots so possibility of high molecular DNA was little and so RAPD cannot be used (Sahajpal et al., 2009).
Figure 3 (Lane 7-13) showed results for amplification using mammalian specific primer for different species as well as blends. Ovis aries (Australian) (Lane 7) produced two amplicons while Ovis aries (Indian) (Lane 8) produced only a single amplicon. The results obtained through the preliminary study show that mammalian specific primer could differentiate between Australian and Indian breeds. Capra hircus laniger, Capra hircus aegagrus, Oryctolagus cuniculus, Camelus dromedaries (Lane 9, 10, 11, 12, respectively) produced only a single amplicon. Blends of all samples (Lane 13) produced two amplicons similar to Ovis aries (Australian) (Lane 7), the amplicon of high molecular weight is only present in Ovis aries (Australian) and absent in all other samples, but present in blends of samples. This showed that mammalian specific primer could be used to study blends and find out the contamination of wool in speciality fibres. Ka-Man et al. (2009) showed that use of universal primer to identify single species as well as mixture of two or three species especially when the origin of sample is unknown. According to his study PCR fragments could be easily separated for all species in sample containing mixture of two species on Agarose gel if their difference in size was ≥20 and ≤300 bp. This difference is due to the resolution power and sensitivity of the technique. Figure 4 showed the amplification pattern for blends of all samples using species specific as well as mammalian specific primers.
 | |
| Fig. 4: | Blends of sample with different primers- Lane 1: Mammalian specific primers, Lane 2: Rabbit specific primer, Lane 3: Sheep specific primer, Lane 4: Camel specific primer, Lane 5: Goat specific primer, Lane 6: Goat specific primer and Lane 7: Mammalian specific primers |
It confirmed the observation that with mammalian specific primer study of blends was possible.
Similar type of work has been carried out by Hamlyn (1997), Hocker (1990) and Nelson et al. (1996) to study high molecular weight DNA from speciality fibres. Suppiger, 2004 studied mtDNA D-Loop region in Cashmere goat and fibre producing goats by sequencing the DNA to distinguish between fibre producing goats and halt false declaration in textiles. Li et al. (2007) studied D-loop in Tibetian sheep, Wu et al. (2003) studied mitochondrial control region in argali wild sheep. Kalbe et al. (1988) studied high molecular mass DNA from human as well as alpaca, angora rabbit, cashmere, cashgora, mohair, merino and yak to identify species. Zhang et al. (2009) studied mtDNA from feces and hair shaft of South China tiger, Huhne et al. (1999) studied mtDNA in human hair shaft.
CONCLUSIONS
In conclusion we found that DNA analysis can be a useful, reliable and authentic tool to study animal natural fibres along with other methods like Polyacrylamide Gel Electrophoresis (PAGE), Isoelectric Focusing (IEF) or even Electron microscopy. With the data obtained after amplification of mtDNA D-Loop region, we observed that individual animal hair fibre or blends could be studied. It was found that with species specific primer it was possible to study inter species variation in animals while intra species variation was difficult. Blends could be studied only if the origin of sample in the blends was known. Mammalian specific primer can be used to study blends and detect wool contamination in speciality fibres. Also, it may help to study intra species variation in sheep breeds. Sequencing of the D-Loop region becomes necessary when visual difference in the molecular weight of the bands obtained through the amplification is impossible. This will help to give sound proof regarding the difference in the DNA sequence of individual animal and helping in differentiating them when present as blends to control adulteration and false declaration of the fibres.
ACKNOWLEDGMENT
I would like to thank Dr. Abhijit Sahasrabudhe who helped me in PCR analysis and Reliable Analytical laboratories, Thane, Maharashtra, India for providing the lab facility for PCR.
REFERENCES
- Miscicka-Sliwka, D., G. Tomasz, M. Wozniak and J. Czarny, 1997. Forensic DNA typing of a single hair:mitochondrial DNA sequencing and highly discriminating STR multiplexes developed for various detection platforms. Proceeding of the 8th International Symposium on Human Identification, (ISHI`97), Promega Corporation, Madison, pp: 132-142.
- Grzybowski, T., B. Malyarchuk, J. Czarny, D. Miscicka-Sliwka and R. Kotzbach, 2003. High levels of mitochondrial DNA heteroplasmy in single hair roots: Reanalysis and revision. Electrophoresis, 24: 1159-1165.
PubMed - Hamlyn, P.F., S. Ramsbottom, B.J. McCarthy and G. Nelson, 1996. Analysis of Speciality Fibres using DNA Amplification Techniques, in Metrology and Identification of Speciality Animal Fibres. In: European Fine Fibre Network, Laker, J.P. and F.J. Wortmann (Eds.). Macaulay Land Use Research Institute, Aberdeen, Scotland, pp: 59-68.
- Hofreiter, M., C. Capelli, M. Krings, L. Waits and N. Conard et al., 2002. Ancient DNA analyses reveal high mitochondrial DNA sequence diversity and parallel morphological evolution of late pleistocene cave bears. Mol. Biol. Evol., 19: 1244-1250.
Direct Link - Huhne, J., H. Pfeiffer, K. Waterkamp and B. Brinkmann, 1999. Mitochondrial DNA in human hair shafts-existence of intra-individual differences. Int. J. Legal Med., 112: 172-175.
PubMed - Mannen, H., Y. Nagata and S. Tsuji, 2001. Mitochondrial DNA reveal that domestic Goat (Capra hircus) are genetically affected by two subspecies of bezoar (Capra aegagurus). Biochem. Genet., 39: 145-154.
CrossRefPubMedDirect Link