
Imtiyaz Hussain Mir
Department of Zoology, University of Kashmir, Srinagar-190006 India
Ashok Channa
Department of Zoology, University of Kashmir, Srinagar-190006 India
Sumaira Nabi
Department of Zoology, University of Kashmir, Srinagar-190006 India
International Journal of Zoological Research
Year: 2011 | Volume: 7 | Issue: 1 | Page No.: 100-106







ABSTRACT
Liver samples of the snow trout, Schizothorax curvifrons have been studied with the aid of Transmission Electron Microscope (TEM) after carrying out their primary fixation in Karnovsky’s fluid buffered with 0.1 M sodium phosphate buffer and secondary fixation in 1% osmium tetraoxide in 0.1 M sodium phosphate buffer with the objectives of describing the normal subcellular structure of hepatic tissue and intrahepatic exocrine pancreatic tissue, setting thereby a strong platform for further studies in relation to pollution, stress conditions in aquaculture, natural and polluted environments and pathobiology of teleost liver. The structural analysis reveals that the hepatocytes possess a centrally located nucleus with distinct nucleolus and heterochromatin situated close to the porous karyolemma. The cisternae of Granular Endoplasmic Reticulum (GER) are arranged in an array parallel to the karyolemma and the mitochondria varying in shape from circular to elongate are found in close association of the Granular Endoplasmic Reticulum (GER), besides a good content of scattered glycogen occupies major part of the cytoplasm of the hepatocytes. The intrahepatic exocrine pancreatic tissue diffused in the hepatic parenchyma exhibit a uniform density of secretory granules at the apical portion, thereby displacing the nucleus towards the basal region of the cell.
PDF Abstract XML References Citation
Received: May 18, 2010;
Accepted: June 29, 2010;
Published: August 21, 2010
How to cite this article
Imtiyaz Hussain Mir, Ashok Channa and Sumaira Nabi, 2011. Ultrastructural Analysis of the Liver of the Snow Trout, Schizothorax curvifrons Heckel. International Journal of Zoological Research, 7: 100-106.
DOI: 10.3923/ijzr.2011.100.106
URL: https://scialert.net/abstract/?doi=ijzr.2011.100.106
DOI: 10.3923/ijzr.2011.100.106
URL: https://scialert.net/abstract/?doi=ijzr.2011.100.106
INTRODUCTION
The teleost liver is a bi or tri lobed dense organ ventrally located in the cranial region of the visceral cavity. Its shape size and volume are adapted to the space available between other visceral organs. The hepatic ultrastructure of teleosts responds very sensitively to environmental changes like, in temperature, seasons, feeding conditions or presence of various chemicals in the water (Storch and Juario, 1983). Attempts have been made to use this responsiveness as an indicator for aquatic pollution effects (Pierce et al., 1978, 1980; Haensly et al., 1982). It is becoming increasingly apparent from studies with fish (Segner and Moller, 1984; Weis, 1974; Hugla and Thome, 1999; Studnicka, 1983) and other vertebrates (Arstila et al., 1974) that morphological alterations associated with sublethal injury are commonly encountered at the subcellular level.
The snow trout, Schizothorax curvifrons locally called as Satter Gad is a prized indigenous herbivorous cold freshwater teleost of Kashmir valley whose population is at decline due to multiple factors. The fish belonging to the family Cyprinidae proves to be morphometrically, meristically and economically most variable and valuable promising food species of the paradise dale and can always be recognized by the combination of large high scale count, high gill raker number and thin lips (Mir and Channa, 2009, 2010). The liver is of utmost physiological importance as it is the chief site for storage of high energy foods (glycogen, lipids) which utilizes the fish at the time of emergency and it also plays a vital role in detoxification, digestion and other physiological processes (Channa and Mir, 2009). There is no published account on the transmission electron microscopic studies of the fishes of Kashmir in general and Schizothorax in particular. So, the present study has been undertaken for the first time as an effort to carry out the ultrastructure of the unaltered liver of a cold freshwater teleost, Schizothorax curvifrons with the intend that this study will provide a base line for future studies in relation to pollution, stress conditions in aquaculture, natural and polluted environments and pathobiology of teleost liver.
MATERIALS AND METHODS
Living specimens of normal and healthy snow trout, Schizothorax curvifrons were caught from their natural habitat. Immediately after catching, the fishes with a body length of 25-30 cm and body weight 300-350 g were killed with a blow to the head and dissected. Following dissection, small blocks of liver of about 1 mm3 were fixed in modified Karnovsky’s fluid (2.5% glutaraldehyde and 4.0% paraformaldehyde) buffered with 0.1 M sodium phosphate buffer (pH 7.4) for 16 h at 4°C temperature (Primary fixation). Following primary fixation the tissue samples were washed in fresh 0.1 M sodium phosphate buffer and post fixed in 1% osmium tetraoxide in 0.1 M sodium phosphate buffer (pH 7.4) for 2 h at 4°C (Secondary fixation). The post fixed samples were washed several times in fresh 0.1 M sodium phosphate buffer, dehydrated in graded acetone solutions and embedded in CY212 araldite. Ultrathin sections of 60-80 nm thickness were cut using an ultracut (Reichert Jung) ultramicrotome and transported to grids, contrasted with uranyl acetate and lead citrate, analyzed and documented photographically in a Philips CM10 Transmission Electron Microscope (TEM) operated at 60-80 kv accelerating voltage at All India Institute of Medical Sciences (AIIMS) New Delhi, India. This study was conducted from 2007 to 2008.
RESULTS AND DISCUSSION
Hepatocytes of mature herbivorous snow trout, Schizothorax curvifrons under transmission electron microscope reveal a centrally located nucleus of high electrodensity with distinct nucleoli (Fig. 1). The nucleus has granular chromatin with more condensed heterochromatin situated close to the karyolemma and the pores through which exchange of materials takes place between karyoplasm and cytoplasm of the hepatocytes are quite distinct in the double layered karyolemma (Fig. 2).
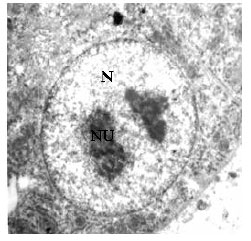 | |
| Fig. 1: | Fine structure of the hepatocyte showing centrally placed round nucleus (N) with nucleolous (NU) X3500 |
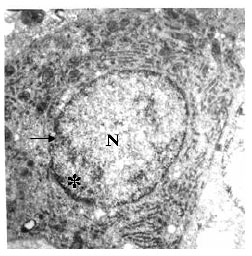 | |
| Fig. 2: | Transmission electron micrograph of the hepatocyte showing nucleus (N) with peripheral heterochromatin (*) and distinct nuclear pores (→) in the nuclear membrane. X3400 |
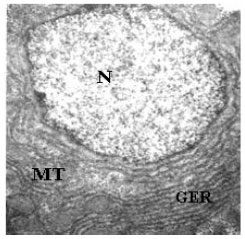 | |
| Fig. 3: | TEM of hepatocyte showing a nucleus (N), mitochondria (MT) and stacks of parallel cisternae of the granular endoplasmic reticulum (GER). X4500 |
Cisternae of Granular Endoplasmic Reticulum (GER) arranged in stacks in the cytoplasm embrace the nucleus as well as each mitochondria varying in shape from circular to elongate (Fig. 3, 4). A large amount of scattered glycogen fills major part of the cytoplasm of the hepatocytes (Fig. 5). The intrahepatic exocrine pancreatic cells of Schizothorax curvifrons are easily differentiated from the other cell types by the presence of large number of secretory granules, being uniform in density (Fig. 6). The secretory granules are limited by a single membrane and are usually located at the apical portion of the cell in addition to the granular endoplasmic reticulum (GER), thereby displacing the spherical nucleus towards the basal portion of the cell.
A high degree of interspecific variation occurs among teleostei. This variation underlines the importance of determining hepatic ultrastructure for individual fish species. Gonzalez et al. (1993) and Brusle and Gonzalez (1996) have reported that the hepatocytes of the fish liver are relatively poor in organelles, indicating a low synthetic activity for secretory proteins. The present investigation on the TEM studies of the liver show the presence of centrally placed nucleus with two prominent nucleoli and distinct nuclear pores in the double nuclear membrane.
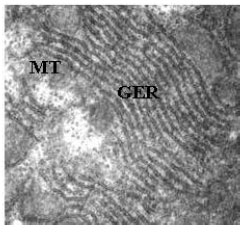 | |
| Fig. 4: | Transmission electron micrograph showing mitochondria (MT) in close association with the granular endoplasmic reticulum (GER) in the cytoplasm of the hepatocyte. X6000 |
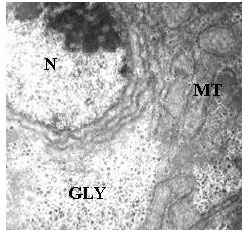 | |
| Fig. 5: | Electron micrograph of the hepatocyte showing nucleus (N), abundance of mitochondria (MT) and a good content of glycogen (GLY) scattered throughout the cytoplasm. X5000 |
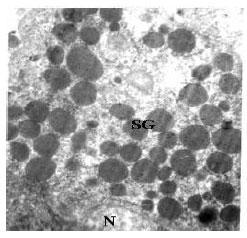 | |
| Fig. 6: | Transmission electron micrograph of intrahepatic exocrine pancreatic cells showing the presence of secretory granules (SG) in the apical cytoplasm and nucleus (N) in the basal part of the cytoplasm. X11500 |
Abundance of rough endoplasmic reticulum with parallel stacks of cisternae are located around the nucleus and along the plasma membrane. The cisternae of the rough endoplasmic reticulum are found to be filled with homogenous deposits of low density material which are uniform attachment of ribosomes with no free polysomes. The mitochondria varying in shape from circular to elongate are observed to be in close association with rough endoplasmic reticulum. These ultrastructural observations are compatible with the studies made earlier (Kendall and Hawkins, 1975; Hinton and Pool, 1976; Hugla and Thome, 1999; Yang and Chen; 2003; Vicentini et al., 2005; Ribeiro et al., 2006). The granular endoplasmic reticulum and mitochondria are equipped with oxidizing, reducing and conjugating abilities, thereby making the fish liver centre of detoxification activities. Chapman (1981) reported the absence of golgi apparatus and agranular endoplasmic reticulum in the hepatocytes of rainbow trout, Salmo gairdneri. In the present study also, the said organelles are noted to be absent from the hepatocytes of Schizothorax curvifrons. The absence or restricted amount of agranular endoplasmic reticulum and golgi apparatus in the normal teleost liver facilitates comparison between control and treated liver which may be encountered with the induction of agranular endoplasmic reticulum and golgi apparatus.
The presence of glycogen in the cytoplasm of hepatocytes is a characteristic feature of various fishes (Hampton et al., 1985; Moon et al., 1985; Gonzalez et al., 1993). The high content of glycogen in the liver of Schizothorax curvifrons confirm previous observations in hepatocytes of Salmo gairdneri (Scarpelli et al., 1963), Carassius auratus (Yamamoto, 1965), Micropterus salmoides (Hinton et al., 1972), Ictalurus punctatus (Hinton and Pool, 1976), Oreochromis niloticus (Vicentini et al., 2005). The presence of glycogen is suggestive of an ability of Schizothorax curvifrons for synthesis or breakdown of glycogen in relation to metabolic needs. Many investigators have reported the occurrence of intrahepatic exocrine pancreatic cells in teleosts and have suggested a secretory role to these cells (Kendall and Hawkins, 1975; Hinton and Pool, 1976; Stipp et al., 1980; Eurell and Haensly, 1982; Beccaria et al., 1992; Vicentini et al., 2005). Such cells have also been observed in the present study. The intrahepatic exocrine pancreatic cells are clearly distinguishable from the hepatocytes by the presence of large number of secretory granules of uniform density delimited by a single membrane. Dicentrarchus labrax (Beccaria et al., 1992), subjected to long fasting showed reduced cellular activity demonstrated by the scarcity of secretory granules. On the other hand, intensively fed fish were seen to have increased cellular activity and abundance of secretory granules. The electrodensed secretory granules are also abundant in Schizothorax curvifrons which are usually located in the apical portion of the cell, thereby displacing the nucleus towards the basal portion of the cell.
CONCLUSION
The fish liver appears well suited to serve as a model for analysis of the interaction between natural environmental changes and hepatic morphology at the subcellular level. Many environmental stressors may act on the liver including metabolic perturbations and lesions, these hepatic alterations may reflect a variety of reactions of the whole organism to intoxication, leading to disease or death. Such responses not only allow the fish liver to be considered as a good indicator of fish health status but also enforce the fact that it may be the future indicator of aquatic hazards. But it should be very clear that this all demands extensive studies at the subcellular level of the liver of normal and treated species of fishes.
ACKNOWLEDGMENTS
Authors wish to thank whole of the technical staff of the electron microscopy center of All India Institute of Medical Sciences (AIIMS) New Delhi, India, for their excellent technical assistance. They also thank Dr. T.C. Nag (Associate Prof. Deptt. of Anatomy AIIMS) not only for his proper and humble guidance but also for his helpful suggestions while carrying out this study.
REFERENCES
- Arstila, A.U., P. Hirsimaki and B.F. Trump, 1974. Studies on the subcellular pathophysiology of sublethal chronic cell injury. Beitr. Path., 152: 211-242.
PubMedDirect Link - Beccaria, C., J.P. Diaz and R. Connes, 1992. Effects of dietary conditions on the exocrine pancreas of the sea bass, Dicentrarchus labrax L. (Teleostei). Aquaculture, 101: 163-176.
CrossRef - Chapman, G.B., 1981. Ultrastructure of the liver of the fingerling rainbow trout Salmo gairdneri Richardson. J. Fish Biol., 18: 553-567.
CrossRefDirect Link - Eurell, J.A. and W.E. Haensly, 1982. The histology and ultrastructure of the liver of Atlantic croaker, Micropogon undulatus L. J. Fish Biol., 21: 113-125.
CrossRefDirect Link - Gonzalez, G., S. Crespo and J. Brusle, 1993. Histo-cytological study of the liver of the cabrilla sea bass, Serranus cabrilla (Teleostei, serranidae) an available model for marine fish experimental studies. J. Fish Biol., 43: 363-373.
CrossRef - Haensly, W.E., J.M. Neff, J.R. Sharp, A.C. Morris, M.F. Begood and P.D. Boem, 1982. Histopathology of Pleuronectes platessa L. from Aber Wrach and Aber Benoit, Brittany, France: Long term effects of the Amoco Cadiz crude oil spill. J. Fish Dis., 5: 365-391.
CrossRefDirect Link - Hampton, J.A., P.A. McCuskey, R.S. McCuskey and D.E. Hinton, 1985. Functional units in rainbow trout (Salmo gairdneri) liver: I. Arrangement and histochemical properties of hepatocytes. Anat. Rec., 213: 166-175.
PubMedDirect Link - Hinton, D.E. and C.R. Pool, 1976. Ultrastructure of the liver in channel catfish, Ictalurus punctatus (Rafinesque). J. Fish Biol., 8: 209-219.
CrossRef - Hinton, D.E., R.L. Snipes and M.W. Kendall, 1972. Morphology and enzyme histochemistry in the liver of largemouth bass (Micropterus salmoides). J. Fish. Res. Board Can., 29: 531-534.
Direct Link - Mir, I.H. and A. Channa, 2009. Gills of the snow trout, Schizothorax curvifrons Heckel: A SEM study. Pak. J. Biol. Sci., 12: 1511-1515.
CrossRefPubMedDirect Link - Mir, I.H. and A. Channa, 2010. A scanning electron microscopic examination of the intestinal tract of the snow trout, Schizothorax curvifrons Heckel. J. Fish. Aquat. Sci., 5: 386-393.
CrossRefDirect Link - Stipp, A.C., S. Ferri and A. Sesso, 1980. Fine structural analysis of a teleost exocrine pancreas cellular components. A freeze fracture and transmission electron microscopic study. Anat. Anz., 147: 60-75.
PubMed - Moon, T.W., P.J. Walsh and T.P. Mommsen, 1985. Fish hepatocytes: A model metabolic system. Can. J. Fish Aquat. Sci., 42: 1772-1782.
Direct Link - Pierce, K.V., B.B. McCain and S.R. Wellings, 1978. Pathology of hepatomas and other liver abnormalities in English sole (Parophrys vetulus) from the Duwamish River estuary, Seattle, Washington. J. Natl. Cancer Inst., 60: 1445-1453.
PubMedDirect Link - Pierce, K.V., B.B. McCain and S.R. Wellings, 1980. Histopathology of abnormal livers and other organs of starry flounder Platichthys stellatus (Pallas) from the estuary of the Duwamish River, Seattle, Washington, USA. J. Fish Dis., 3: 81-91.
CrossRef - Ribeiro, V.M.A., N. Bazzoli, T.A. Maria and G.B. Santos, 2006. Ultrastructural changes in female hepatocytes during ovarian maturation of Steindachnerina insculpta (Pisces: Curimatidae). Braz. J. Biol., 66: 957-962.
PubMed - Scarpelli, D.G., M.H. Greider and W.J. Frajola, 1963. Observations on hepatic cell hyperplasia, adenoma and hepatoma of rainbow trout (Salmo gairdneri). Cancer Res., 23: 848-857.
Direct Link - Segner, H. and H. Moller, 1984. Electron microscopical investigations on starvation-induced liver pathology in flounders Platichthys flesus. Mar. Ecol. Prog. Ser., 19: 193-196.
Direct Link - Studnicka, M., 1983. Histological and ultrastructural studies of muscles, liver and kidneys of brown bullhead (Ictalurus nebulosus) after experimental contamination with mercury. Acta Ichthyologica Piscatoria, 13: 99-122.
Direct Link - Vicentini, C.A., I.B. Franceschini-Vicentini, M.T.S. Bombonato, B. Bertolucci, S.G. Lima and A.S. Santos, 2005. Morphological study of the liver in the teleost Oreochromis niloticus. Int. J. Morphol., 23: 211-216.
Direct Link - Weis, P., 1974. Ultrastructural changes induced by low concentrations of DDT in the livers of the zebrafish and the guppy. Chem. Biol. Interaction, 8: 25-30.
CrossRef - Yang, J.L. and H.C. Chen, 2003. Serum metabolic enzyme activities and hepatocyte ultrastructure of common carp after gallium exposure. Zool. Stud., 42: 455-461.
Direct Link - Yamamoto, T., 1965. Some observations on the fine structure of the intrahepatic biliary passages in gold fish (Carassius auratus). Z. Zellforsch. Mikrosk. Anat., 65: 319-330.
PubMedDirect Link