
Y. Norma-Rashid
Institute of Biological Sciences, Faculty of Science, University of Malaya, 50603 Kuala Lmpur, Malaysia
N.A. Rahman
Institute of Biological Sciences, Faculty of Science, University of Malaya, 50603 Kuala Lmpur, Malaysia
D. Li
Department of Biological Sciences, National University of Singapore, 119260, Singapore
International Journal of Zoological Research
Year: 2009 | Volume: 5 | Issue: 1 | Page No.: 9-15







ABSTRACT
This study examines the diversity and distribution of spiders in the mangrove areas in Peninsular Malaysia which is hoped to contribute towards understanding the dynamics of the mangrove ecosystem. Little research has been done on local spiders and inventory from this habitat is rare. From faunal samples taken along transects from upper to lower zones of Morib mangrove forest and studies on available museum specimens in the Raffles Museum of Biodiversity Research (RMBR), yielded 26 species belonging to 10 family groups. There exist 4 species as new records for Malaysia which are Clubiona meraukensis, Dolomedes mizhoanus, Pardosa zhanjiangensis and Telamonia dimidiata. The jumpers or family Salticidae has an overall wide distribution across all zones sampled. Generally, the spider distribution depended on the zone location and floral diversity. Spider zonation pattern is probably influenced by complex factor combinations rather than one factor which include biotic relationships such as competition and predation.
PDF Abstract XML References Citation
How to cite this article
Y. Norma-Rashid, N.A. Rahman and D. Li, 2009. Mangrove Spiders (Araneae) of Peninsular Malaysia. International Journal of Zoological Research, 5: 9-15.
DOI: 10.3923/ijzr.2009.9.15
URL: https://scialert.net/abstract/?doi=ijzr.2009.9.15
DOI: 10.3923/ijzr.2009.9.15
URL: https://scialert.net/abstract/?doi=ijzr.2009.9.15
INTRODUCTION
Mangroves are unique inter-tidal habitats that have been rapidly destroyed for development of coastal zones throughout the tropical world. Damage to the system can result in irreversible coastal erosion. Malaysia has 4,800 km coastline and 15% of this is lined with areas of mangrove in character (Lim, 2007) of which much of that are scarred by adjacent developments. Reclamation of the mangrove swamps for various development projects means destruction of the Rhizophora and Brugeria zones. However, the future of local mangroves may not be so bleak. In Malaysia there have been several attempts at replanting of mangroves with varying degree of success. Bamboo poles were used in the seventies to hold the mud while waiting for the mangroves to establish themselves. More recently, Lim (2007) reported the formulation of a feasible concept using sausage shaped geotubes named Pillowtubes designed to absorb the wave actions have been utilized to protect not only mangrove seedlings from being washed to sea but enabling young mangrove saplings to sprout and take root within the mudflats. There are suggestions from the public that the existing 100,000 ha of mangrove remains should be gazetted as protected areas.
The diversity of mangrove plants of Peninsular Malaysia comprised of 33 true and 36 transitional species (Wee and Corlett, 1986). Mangroves colonize the coastal mudflats in a series of succession (Berry, 1963; Macnae, 1968; Liow, 2000). The pioneer species are the Avicennia and once this zone has established itself, the landward mangrove belt will have low salinity due to the landward influence giving rise to the succession of Rhizophora and the Brugeria species. Erosion of the muddy coasts are damaging, causing the roots of the mangroves fringing the sea (Avicennia) to be exposed which eventually collapse. Erosion is slowed down in the Rhizophora and Brugeria zones which have prop roots that anchor deep into the mud to withstand the natural forces.
Retaining the mangroves has the added bonus of preserving the fauna that depends on these habitats. Of late issues pertaining to man-induced extinction of species have increasingly gained importance in a global context. Ong (1994) highlighted the paucity of baseline data pertaining to mangroves which still harbors wildlife. The Southeast Asian mangroves are said to be biologically richer than anywhere else in the world. Realization to conserve wildlife, including those found in the mangroves is inevitable (Liow, 2000). A baseline data is urgently needed for the mangroves whether it is taxonomic, community, ecological or biochemical. The lack of information impedes impact assessment, proper management and contingency plans for the faunal component of the mangroves (Liow, 2000).
Spiders are present in local mangrove forests around Malaysia but virtually nothing is known about their specific diversity and ecology. Some pertaining information can be found embedded within work done by Koh (1991) dealing with araneids in the mangrove and Ng and Sivasothi (1999). The present study, therefore, takes this into account and was done to provide necessary quantitative observations of the diversity and distribution of spiders in mangrove forests. The findings will assist in providing the background against which any future environmental deterioration in specifically Malaysian coastal mangrove forests might be assessed.
MATERIALS AND METHODS
Two types of studies were conducted for this work. For the spider diversity aspect besides field sampling that was conducted in the mangrove areas of Morib, Malaysian specimens that are available in the Raffles Museum of Biodiversity Research (RMBR), National University of Singapore (NUS) were examined in October 2006. Data on spiders zonation and distribution were collected during a field survey in Morib, Selangor, Peninsular Malaysia. Morib is situated about 50 km south of Port Klang on the west coast of Peninsular Malaysia (2° 44′N, 101° 28′E). Three rivers, Sungai Langat, Sungai Morib and Sungai Kancong drain into the sea near the study site. Manual sampling was done in 12 successive sessions beginning in June 1997 during low tides.
The sampling zonation within the mangrove areas in Morib can be described and divided into 1 to 5 major zones and within zone 2 can further be subdivided into 3 subzones that is 2a-c (Table 1). Manual samplings and the beating method were conducted within line transects of 100 m in all major and subzones. The number and species of spiders in each major and subzones were recorded. Due to accessibility reasons arachnids were sampled at heights of 1.65 m and below.
The statistical software program statgraphic Version 2.6 was used for data analysis. To test for significant numerical differences the t-test was used. Cluster analysis was also employed to test the similarity in distribution of spider species between zones.
| Table 1: | Descriptive and sampling zones through the Mangrove forest in Morib, Peninsular Malaysia |
 | |
RESULTS
The mangrove trees have different animals living upon them in differing abundances at various horizontal and vertical zonations. Other than the arachnids, the insects present in field samplings, belonged to the Order Coleoptera, Diptera, Hemiptera, Homoptera, Hymenoptera, Lepidoptera, Mantoidea, Neuroptera, Odonata and Orthoptera. Identification of spiders from field collections and museum work, yielded 27 species belonging to 10 family groups (Table 2). The country distributions where these species are also found are shown in Table 2. Presence of these species in the mangrove habitat had been reported in other references (Table 2) except for 4 species considered as new records for Malaysia. These new records are Clubiona meraukensis, Dolomedes mizhoanus, Pardosa zhanjiangensis and Telamonia dimidiata.
The proportional abundance of the spiders in the field in descending order as indicated in the pie chart (Fig. 1) are as follows: the most dominant in numbers are the jumping spiders, Salticidae, comprised of 42% followed by the lynx spiders, Oxyopidae with 25%, in moderate numbers are the nursery web spiders, Pisauridae (16%) and long jawed spiders, Tetranagthidae (11%) and smaller densities are the daddy long legs, Pholcidae (4%) and Psecridae (2%).
| Table 2: | Diversity list of mangrove spiders found in specific areas in Peninsular Malaysia and distribution within South East Asia reported as in references indicated |
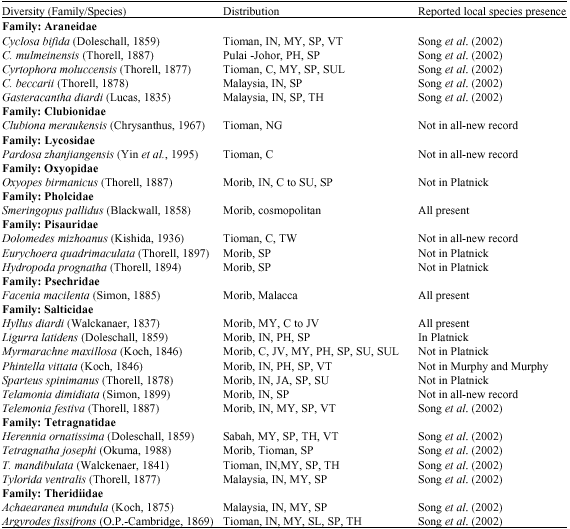 | |
| Named localities within Malaysia are given in full while other countries abbreviated as: C: China, IN: Indonesia, JV: Java, MY: Myanmar, NG: New Guinea, PH: Phillipines, SL: Sri Lanka, SP: Singapore, SU: Sumatra, SUL: Sulawesi, TH: Thailand, TW: Taiwan and VT: Vietnam). The presence of reported local species besides from our current study are also in reference to Murphy and Murphy (2000), Ng and Sivasothi (1999), Platnick (2006) and Song et al. (2002) | |
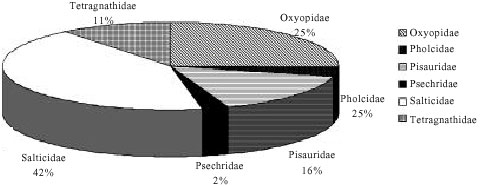 | |
| Fig. 1: | The overall percentage (%) availability of spider family groups found from the study site in Morib, Peninsular Malaysia |
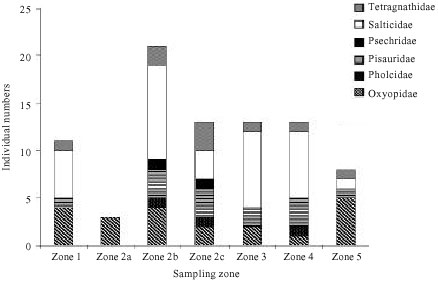 | |
| Fig. 2: | The frequency distribution of spider family groups (individual numbers) across species found in all sampling zones (zone 1 to 5) in the mangrove forest of Morib, Peninsular Malaysia |
There was a great variation in the diversity and densities of spiders among zones (Fig. 2). The greatest density and diversity of species occurred in the middle zones (zones 2b and c) with the greatest faunal heterogeneity rather than other zones of the forest especially which contained homogeneous mangrove trees of Avicennia species. This is shown by the marked difference (t-test: t(1,5) = 6.32456, p = 0.0015) between mixed forest zone (zone 2b) and monogeneous Avicennia trees, adjacent to the sandy beach (zone 3). Generally, the density depended on the zone location and floral diversity. Interestingly, the salticids occurred in all zones of the mangrove forest.
The spider fauna in all zones were also compared by means of cluster analysis. In reference to the associated fauna and most dissimilar point, 3 main clusters are suggested (Fig. 3). This revealed similarities in components of zones 2a and 5, these are neighboring areas of Avicennia forests. Zones 3 and 4 are more linked in their spider components although comprised of different floral species but they are areas fringing open habitats and closely associated to human presence. Zone 2c is an outgroup which distinctly differed in spider species contents.
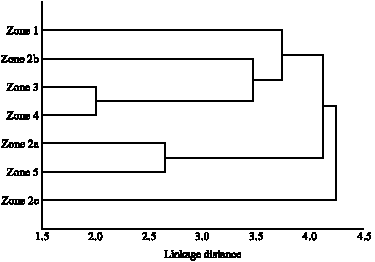 | |
| Fig. 3: | A dendrogram formed using cluster analysis on the spiders found in zones 1 to 5. The scale shows numerical similarities |
DISCUSSION
Berry (1972) stated that mangroves bear a fascinating fauna whose study can be highly rewarding to the hardy naturalist, ecologist or other scientists. According to him the fauna can be broadly divided into aquatic and terrestrial components in which spiders are included in the terrestrial fauna living in the mangrove trees and thus revealed a vertical zonation of the fauna from those in the soil up to those in the higher parts of the trees. Berry (1972) stated the presence of Oecophylla smaragdina above 9.5 feet but with no mention of spiders in the vertical zonation range that was sampled. Sasekumar (1974) briefly mentioned (no named species) the availability of the mangrove spiders which he categorized them as the tree fauna confined to the vegetation.
Liow (2000) highlighted the invertebrates being the least known in the mangroves, thus giving rise to potentially new findings. Here, four new records of spiders are reported for Peninsular Malaysia although it is difficult to conclude whether these species are confined exclusively to the mangroves due to sparse of data from previous local work or reports. The extensive leafy canopy of the mangrove forests provide a cool, stable and shaded environment subjected to great humidity for faunal colonization (Sasekumar, 1974; Ross and Underwood, 1997). This is supported by Macnae (1968), who reported that mangroves are infested with mosquitoes and midges (often mistaken for sandflies), bees visited mangrove flowers, termites invaded dead wood together with cockroaches and beetles while canopy dwellers such as ants, spiders with fireflies twilight aggregations. The canopy provided shelter for spider retreat which otherwise would expose them to greater risk of desiccation. Berry (1972) reported, other than insects, organisms on trunks of trees were abundant from the surface of the mud to 40 cm upwards with oysters, barnacles, limpets and grazing littorinid snails on mangrove trunks.
The composition and properties of mangrove flora may affect the distribution and abundance of spiders analogous to faunal zonation with possible dependent variables such as, increasing distance from the seaward edge of the forest, height above low tidal level, orientation of the substratum and biotic interactions such as competition or predatory relationships. Macnae (1968) stated that mangroves are limited to a few dominant groups, here the jumping spiders, Salticidae are predominant and widespread throughout all zones, this is incontrast to work reported by Macintosh and Ashton (2002), who found abundance of web building spiders and Koh (1991) discovered one new species, Argiope mangal (Family: Araneae) in Singapore mangroves. The success of jumping spiders can be attributed to their known predatory habits, aggressively stalking for wide ranging prey and the unrestricted foraging strategies from canopy to the exposed ground when tide levels are low. This kind of active hunting for prey is in concordance to the wolf spiders, Pardosa known to adapt as semi aquatic life form and exploiting the juveniles of fiddler crab as food resources (Macintosh and Ashton, 2002). In other animals, Maza and Soberon (1998) revealed that the multi taxonomic groups of butterflies species having distinct morphological and ethological features are consistently associated with certain habitats.
Sasekumar (1974) described the distribution of macrofauna and recognized their zonation in distribution. According to Macnae (1968), the widest zones in the mangroves are the forested areas of Bruguiera which are separated from the sea by Avicennia or Sonneratia fringes. Here, the physical environment is potentially less severe due to the canopy of the trees with extensive root growths and restricted movement of water (Ross and Underwood, 1997). Such conditions provided suitable living quarters for animals to thrive as shown in this study, where, maximum spider densities are found in the middle zones of mixed forests (zones 2b and c). In contrast, the open zone closest to the seaward edge (zone 5) appeared as rather a harsh environment, too poor in fauna or flora to be of interest (Berry, 1972), here shown, able to support a minimum spider community especially the more hardy species of salticids, lynx, long jawed and nursery web spiders. Thus, there exist clear spatial patterns of spiders in the mangrove forests. Understanding the dynamics and components of mangrove ecosystem is crucial from many points of view especially now when national development is depleting and encroaching into this fragile environment. Lim (2007) quoted it takes more than good intentions to put the groove back into mangroves. One likely response to this would be more data needed, more actions wanted and only then more mangroves saved.
ACKNOWLEDGMENTS
We wish to thank all involved that has made this study possible. The first author wishes to acknowledge University of Malaya for financial aid and National University of Singapore for sabbatical attachment and especially staffs of RMBR and members of the Spider Laboratory for help, support and many memories to treasure.
REFERENCES
- Berry, A.J., 1972. The natural history of West Malaysian mangrove faunas. Malaya Nat. J., 25: 135-162.
Direct Link - Liow, L.H., 2000. Mangroves conservation in Singapore: A physical or a psychological impossibility? Biodivers. Conserv., 9: 309-332.
Direct Link - Macnae, W., 1968. A general account of the fauna and flora of mangrove swamps and forests in the Indo-West-Pacific region. Adv. Mar. Biol., 6: 73-270.
CrossRefDirect Link - De La Maza, R. and J. Soberon, 1998. Morphological grouping of Mexican butterfliesin relation to habitat association. Biodivers. Conserv., 7: 927-944.
CrossRefDirect Link - Ross, P.M. and A.J. Underwood, 1997. The distribution and abundance of barnacles in a mangrove forest. Aust. J. Ecol., 22: 37-47.
CrossRefDirect Link - Sasekumar, A., 1974. Distribution of macrofauna on a Malayan mangrove shore. J. Anim. Ecol., 43: 51-69.
Direct Link