
H. K. Hussein
Department of Biological Sciences, Faculty of Science, King Abdul-Aziz University, Jeddah, Saudi Arabia
O. A. Abu-Zinadah
Department of Biological Sciences, Faculty of Science, King Abdul-Aziz University, Jeddah, Saudi Arabia
International Journal of Zoological Research
Year: 2010 | Volume: 6 | Issue: 4 | Page No.: 266-276







ABSTRACT
This study aimed to determine the protective effect of curcumin on streptozotocin (STZ)-induced oxidative stress in various tissues of albino Wister rats. Adult male rats (8 weeks), weighing 195 to 225 g was made diabetic by injecting STZ (65 mg kg-1 body weight) intraperitoneally. During the whole experimental period, animals were fed with a balanced commercial chow and water ad libitum. Diabetic rats given either water or ethanolic curcumin extracts (80 mg kg-1 body weight) in aqueous suspension daily for a period of seven weeks. The levels of oxidative stress parameters and activity of antioxidant enzymes were determined in various tissues. STZ-induced hyperglycemia resulted in increased glucose level, glycosylated haemoglobin in red blood cells and other tissues and altered antioxidant enzyme activities such as AST and ALT. These elevated blood parameters and enzymatic activities induced by hyperglycemia were significantly restored to near normal by oral administration of curcumin once daily for 7 weeks, as compared to untreated rats. There was a significant elevation in the level of liver and kidney malondialdhyde (MDA), while the activities of antioxidant enzymes superoxide dismutase and catalase (SOD and CAT) were significantly decreased in STZ rats which also restored to normal after curcumin treatment. The results obtained indicated that ethanolic extract has more potent protective action than water extract against all hyperglycemic parameters. Biochemical observations were supplemented by histopathological examination of liver and kidney sections. Interestingly, feeding curcumin to the diabetic rats controlled oxidative stress by inhibiting the increase in TBARS and protein carbonyls and reversing altered antioxidant enzyme activities without altering the hyperglycemic state in most of the tissues. So, curcumin appear to be beneficial in preventing diabetes-induced oxidative stress in rats despite unaltered hyperglycemic status.
PDF Abstract XML References Citation
Received: March 31, 2010;
Accepted: May 06, 2010;
Published: June 10, 2010
How to cite this article
H. K. Hussein and O. A. Abu-Zinadah, 2010. Antioxidant Effect of Curcumin Extracts in Induced Diabetic Wister Rats. International Journal of Zoological Research, 6: 266-276.
DOI: 10.3923/ijzr.2010.266.276
URL: https://scialert.net/abstract/?doi=ijzr.2010.266.276
DOI: 10.3923/ijzr.2010.266.276
URL: https://scialert.net/abstract/?doi=ijzr.2010.266.276
INTRODUCTION
Hyperglycemia causes the autoxidation of glucose, glycation of proteins and the activation of polyol metabolism. These changes accelerate generation of Reactive Oxygen Species (ROS) and increases in oxidative chemical modification of lipids, DNA and proteins in various tissues. Oxidative stress may play an important role in the development of complications in diabetes such as lens cataracts, nephropathy and neuropathy. Glycation reactions, especially Millard reactions, occur in vivo as well as in vitro and are associated with the chronic complications of diabetes mellitus and aging and age-related diseases by increases in oxidative chemical modification of lipids, DNA and proteins (Meghana et al., 2007). Diabetes mellitus (type 2) is associated with increased oxidative stress (Mc-Coll et al., 1997). Free radicals are continually produced in the body as the result of normal metabolic processes and interaction with environmental stimuli. Under physiological conditions, a wide range of antioxidant defenses protects against adverse effects of free radical production in vivo (Halliwell and Gutteridge, 1989). Oxidative stress results from an imbalance between radical production and reduced activity of antioxidant defenses or both these phenomena. Hyperglycemia causes release of tissue damaging Reactive Oxygen Species (ROS) balance between radical production and protective antioxidant defense (Signorini et al., 2002; Halliwell and Gudtteridge, 1990). It has been proposed that streptozotocin (STZ) acts as a diabetogenic agent owing to its ability to destroy pancreatic β-cells, possibly by a free radical mechanism (Halliwell and Gutteridge, 1994).
The level of lipid peroxidation in cell is controlled by various cellular defense mechanisms consisting of enzymatic and nonenzymatic scavenger systems, the levels of which are altered in diabetes (Wohaieb and Godin, 1987). Moreover, disturbances of antioxidant defense systems in diabetes were shown: alteration in antioxidant enzyme (Strain, 1991), impaired glutathione metabolism (McLennan et al., 1991; Sajithlal et al., 1998) and decreased ascorbic acid (Jennings et al., 1987).
In recent years, considerable focus has been given to an intensive search for novel type of antioxidants from numerous plant materials (Srivastava et al., 1993). Management of diabetes without any side effects is still a challenge to the medical system. There is an increasing demand by patients to use the natural product switch antidiabetic activity, because insulin and oral hypoglycemic drugs possess undesirable side effects (Rao and Appa, 2001). Plants with antidiabetic activities provide useful sources for the development of drugs in the treatment of diabetes mellitus. Phytochemicals isolated from plant source are used for the prevention and treatment of cancer, heart disease, diabetes and high blood pressure etc. (Waltner-Law, 2002).
Curcumin (diferuloylmethane) is a naturally occurring yellow pigment isolated from the rhizomes of the plant Curcuma longa (Linn) found in south Asia (Lodha and Baggha, 2000) and is a potent antioxidant agent and free radical scavenger (Fujisawa et al., 2004). Along with being an inhibitor of lipid peroxidation (Sreejayan- Rao, 1994), it is also an inhibitor of Nitric Oxide Synthase (NOS) overexpression (Spinas, 1999; Pan et al., 2000) and of nuclear factor kappa B activation (Weber et al., 2006). The efficacy of curcumin has been widely observed in reducing various diabetic secondary complications such as diabetic nephropathy/renal lesions (Sharma et al., 2006), retinopathy (Kowluru and Kanwar, 2007), wound healing (Panchatcharam et al., 2006) and reduction of advanced glycation end products (Sajithlal et al., 1998). Its potential as a hypoglycemic agent has also been studied in animals (Pari and Murugan, 2005; Hussain, 2002) and humans (Meghana et al., 2007) with conflicting results. Furthermore, although many aspects of curcumin-induced cytoprotection are studied, its efficacy in protecting against streptozotocin-induced free radical-mediated damage has not been demonstrated so far. We hypothesized that curcumin may protect cells against streptozotocin-induced oxidative stress and resulting tissue damage and organ dysfunction. Logically, curcumin would impart some protection against oxidative damage to different cells, but it would be of interest to investigate the mode of curcumin-induced cytoprotection. In this study, the protective action of curcumin on streptozotocin-induced diabetes mellitus in rats was studied.
MATERIALS AND METHODS
Animals
Eight week male albino Wister rats weighing (195- 225 g) were used in this study (Vide No. 291, 2009). The animals were obtained from the animal experimental unit of King Fahd Medical Research Center, King Abdul Aziz University, Jeddah, Saudi Arabia. The rats were housed in well-aerated individual cages and maintained in a temperature-controlled room (27±2°C) and humidity 45-56% with12 h light: 12 h dark cycle for one week before and during the experiments. Animals were provided with standard commercial chow and water ad libitum. The care and use of all experimental animals complied with relevant animal welfare laws. Streptozotocin, all reagents and solvents used in this study were purchased from Sigma-Aldrich Corp., St Luis. MO, USA but Cinnamon powder was obtained from the local market at Jeddah, Saudi Arabia. Type 2 diabetes mellitus was induced in overnight fasted rats by a single intraperitonial injection (i.p.) of 65 mg kg-1 body weight Streptozotocin STZ (Masiello et al., 1998). STZ was dissolved in citrate buffer (pH 4.5). Hyperglycemia was confirmed by the elevated glucose levels in plasma, determined at 72 h and then on day 7 after injection. The animals with blood glucose concentration more than 200 mg dL-1 will be used for the study.
Preparation of Curcumin Extract
The dried powder was defatted with petroleum ether (100 g in 200 mL ether). The defatted material was extracted with 95% ethanol and then vacuum dried. One part of powder was extracted in boiling water and then filtered and vacuum dried.
Experimental Design
In the experiment, a total of 40 rats (30 diabetic surviving rats, 10 normal rats) were used. The rats were divided into four groups of 10 each. The experimental period was seven weeks beginning after the induction of STZ diabetes. Group I: normal untreated rats. Group II: diabetic control rats (STZ group). Group III and IV: were diabetic rats given either aqueous or ethanolic extracts of curcumin (80 mg kg-1 body weight) in aqueous suspension daily using an intragastric tube (Arun and Nalini, 2002) for seven weeks. At the end of experimental period, the rats were deprived of food overnight and blood was collected in a tube containing potassium oxalate and sodium fluoride for the estimation of blood glucose, haemoglobin and glycosylated haemoglobin. Plasma was separated for the assay of insulin. The liver and kidney were also dissected out and were divided into two parts. The first was kept at -20°C in ice-cold containers for biochemical analysis, while the second part was used for histopathological studies.
Serum Biochemical Assay
Serum enzymes aspartate aminotransferase (AST) and serum glutamate pyruvate transaminase (ALT) were determined according to Reitman and Frankel (1957).
Estimation of MDA, SOD, CAT in Liver and Kidney Tissues
Liver and kidney samples were dissected out and washed immediately with ice cold saline to remove as much blood as possible. Each tested tissue homogenates (5% w/v) were prepared in cold 50 mM potassium phosphate buffer (pH 7.4) using glass homogenizer in ice. The cell debris was removed by centrifugation at 5000 rpm for 15 min at 40°C using refrigerated centrifuge. The supernatant was used for the estimation of malondialdehyde (MDA) (Ohkawa et al., 1979), superoxide dismutase (SOD) (Kakkar et al., 1972) and catalase (CAT) (Sinha, 1972) levels.
Histopathological Studies
The liver and kidney tissues was dissected out and fixed in 10% formalin, dehydrated in gradual ethanol (50-99%), cleared in xylene and embedded in paraffin. Sections were prepared and then stained with hematoxylin and eosin dye for microscopic investigation.
Statistical Analysis
Statistical analysis was performed on a PC using SPSS, V.13, (special package for social sciences). Data are presented as arithmetic mean±SD. The difference among means has been analyzed by one way ANOVA followed by student t-test. A value of p<0.05 was considered as statistically significant.
RESULTS AND
DISCUSSION
The results of protective effect of water or ethanol extracts of curcumin on streptozotocin-induced diabetic rats (STZ) are shown in Table 1. In the STZ group, serum AST and ALT were significantly increased as compared to control group (p<0.001). The elevated activities of serum AST and ALT were significantly reduced in the animal groups treated with either water or ethanolic extracts. Treatment with ethanolic extract showed significantly more activity (p<0.001) than with water extract. Thus, the ethanol extract treated group was superior to the water extract. Results obtained revealed an increase in the level of liver and kidney MDA in STZ rats compared to control group. Treatment with extracts significantly prevented this raise in levels. The activities of SOD and CAT were significantly reduced in the STZ group, while they were significantly elevated near the normal values in the groups pretreated with either extracts. Ethanolic extract has been shown to be more protective than water extract.
| Table 1: | Antioxidant enzyme activities of liver and kidney of rat; superoxide dismutase (SOD), catalase (CAT), lipid peroxide 7 product or Malendialdlyde (MDA) and serum aminotransferase enzymes (ALT and AST) of all studied groups (Mean±SD) |
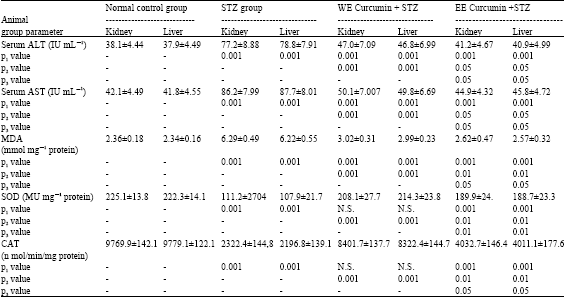 | |
| STZ: Streptozotocin, WE: Water extract, EE: Ethanol extract, P1: Comparison to normal control, P2: Comparison to STZ group, P3: Water E versus ethanol E, N.S= Not significant | |
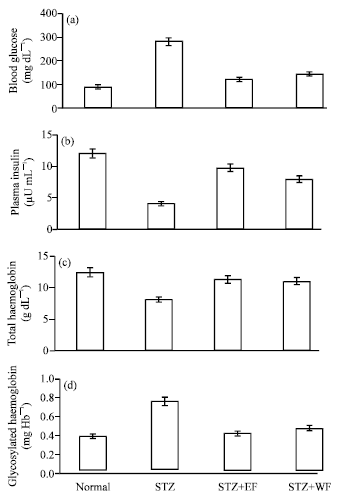 | |
| Fig. 1: | Effect of curcumin on (a) The levels of blood glucose, (b) Plasma insulin, (c) Total haemoglobin and (d): Glycosylated haemoglobin in normal and experimental rats: STZ, Diabetic control; STZ+EE, Diabetic treated with ethanolic curcumin extract; STZ+WE, Diabetic treated with water curcumin extract. Values are given as means for 10 rats in each group±SD |
The levels of blood glucose, total haemoglobin, glycosylated haemoglobin and plasma insulin of different experimental groups were shown in Fig. 1a-d. There was a significant elevation in blood glucose level, whereas plasma insulin levels decreased significantly in diabetic rats (STZ groups), compared with normal rats. Administration of water or ethanolic curcumin extracts tended to bring blood glucose and plasma insulin towards normal. The diabetic control rats showed a significant decrease in the level of total haemoglobin and significant increase in the level of glycosylated haemoglobin. No significant difference between liver and kidney parameters were found in all studied groups. Oral administration of water and ethanolic curcumin extracts to diabetic rats significantly restored total haemoglobin and glycosylated haemoglobin levels. The effect of ethanolic extract was more prominent when compared with water extract.
Figure 2 shows the histological examination of liver sections of control animals (Fig. 2A) showed normal hepatic cells with well preserved cytoplasm prominent nucleus. The livers of STZ rats showed massive fatty changes, necrosis and broad infiltration of the lymphocytes (Fig. 2B). The histological architecture of liver sections of the rats treated with either aqueous (Fig. 2C) or ethanolic extracts (Fig. 2D) showed more or less normal patterns, with a mild degree of fatty change, necrosis and lymphocyte infiltration, almost comparable to those of the control group.
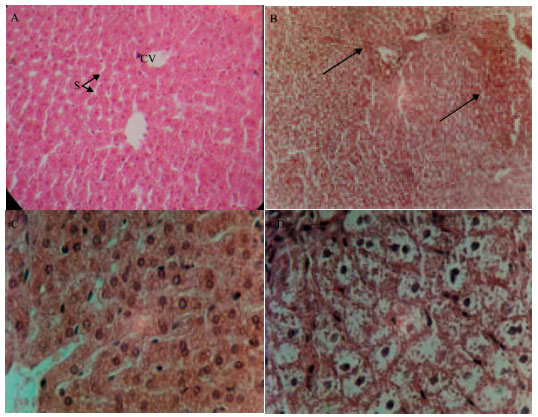 | |
| Fig. 2: | (A) Normal hepatic tissues showing hepatic strands of cells around the central vein (CV) leaving blood sinusoids (S) X 440, (B) hepatic tissues of STZ group showing cellular necrosis around the central veins (arrow) X 250, (C) hepatic tissues pretreated with water extract X 440 and (D) pretreated with ethanol extract showing absence of necrosis X 750 (H and E stains) |
Pathological changes of kidney (Fig. 3) showing that the normal renal tissues illustrated in Fig. 3A which showing normal uriniferous tubules and glomeruli were changed in diabetic control rats (STZ group). Diabetic control rat's kidney showed tubular epithelial damage, capillary proliferation certain degenerated uriniferous tubules and dilatation of Bowman's capsule (Fig. 3B). The above pathological changes were reduced in rats treated with water and ethanolic curcumin extracts (Fig. 3C and D, respectively).
Although many reports confirm the efficacy of curcumin in cytoprotection of various cell lines against toxicity (Grandjean-Laquerriere et al., 2002; Yadav et al., 2005; Masamune et al., 2006) few studies report the efficacy of curcumin in mitigating streptozotocin-induced damage to liver and kidney. Most diabetes related studies using curcumin show the efficacy of oral administration at various doses for reduction of secondary complications in streptozotocin-induced diabetic animals (Babu and Srinivasan, 1995; Nishizono et al., 2000; Srinivasan et al., 2003). However, almost no reports exist that show the prophylactic importance of curcumin as a diabetic protective agent. Present findings here come in agreement with the above cited findings and contribute towards retention of type 2 diabetes which is the most prevalent and serious metabolic disease affecting people all over the world and cause secondary complications by scavenging free radicals that cause pathological changes and dysfunction of various organs.
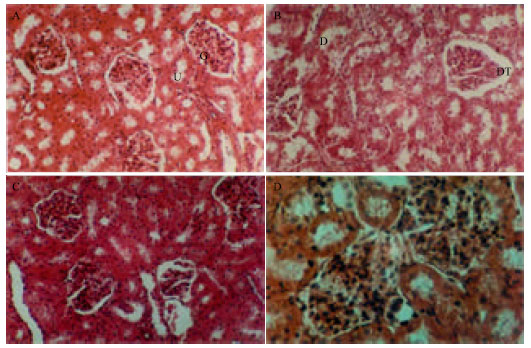 | |
| Fig. 3: | (A) Normal renal tissues showing normal urineferous tubules (u) and glomeruli (G) X40, (B) renal tissues of STZ group showing certain degeneratedated urineferous tubules (D) and dilatation of Bowman's capsule (DT), X 440, (C): renal tissues pretreated with water extract X 440 and (D) Pretreated with ethanol extract showing normal renal structure X 750 (H and E stains) |
Present results support the finding of Koenig et al. (1976), who stated that the glycosylated haemoglobin was significantly increased in diabetic control rats and this increase is directly proportional to fasting blood glucose. Anemia is much more common disease in type 2 diabetic patients, contributing to the pathogenesis of diabetic complications. In the present study, the decreased concentration of haemoglobin indicates the anemia in STZ diabetic rats, in as much as during diabetes, the excess glucose transport in the blood reacts with haemoglobin to form glycosylated haemoglobin. Free radicals react with lipids and causes peroxidative changes that result in enhanced lipid peroxidation (Simmons, 1984; Girotti, 1985). In this study, the lipid peroxidation markers (TBARS) were elevated in erythrocytes of diabetic rats which support the results reported earlier by Zhang and Swaan (1999). The increase in lipid peroxidation might be a reflection of decrease in enzymatic and nonenzymatic antioxidants of defense systems.
Serum AST and ALT activities were used as a marker of tissue damage. Diabetes mellitus by STZ produces an experimental damage due to its toxic metabolite (Zhang and Swaan, 1999). The toxic metabolite free radical is produced by cytochrome p450 which further reacts with oxygen to produce trichloromethyl peroxy radicals. These radicals bind covalently with the macromolecule and cause peroxidative degradation of lipid membranes of the liver and kidney. The reduction of AST and ALT activities by the extracts is an indication of repair of tissue damage induced by diabetes complications. This is in agreement with Shahidi and Wanasundara (1992), who found that serum transaminases returned to normal activities with the healing of tissue parenchyma and regeneration of hepatocytes and renal tissues. The ethanolic extract induced suppression of increased ALT and AST activities. Thus, administration of ethanolic or aqueous extracts of curcumin revealed protective activity against the toxic metabolites of diabetes, which is also supported by histological studies.
Treatment with curcumin brought back lipid peroxidation markers to near normal levels, which could be as a result of improved glycemic control and antioxidants status. We have reported that curcumin has significant glucose reducing property in STZ diabetic rats. Increased lipid peroxidation under diabetic conditions can be due to increased oxidative stress in the cell as a result of depletion of antioxidant scavenger systems. Associated with the changes in lipid peroxidation the diabetic tissues showed decreased activities of key antioxidants SOD and CAT and increase MDA which play an important role in scavenging the toxic intermediate of incomplete oxidation. SOD and CAT are the two major scavenging enzymes that remove toxic free radicals in vivo. Previous studies have reported that the activity of SOD is low in diabetes mellitus (Feillet-Coudray et al., 1999) come in agreement with our results. A decrease in the activity of these antioxidants can lead to an excess availability of superoxide anion O2←- (free radical anion) and hydrogen peroxide in biological systems, which intern generate hydroxyl radicals, resulting in initiation and propagation of lipid peroxidation (Kumthekar and Katyare, 1992). The result of increased activities of SOD and CAT suggest that curcumin contains a free radical scavenging activity, which could exert a beneficial effect against pathological alterations caused by the presence of O2← and OH←. The increased activity of SOD accelerates dismutaion of O2←- to hydrogen peroxide (H2O2) which is removed by CAT (Aebi, 1984). This action could involve mechanisms related to scavenging activity of curcumin. Lipid peroxidation is accelerated when free radicals are formed as the results of losing a hydrogen atom from the double bond in the structure of unsaturated fatty acids. The free radical scavenging activity of water or ethanolic extracts of curcumin was evaluated.
Scavenging of free radicals is one of the major antioxidation mechanisms to inhibit the chain reaction of lipid peroxidation. Reduced lipid peroxidation was revealed by a significant decrease in MDA level in groups pretreated with water or ethanol extracts, simultaneously with a significant elevation in SOD and CAT activities. The present study revealed that SOD and CAT activities decreased in STZ animals, which may be due to altered antioxidant status. This is in accordance with results that indicated a decreased CAT in STZ animals may be due to the utilization of antioxidant enzymes in the removal of released H2O2 released (Cerutti et al., 1994). SOD and CAT activities increased significantly in the treated group versus the untreated animals.
In present study, histopathological observation in diabetic control rats causes conges mild inflammation; sinusoidal congestion with fatty degeneration in the form of fat lake in the liver and tubular epithelial damage messangial capillary proliferation; fatty infiltration in the kidney. The reaction is provoked by the increased production of highly reactive intermediates of STZ, which are normally detoxified by endogenous Growth Stimulating Hormone (GSH) but when present in excess, can deplete GSH stores, allowing the reactive intermediate to react with and destroy hepatic, renal cells (Blum and Fridovich, 1985). The above pathological changes were reduced in diabetic rats treated curcumin. The histological evidence of diabetic control rats suggest that structural alterations at the end of experiment are due to STZ induced free radical generation quite early in diabetes. Thus in addition to blood glucose lowering effect, histopathological observations also supports the notion that curcumin produced significant antihyperglycemic activity by protecting the tissues against STZ action. The results obtained revealed that ethanolic extract of curcumin has more potent antioxidant activity than water extract. The antioxidant properties of curcumin extracts are attributable to the ability of its phenolic constituents to quench reactive oxygen species. The protective effect is documented by the biochemical and histopathological data obtained. This data supports the uses of curcumin extract in treatment of some hepatic and renal disorders.
In conclusion, the present investigation shows that curcumin possesses antioxidant effect that may contribute to its protective action against lipid peroxidation and enhancing effect on cellular antioxidant defense. This activity contributes to the protection against oxidative damage in STZ induced diabetes. Further study will be carried out to identify the types of phenolic compounds present and to test for antitumor activity.
REFERENCES
- Arun, N. and N. Nalini, 2002. Efficacy of turmeric on blood sugar and polyol pathway in diabetic albino rats. Plant Foods Hum. Nutr., 57: 41-52.
CrossRefPubMedDirect Link - Babu, P.S. and K. Srinivasan, 1995. Influence of dietary curcumin and cholesterol on the progression of experimentally induced diabetes in albino rat. Mol. Cell Biochem., 152: 13-21.
PubMed - Blum, J. and I. Fridovich, 1985. Inactivation of glutathione peroxidase by superoxide radical. Arch. Biochem. Biophys., 240: 500-508.
CrossRefPubMedDirect Link - Cerutti, P., R. Ghosh, Y. Oya and P. Amstad, 1994. The role of cellular antioxidant defence in oxidant carcinogenesis. Environ. Health Perspect, 102: 123-130.
Direct Link - Feillet-Coudray, C., E. Rock, C. Coudray, K. Grzelkowska, V. Azais-Braesco, D. Dardevet and A. Mazur, 1999. Lipid peroxidation and antioxidant status in experimental diabetes. Clin. Chim. Acta, 284: 31-34.
PubMed - Fujisawa, S., T. Atsumi, M. Ishihara and Y. Kadoma, 2004. Cytotoxicity, reactive oxygen species-generating activity and radical scavenging activity of curcumin and other related compounds. Anti-Cancer Res., 24: 563-570.
Direct Link - Halliwell, B. and J.M.C. Gutteridge, 1990. Role of free radicals and catalytic metal ions in human disease: An overview. Methods Enzymol., 186: 1-85.
CrossRefDirect Link - Halliwell, B. and J.M. Gutteridge, 1994. Lipid peroxidation, oxygen radicals, cell damage and antioxidant theraphy. Lancet, 1: 1396-1397.
PubMed - Hussain, H.E.M.A., 2002. Hypoglycemic, hypolipidemic and antioxidant properties of combination of Curcumin from Curcuma longa, Linn and partially purified product from Abroma augusta, Linn. in streptozotocin induced diabetes. Indian J. Clin. Biochem., 17: 33-43.
CrossRefDirect Link - Jennings, P.B., S. Chirico, A.F. Jones, J. Lunec and A.H. Barnett, 1987. Vitamin C metabolites and microangiopathy in diabetes mellitus. Diabetes Res., 6: 151-154.
Direct Link - Kakkar, P., B. Das and P.N. Viswanathan, 1984. A modified spectrophotometric assay of superoxide dismutase. Indian J. Biochem. Biophys., 21: 130-132.
PubMedDirect Link - Rao, B.K. and C.H. Rao, 2001. Hypoglycemic and antihyperglycemic activity of Syzygium alternifolium (Wt.) Walp. seed extracts in normal and diabetic rats. Phytomedicine, 8: 88-93.
PubMedDirect Link - Koenig, R.J., C.M. Peterson, R.L. Jones, C. Saudek, M. Lehrman and A.C. Cerami, 1976. Correlation of glucose regulation and haemoglobin Aic in diabetes mellitus. N. Eng. J. Med., 295: 417-420.
Direct Link - Kowluru, R.A. and M. Kanwar, 2007. Effects of curcumin on retinal oxidative stress and inflammation in diabetes. Nutr. Metab., 4: 1-8.
PubMed - Kumthekar, M.M. and S.S. Katyare, 1992. Altered kinetic attributes of Na++K+-ATPase activity in kidney, brain and erythrocyte membranes in alloxan-diabetic rats. Indian J. Exp. Biol., 30: 26-32.
Direct Link - Lodha, R. and A. Baggha, 2000. Traditional Indian systems of medicine. Ann. Acad. Med. Singapore., 29: 37-41.
PubMed - Waltner-Law, M.E., L.X. Wang, B.K. Law, R. Hal, M. Nawano and D.K. Granner, 2002. Epigallocatechin gallate, a constituent of green tea, represses hepatic glucose production. J. Biol. Chem., 277: 34933-34940.
CrossRefDirect Link - Masiello, P., C. Broca, R. Gross, M. Roye and M. Manteghetti et al., 1998. Experimental NIDDM: Development of a new model in adult rats administered streptozotocin and nicotinamide. Diabetes, 47: 224-229.
CrossRefDirect Link - McLennan, S.V., S. Heffernan, L. Wright, C. Rae, E. Fisher, D.K. Yue and J.R. Turtle, 1991. Changes in hepatic glutathione metabolism in diabetes. Diabetes, 40: 344-348.
CrossRefDirect Link - Meghana, K., G. Sanjeev and B. Ramesh, 2007. Curcumin prevents streptozotocin-induced islets damage by scavenging free radicals: A prophylactic and protective role. Eur. J. Phamacol., 577: 183-191.
Direct Link - Nishizono, S., T. Hayami, I. Ikeda and K. Imazumi, 2000. Protection against the diabetogenic effect of feeding tert butylhydroquinone to rats prior to the administration of streptozotocin. Biosci. Biotechnol. Biochem., 64: 1153-1158.
PubMed - Panchatcharam, M., S. Miriyala, V.S. Gayathri and L. Suguna, 2006. Curcumin improves wound healing by modulating collagen and decreasing reactive oxygen species. Mol. Cell. Biochem., 290: 87-96.
CrossRefDirect Link - Pari, L. and P. Murugan, 2005. Effect of tetrahydrocurcumin on blood glucose, plasma insulin and hepatic key enzymes in streptozotocin induced diabetic rats. J. Basic Clin. Physiol. Pharmacol., 16: 257-274.
Direct Link - Reitman, S. and S. Frankel, 1957. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol., 28: 56-63.
CrossRefPubMedDirect Link - Shahidi, F., P.K. Janitha and P.D. Wanasundara, 1992. Phenolic antioxidants. Crit. Rev. Food Sci. Nutr., 32: 67-103.
CrossRefDirect Link - Sajithlal, G.B., P. Chithra and G. Chandrakasan, 1998. Effect of curcumin on the advanced glycation and cross-linking of collagen in diabetic rats. Biochem. Pharmacol., 56: 1607-1614.
Direct Link - Sharma, S., S.K. Kulkarni and K. Chopra, 2006. Curcumin, the active principle of turmeric (Curcuma Longa), ameliorates diabetic nephropathy in rats. Clin. Exp. Pharmacol. Physiol., 33: 940-945.
CrossRefPubMedDirect Link - Signorini, A.M., C. Fondelli, E. Renzoni, C. Puccetti, G. Gragnoli and G. Giorgi, 2002. Antioxidant effect of gliclazide, glibenclamide and metformin in patients with type 2 diabetes mellitus. Curr. Ther. Res., 63: 411-420.
CrossRef - Simmons, K., 1984. Defense against free radicals has therapeutic implications. J. Am. Med. Assoc., 251: 2191-2192.
PubMed - Spinas, G.A., 1999. The dual role of nitric oxide in islet β-cells. News Physiol. Sci., 14: 49-54.
Direct Link - Sreejayan, N. and M.N.A. Rao, 1994. Curcuminoids as potent inhibitors of lipid peroxidation. J. Pharm. Pharmacol., 46: 1013-1016.
CrossRefDirect Link - Srivastava, Y., H. Venkatakrishna-Bhatt, Y. Verma, K. Venkaiah and B.H. Raval, 1993. Antidiabetic and adaptogenic properties of Momordica charantia extract: An experimental and clinical evaluation. Phytother. Res., 7: 285-289.
Direct Link - Strain, J.J., 1991. Disturbances of micronutrient and antioxidant status in diabetes. Proc. Nutr. Soc., 50: 591-604.
Direct Link - Wohaieb, S.A. and D.V. Godin, 1987. Alterations in free radical tissue defense mechanism in STZ induced diabetes in rat, effects of insulin treatment. Diabetes, 36: 1014-1018.
PubMed - Weber, W.M., L.A. Hunsaker, C.N. Roybal, E.V. Bobrovnikova-Marjon and S.F. Abcouwer et al., 2006. Activation of NF kappa B is inhibited by curcumin and related enones. Bioorg. Med. Chem., 14: 2450-2461.
PubMed - Yadav, V.S., K.P. Mishra, D.P. Singh, S. Mehrotra and V.K. Singh, 2005. Immunomodulatory effects of curcumin. Immunopharmacol. Immunotoxicol., 27: 485-497.
PubMedDirect Link - Ohkawa, H., N. Ohishi and K. Yagi, 1979. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem., 95: 351-358.
CrossRefPubMedDirect Link - Zhang, E.Y. and P.W. Swaan, 1999. Determinition of membrane protein glycation in diabetic tissue. AAPS Pharm. Sci., 20: 1-7.
CrossRefDirect Link