
K. Granados- Flores
Instituto de Ciencias Del Mar y Limnologia, UNAM, Apartado Postal 70-305, Mexico D.F., 09340, Mexico
M. Gallardo- Cabello
Instituto de Ciencias Del Mar y Limnologia, UNAM, Apartado Postal 70-305, Mexico D.F., 09340, Mexico
E. Espino- Barr
CRIP-Manzanillo, INAPESCA, Playa Ventanas s/n, Manzanillo, Colima, 28200, Mexico
E.G. Cabral- Solis
CRIP-Manzanillo, INP, Playa Ventanas s/n, Manzanillo, Colima, 28200, Mexico
International Journal of Zoological Research
Year: 2010 | Volume: 6 | Issue: 1 | Page No.: 1-12







ABSTRACT
This study deals with age determination of the brassy grunt Microlepidotus brevipinnis by reading otoliths (sagittae) and scales which allowed the identification of 13 age groups. Growth of otoliths and scales is proportional to the growth of the fish. The time of formation of fast and slow growth bands in sagittae, as well as the time of ring formation in the scales is one year. The highest growth in length of this species takes place during the first year of life, in which the organism reaches 125.0 mm, this reduces natural mortality of the individuals by decreasing depredation. Growth in weight of this species is isometric. Sexual differentiation of the organisms is apparent after the age of two years and there were differences in the average length for each age in scales and otoliths for males and females. The age of other members of the Haemulidae family were compared with those obtained in the present study and M. brevipinnis reaches its oldest age in 13 years. It occupies a middle point (442.4 mm) in relation to the total lengths of other haemulids in the Mexican Pacific coast.
PDF Abstract XML References Citation
How to cite this article
K. Granados- Flores, M. Gallardo- Cabello, E. Espino- Barr and E.G. Cabral- Solis, 2010. Age Determination of Microlepidotus brevipinnis (Steindachner, 1869) (Pisces: Haemulidae) in the Coast of Jalisco, Mexico, by Reading Otoliths and Scales. International Journal of Zoological Research, 6: 1-12.
DOI: 10.3923/ijzr.2010.1.12
URL: https://scialert.net/abstract/?doi=ijzr.2010.1.12
DOI: 10.3923/ijzr.2010.1.12
URL: https://scialert.net/abstract/?doi=ijzr.2010.1.12
INTRODUCTION
Microlepidotus brevipinnis (Steindachner, 1869) (Pisces: Haemulidae) (Fig. 1) represents 10% of the catch of the artisanal fishery in the coast of Jalisco, Mexico (Espino-Barr et al., 2004a), 2 to 11 tons from 2000 to 2007. Locally it is very appreciated due to its low price and great flavor.
This and other species of the Haemulidae family are mentioned in the taxonomic lists of Castro-Aguirre (1978), López and Bussing (1982), Van Der Heiden (1985), Chávez-Ramos et al. (1994), Allen and Robertson (1994), Béarez (1996), Madrid-Vera et al. (1998), Espion-Barr et al. (2003), Mariscal-Romero and van der Heiden (2006). Age and growth analysis of five species of this family were performed by Cruz-Romero et al. (1993) using an indirect method of length-frequency. Gallardo-Cabello et al. (2003) and Espino-Barr et al. (2004a, b) also analyzed age growth of Anisotremus interruptus. Taking in account the scarce information available on these species, it is important to analyze the parameters of their population dynamics and determine the age by direct methods.
 | |
| Fig. 1: | Brassy grunt Microlepidotus brevipinnis |
The direct methods to determine the age based on the interpretation of growth rings in hard structures as: scales (Gallardo-Cabello et al., 2005), otoliths (Gallardo-Cabello et al., 2006; Espino-Barr et al., 2006), vertebrae (Anislado-Tolentino and Robinson-Mendoza, 2001) and spines (Alvarado-Castillo and Félix-Uraga, 1998), among others. These direct methods allow determining the age groups with a higher precision as compared to the indirect methods of length frequency analysis, eliminating most of the biases produced by small samples taken with low periodicity and allowing to identify the first age groups (Peñailillo and Araya, 1996; Campana, 1999; Campana and Thorrold, 2001; Mascareñas-Osorio et al., 2003; Berg et al., 2005; Francis et al., 2005).
The purpose of this study was
| • | Determine age groups of M. brevipinnis using otoliths and scales, to validate each other |
| • | Evaluate the time of formation of growth bands |
| • | Relationship between the growth of these structures and of the fish |
| • | Analyze differences between sexes |
| • | Compare lengths at age with other species of the Haemulidae family |
These results will provide information on some aspects of the life history of this species that will help assess the fishery and suggest regulation measures.
MATERIALS AND METHODS
During five days of every month from January 2005 to June 2006, samplings of 1050 individuals from the commercial catch were taken in Melaque, coast of Jalisco (104°40’30” to 104°43’30”W and 19°12’00” to 19°13’30”N). Of each individual total and standard length, height, total and eviscerated weight was measured. Sex was determined for 480 individuals; the other 570 were eviscerated by fishers before landing. For age study two structures were obtained from 353 individuals: otoliths (sagittae) and scales.
To obtain the otoliths sagittae, a transverse cut was made in the ventral cranial cavity and the brain was removed; the left and right semicircular canals were extracted and otoliths separated, rinsed with water, dried and stored dry in Eppendorf tubes labeled with number, date, total length and sex. Data on the length and the width were regarded for each otolith through their observation in a stereoscopic microscope with a graduated ocular lens (Fig. 2a, b). All measures were made on the right-sided sagitta, in order to eliminate possible differences due to variations between the otoliths in the same organism. The sample size was calculated with the formula described by Daniel (2002).
In the case of the sagittae, constants of the relationship were calculated for Rostrum Length (RL), Antirostrum Length (AL) and Width (Wi). The rostrum length was also related to the total length of the fish.
 | |
| Fig. 2: | Left and right sagittae of M. brevipinnis, (a) external and (b) internal aspect |
 | |
| Fig. 3: | Scale of M. brevipinnis |
Regressions were done by least squares and a variance analysis (anova) was carried out (Mendenhall, 1987; Zar, 1996). This last test was also applied to analyze possible morphometric differences between the otoliths of males and females. Identification of growth rings was done observing the sagittae in a stereoscopic microscope with transmitted light. The time of the growth ring formation was analyzed, observing whether the sagittae borders had fast or slow growth rings.
Around 15 scales were taken from the area under the left pectoral fin, below the lateral line (Ehrhardt, 1981; Holden and Raitt, 1975; Ruiz-Durá et al., 1970) and stored in dry labeled envelopes. Following the method described by Holden and Raitt (1975) the scales were washed to clean them of any tissue stuck to them. Later, 10 scales of each individual were placed between two slides, sealing them with adhesive tape and labeled. Reading of the scales was done with the help of a transparency projector Kodak Ektagraphic with a 127 mm lens (which increases the size of the scale 13.4 times). Scales were read independently by two different individuals and the results were compared (Fig. 3).
Determination of the marginal increment was carried out monthly for each individual, in order to determine the date in which the mark is formed and to validate its periodicity.
Analysis of otoliths and scales showed that M. brevipinnis recruited by the fishing gear has four growth rings, therefore the sampling strategy was modified and a biological sampling was included to capture younger specimens, with a ½ inch mesh net.
RESULTS
Morphometric Analysis of the Species
Table 1 shows the relationship between total length vs. standard length, height, total and eviscerated weight of M. brevipinnis. The relationship between total length and standard length and total length and height are expressed by the values of the exponents b = 1.000 and b = 1.042, respectively for the species which correspond to an isometric growth (Table 2). Some differences of these values are shown for sexes and the undetermined specimens.
The relationship between total length and total and eviscerated weight are described by the values of the exponents b = 3.042 and b = 2.954, respectively, which represents an isometric growth, which means that the fish grows similarly in length and weight (Table 3).
| Table 1: | Values of standard length, height, total and eviscerated weight for total length classes of M. brevipinnis |
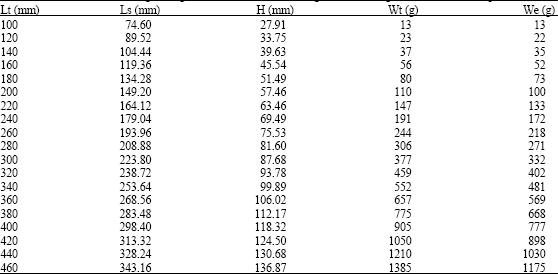 | |
| Table 2: | Values for total length vs. standard length and height for the species, females, males and undetermined of M. brevipinnis |
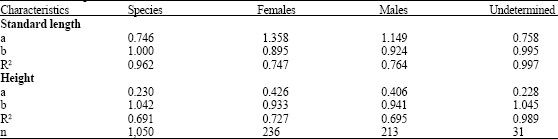 | |
| Table 3: | Values for total length vs. total and eviscerated weight for the species, females, males and undetermined individuals of M. brevipinnis |
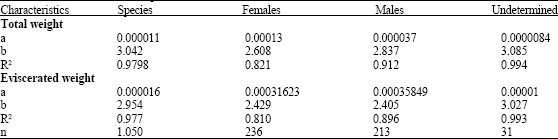 | |
In the case of males and females, values show a tendency to a negative allometry. The highest values of these indexes are reached by undetermined specimens with values b = 3.085 and b = 3.027 for total and eviscerated weight, respectively.
Otoliths Description of the Sagittae
The anterior margin exhibits a small excisura, as a tiny notch, showing an underdeveloped antirostrum (Fig. 2). The shape of the rostrum varies between individuals and also between the right and the left sagittae in the same specimen. The posterior margin presents a sharped postrostrum without an excisura minor and a pararostrum. The dorsal margin is rectilinear from the antirostrum to the center of the otolith, beyond which the dorsal margin descends with a marked inclination towards the posterior margin. The ventral margin is curved from the rostrum to the posterior margin. The anterior, posterior and ventral margins show regular denticles that spread irregularly and disappear in some sections of the otolith as the fish ages. The internal aspect of the otolith is convex; this feature increases with age; its surface is smooth and marked lengthwise by a deep sulcus which decreases in width from the center of the otolith to the end of its posterior margin, showing a clear difference between the ostium and the cauda. The external aspect is concave; its thickness decreases abruptly in the longitudinal sense from the middle area to the anterior margin. The average width of the sagitta is 2.1 times its length.
Growth of the Sagitta
Results of fish length classes and different measures of the otolith are shown in Table 4. The relationship between length and width of the sagitta is expressed by the value of exponent b = 0.64 which corresponds to a negative allometric growth (Table 5). The determination index of the relationship of the two series of data is R2 = 0.75, with F = 997 of the anova, which indicates that the sagitta grows more in length than widthwise.
| Table 4: | Rostrum (RL), width (Wi) and antirostrum (AL) length in different size classes of the sagitta of Microlepidotus brevipinnis |
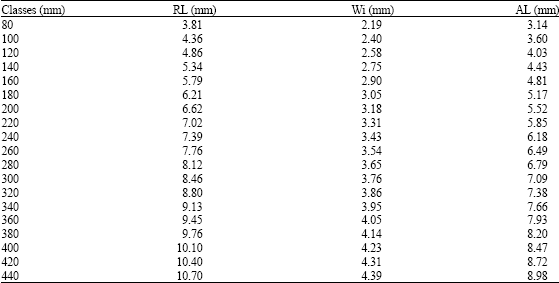 | |
| Table 5: | Relationships between the rostrum and other measures of the sagittae |
The allometric relationship between the length of the rostrum and of the antirostrum is expressed in the isometric growth index b = 1.02, with the highest values of R2 and F obtained for this analysis, i.e., 0.96 and 7751, respectively. This shows that among the sizes and age groups, the length of the antirostrum and rostrum increase in a similar way.
The relationship between the fish total length and the length and width of the sagitta is shown in the Table 6. The highest value of the allometric index related to the total length of the fish corresponds to the rostrum and antirostrum length, b = 0.60; this means that there is a proportionality between the increase of the length of the sagitta and of the fish. This structure is suitable to describe the growth in length of the organism. The values of the determination index R2 and anova (F) show a high correlation between the analyzed structures in every case. The values of the allometric growth indexes are lower than 1 due to the difficulty to relate very small structures with the corporal length of the fish.
Identification of the Growth Rings
Analysis of the growth bands in the sagittae allowed the identification of thirteen groups (Table 9). The percentage of the otoliths which showed perfectly defined bands of growth was 93.8%. The growth bands can be seen more clearly in the rostrum aspect of the otolith (Fig. 2), where the deposit of aragonite and otolin takes place with more intensity.
Formation Time of the Fast and Slow Growth Rings
In most of the sagittae the fast growth band (opaque) occurs in November and the slow growth band (hyaline) in July, which proves that formation of growth rings takes place once a year.
Scales Description of the Scale
Scales of M. brevipinnis are catenoid and rectangular-shaped; the length is contained around 1.6 times the width. The anterior area is divided in sectors by very marked radios that converge in the focus; their number varies between 6 and 8. The anterior margin presents lobes and in some adults shows serial fissures as distinct bifurcations. The focus is well defined and in an eccentric position due to the faster growth of the anterior area than the posterior area. The lateral borders are smooth. The growth rings are well defined and were interpreted as the formation of a double dark and continuous line defined by a clear and translucent space. The triangular-shaped ctenii are in the rear area and project toward the posterior margin which is thinner and brittle compared with the rest of the scale (Fig. 3).
Growth of the Scale
The relationship between length and width of the scale is expressed by the value of exponent b = 0.885 which corresponds to a negative allometric growth. The determination index of the relationship of the two series of data is R2 = 0.79, with F = 967 of the anova, which indicates that the scale grows more in length than widthwise (Table 7).
| Table 6: | Relationships between the total length of the fish (L) and the measures of the sagittae |
 | |
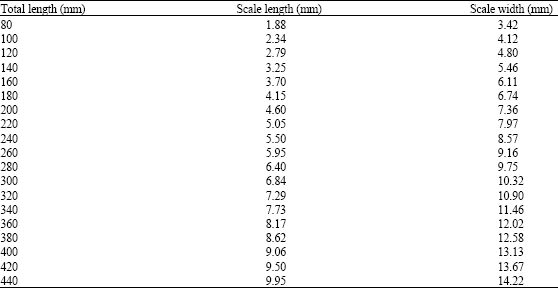
| Table 7: | Relationship between length classes, length and width of the scale of M. brevipinnis |
 | |
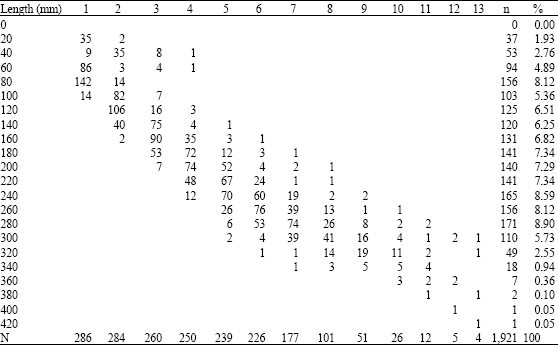
| Table 8: | Age-length distribution of M. brevipinnis with scales |
 | |
The relationship between total fish length and length of the scale is b = 0.977, which is very close to isometry (R2 = 0.483 and anova F = 229.05). This means that there is a high proportionality between the increase of scale length and of fish length; for this reason the scale is suitable to describe the growth in length of the fish.
Identification of the Growth Rings
The analysis of the growth rings in the scales allowed the identification of thirteen groups (Table 9). The percentage of the scales which showed perfectly defined rings of growth was 85%; the rest (15%) were regenerated scales. Table 8 shows the age-length distribution.
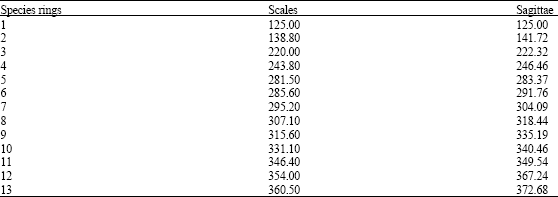
| Table 9: | Mean total length (mm) for each ring identified with scales and sagittae of M. brevipinnis |
 | |
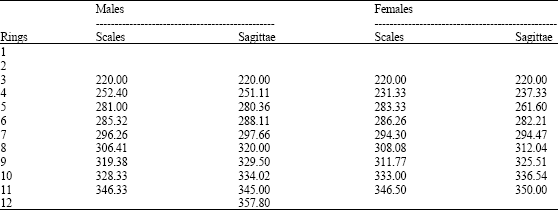
| Table 10: | Mean total length (mm) for each ring identified with scales and sagittae for sexes of M. brevipinnis |
 | |
Determination of the Marginal Increment
Maximum, average and minimum data of the margin between the last ring and the scale border for every month. June was the month with the smallest value of the margin which validates the use of scales to determine the age in this species and its annual rhythm.
Differences Between Sexes
Table 10 shows the differences in the mean total length for each ring for males and females observed by the study of the sagittae and scales. Sexual differentiation is possible from ring 2 on, that is, 220 mm of total length. Data for rings 12 and 13 correspond to 8 individuals that were eviscerated by fishers before landing and their sex could not be determined.
DISCUSSION
Morphometric analysis of the species showed that M. brevipinnis is stouter during the first stage of its life; once it reaches sexual maturity (approximately at 220 mm, Table 10) it becomes slimmer because waste of fatty acids is higher during the spawning season and the individuals’ loose weight. Cruz-Romero et al. (1993) in Manzanillo coast and Espino-Barr et al. (2004a, b) in Jalisco coast, also found negative allometric indexes between standard length and eviscerated weight for mature individuals of M. brevipinnis with values of b = 2.45 and b = 2.53, respectively.
Other members of the Haemulidae family show a negative allometric index of the relationship between standard length and eviscerated weight: Haemulon steindachnerii (b = 2.13), H. flavigutattum (2. 35), Pomadasys panamensis (2.49), Anisotremus caesius (2.65), A. taeniatus (2.71) and Xenichthys xanti (2.76). Other species show an isometric index: H. sexafasciatum (2.91), H. maculicauda (2.99), A. interruptus (3.027, P. elongatus (3.21) and P. bayanus (3.26) (Espino-Barr et al., 2004b). Microlepidotus brevipinnis is in a middle position in the Haemulidae family.
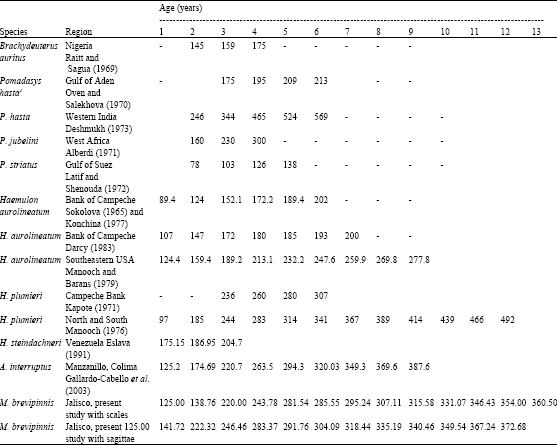
| Table 11: | Mean length (mm) for each age for hemulids. This table was modified from Table 5 in Gallardo-Cabello et al. (2003) |
 | |
| 1males | |
Table 9 shows that the mean length of the organisms with rings 1 to 5 have similar sizes in otoliths and scales and greater differences occur in groups 6 to 13. Ring 9 has the highest difference between these structures, because from 6th ring on there is a decrease in the number of organisms (Table 8) and possibly a higher bias in the data. Both cases show the highest growth rate from 0 to 1 ring, with an increment of 125 mm. From the 2nd ring on, growth rate decreases abruptly. Reduction of organisms from age six on, is due to fishing pressure.
There is very little information on M. brevipinnis, but as a family, Table 11 shows the average lengths for each age of different Haemulidae species in different countries. According to this Table 11, M. brevipinnis reaches the higher age (13 years), followed by H. plumieri (12 years) in North and South Carolina (Manooch, 1976) and A. interruptus in Manzanillo, Colima (Gallardo-Cabello et al., 2003) and H. aerolineatum in the Southeastern USA (Manooch and Barinas, 1979) both with 9 years. Other reports show that Haemulids do not reach more than 7 years of age.
The smaller species are Pomadasys jubelini (3 years) (Alberdi, 1971), H. steindachnerii (3 years) (Eslava, 1991), P. striatum (4 years) (Latif and Shenouda, 1972), Brachydeuterus auritus (4 years) (Raitt and Sagua, 1969) and P. hasta (from Western India, 5 years, Deshmukh, 1973). However P. hasta reaches the largest length of this family (569 mm, Deshmukh, 1973).
Haemulids which reach larger lengths in Jalisco, according to Espino-Barr et al. (2004a) are H. sexfasciatus (650.2 mm), A. interruptus (621.8 mm), P. panamensis (557.5 mm), H. flaviguttatum (492.1 mm), A. caesius (445.6 mm) and M. brevipinnis (442.4 mm). Smaller species of Haemulidae are: P. elongates (430.2 mm), P. bayanus (395.7 mm), A. taeniatus (334.0 mm), H. scudderi (310.0 mm), H. maculicauda (301.0 mm), A. dovii (283.0 mm), H. steindachnerii (274.8 mm), A. pacifici (249.0 mm) and Xenichthys xanti (238.4 mm). According to these results, M. brevipinnis occupies a middle point in the mid Mexican Pacific coast. Species that reach smaller ages as B. auritus, H. steindachnerii, P. hasta, P. jubelini, P. striatum, H. plumieri and H. aurolineatum these last two in Campeche are found in lower latitudes, near the equator, where temperature is higher, which coincides with Taylor’s (1958, 1960), who reported that organisms living in high temperatures reach smaller ages. Conversely Haemulids living in higher latitudes reach older ages, i.e., H. aurilineatum (Southeastern USA., Manooch and Barans 1979), H. plumieri (North and South Carolina, Manooch, 1976), A. interruptus (Manzanillo, Colima, Gallardo-Cabello et al., 2003) and M. brevipinnis (Jalisco, present study).
This study provided information on population structure in length, weight and age classes, necessary for the assessment plans of the fishery.
ACKNOWLEDGMENTS
We want to thank the artisanal fishers from Jalisco, who were very helpful in giving us information and facilitating organisms. Arturo Garcia-Boa and Marcos Puente-Gómez helped in the field and lab., work; also MS Alejandro Gómez-Ponce and Biol. Claudio Padilla-Hernández for their help in the lab. Several institutions gave financial support: UNAM, CONACyT and INP.
REFERENCES
- Alberdi, P.G., 1971. Biologie et Peche du Sompat Pomadasys jubelini (Cuvier, 1830) des Eaux-Senegambiennes. ORSTOM, Dakar, Pages: 30.
Direct Link - Alvarado-Castillo, R. and R. Felix-Uraga, 1998. Crecimiento de Istiophorus platypterus (Pisces: Istiophoridae) en la boca del Golfo de California. Rev. Biol. Trop., 46: 115-118.
Direct Link - Anislado-Tolentino, V. and C. Robinson-Mendoza, 2001. Age and growth for the scalloped hammerhead shark Sphyrna lewini (Griffith and Smith, 1834) along the central Pacific coast of Mexico. Ciencias Marinas, 27: 501-520.
Direct Link - Bearez, P., 1996. Lista de los peces marinos del Ecuador Continental. Rev. Biol. Trop., 44: 731-741.
Direct Link - Berg, E., T.H. Sarvas, A. Harbitz, S.E. Fevolden and A.B. Salberg, 2005. Accuracy and precision in stock separation of North-East Arctic and Norwegian coastal cod by otoliths-comparing readings, image analyses and a genetic method. Mar. Freshwater Res., 56: 753-762.
Direct Link - Campana, S.E., 1999. Chemistry and composition of fish otoliths: Pathways, mechanisms and applications. Mar. Ecol. Prog. Series, 188: 263-297.
CrossRef - Campana, S.E. and S.R. Thorrold, 2001. Otoliths, increments and elements: Keys to a comprehensive understanding of fish populations. Can. J. Fish. Aquat. Sci., 58: 30-38.
CrossRefDirect Link - Espino-Barr, E., M. Gallardo-Cabello, F. Gonzalez-Orozco and A. Garcia-Boa, 2004. Analisis del crecimiento y la mortalidad de Anisotremus interruptus (Gill, 1863) (Perciformes: Haemulidae) en la costa de Colima, Mexico. Bol. Invest. Mar. Cost., 33: 67-75.
Direct Link - Espino-Barr, E., M. Gallardo-Cabello, A. Garcia-Boa, E.G. Cabral-Solis and M. Puente-Gomez, 2006. Morphologic and morphometric analysis and growth rings identification of otoliths: Sagitta, asteriscus and lapillus of Caranx caninus (Pisces: Carangidae) in the Coast of Colima, Mexico. J. Fish. Aquat. Sci., 1: 157-170.
CrossRefDirect Link - Francis, C., S.J. Harley, S.E. Campana and P. Doering-Arjes, 2005. Use of otolith weight in length-mediated estimation of proportions at age. Mar. Freshwater Res., 56: 735-743.
Direct Link - Gallardo-Cabello, M., E. Espino-Barr, F. Gonzalez-Orozco and A. Garcia-Boa, 2003. Age determination of Anisotremus interruptus (Perciformes: Haemulidae) by scale reading, in the coast of Colima, Mexico. Rev. Biol. Trop., 51: 519-528.
PubMedDirect Link - Gallardo-Cabello, M., E.G. Cabral-Solis, E. Espino-Barr and A.L. Ibanez-Aguirre, 2005. Growth analysis of white mullet Mugil curema (Valenciennes, 1836) (Pisces: Mugilidae) in the Cuyutlan Lagoon, Colima, Mexico. Hidrobiologica, 15: 321-325.
Direct Link - Espino-Barr, E., M. Gallardo-Cabello, A. Garcia-Boa, E.G. Cabral-Solis and M. Puente-Gomez, 2006. Morphologic and morphometric analysis and growth rings identification of otoliths: Sagitta, asteriscus and lapillus of Caranx caninus (Pisces: Carangidae) in the coast of Colima, Mexico. J. Fish. Aquat. Sci., 1: 157-170.
CrossRefDirect Link - Madrid-Vera, J., A. Ruiz-Luna and I. Rosado Bravo, 1998. Peces de la plataforma continental de Michoacan y sus relaciones regionales en el Pacifico Mexicano. Rev. Biol. Trop., 42: 267-276.
Direct Link - Mariscal-Romero, J. and A.M. van der Heiden, 2006. Peces de Importancia Ecologica y Comercial Asociados a Fondos Blandos en la Plataforma Continental de Jalisco y COLIMA, Mexico. In: Los Recursos Pesqueros y Acuicolas de Jalisco Colima y Michoacan, Jimenez-Quiroz and M.E. Espino-Barr (Eds.). INP, SAGARPA, UK., pp: 180.
- Penailillo, J.P. and M.C. Araya, 1996. Formation moment and periodicity of the growth microincrements in the otoliths of pejerrey larvae Austromenidia regia maintained in the laboratory. Invest. Mar., 24: 31-38.
Direct Link - Raitt, D.S.F. and V.O. Sagua, 1969. Preliminary investigations on the biology of Brachydeuterus auritus (Val., 1831), in Nigerian waters. Proceedings of the Symposia Oceanographyc and Fisheries Resources of the Tropical Atlantic, (SOFRTA'69), UNESCO. Iveory COSAT. Nigeria, pp: 397-401.
Direct Link - Ruiz-Dura, M.F. Y. Orijel-Arenas and G.H. Rodriguez, 1970. Lineas de Crecimiento en Escamas de Algunos Peces de Mexico. Instituto Nacional de Investigaciones Biologico Pesqueras, Mexico, pp: 97.
Direct Link - Taylor, C.C., 1960. Temperature, growth and mortality-the Pacific cockle. J. Conseil Int. Explor. Mer., 26: 117-124.
CrossRef