
O.A. Al- Dokhi
Department of Zoology, College of Science, King Saud University, P.O. Box 2455, Riyadh-11451, Saudi Arabia
International Journal of Zoological Research
Year: 2009 | Volume: 5 | Issue: 4 | Page No.: 136-149







ABSTRACT
The present study aimed to describe the morphogenesis of the acrosomal vesicle during spermiogenesis in the lizard Ptyodactylus hasselquisti. Five adult male lizards were captured during the period of sexual activity (April and May, 2005) from old houses in the city of Riyadh (25° 10' N, 46° 50' E), Saudi Arabia. Ultrastructural examination revealed proliferation and hypertrophy of Golgi complex elements as the initial event in formation of the acrosomal vesicle. Numerous small vesicles were released from Golgi complex and then coalesced to form a large proacrosomal vesicle which later attached to the proximal surface of spermatid nucleus. A proximal concavity appeared in the spermatid nucleus to completely lodge the acrosomal vesicle, then the spermatid nucleus with the lodged vesicle were transported to be directly apposed to the spermatid plasmalemma. This was associated with the appearance of a single acrosomal granule at the vesicle base. Subsequently, the progressively pushed acrosomal vesicle was flattened on the proximal nuclear surface. Some acrosomal vesicle deformities were also illustrated. The successive morphogenetic stages of the acrosomal vesicle were discussed in comparison with that reported in the previous studies concerned with spermiogenesis in other reptile species.
PDF Abstract XML References Citation
How to cite this article
O.A. Al- Dokhi, 2009. Morphogenesis of the Acrosomal Vesicle During Spermiogenesis in the House Gecko
Ptyodactylus hasselquisti (Squamata, Reptilia). International Journal of Zoological Research, 5: 136-149.
DOI: 10.3923/ijzr.2009.136.149
URL: https://scialert.net/abstract/?doi=ijzr.2009.136.149
DOI: 10.3923/ijzr.2009.136.149
URL: https://scialert.net/abstract/?doi=ijzr.2009.136.149
INTRODUCTION
Spermiogenesis is the process by which haploid spermatids (gametes produced by meiotic division) are transformed into mature spermatozoa (Hiatt and Gartener, 1997; Stevens and Lowe, 1997; Young and Heath, 2002).
The data derived from the ultrastructural studies on reptile's spermiogenesis have clarified some aspects of fertilization and egg activation (Teixeira et al., 1999). These ultrastructural data are also among the contributors of phylogenetic analysis (Jamieson et al., 1995, 1999; Teixeira et al., 1999). Ultrastructure of spermiogenesis and spermatozoa of lizards was the target of considerable number of studies (Clark, 1967; Furieri, 1970, 1974; Del Conte, 1976; Landim and Hoffling, 1977; Butler and Gabri, 1984; Courtens and Depeiges, 1985; Al-Hajj et al., 1987; Dehlawi and Ismail, 1990, 1991; Dehlawi, 1992; Dehlawi et al., 1993; Teixeira et al., 1999; Ferreira and Dolder, 2002, 2003; Al-Dokhi, 2004; Mubarak, 2004; Vieira et al., 2004; Al-Dokhi, 2006).
Formation of the acrosomal vesicle is one of the essential morphological transformations during the process of spermiogenesis (Landim and Hoffling, 1977; Butler and Gabri, 1984; Al-Hajj et al., 1987; Mubarak, 2004; Al-Dokhi, 2006). However, detailed ultrastructural description of the acrosomal vesicle development in the lizard species has not yet been reported. Therefore, the present study has been intended to elucidate the morphogenesis of the acrosomal vesicle during spermiogenesis in the lizard Ptyodactylus hasselquisti (P. hasselquisti). The obtained data would be helpful with the other morphological studies to form a base for phylogenesis of lizard species.
Attention has been paid to compare the obtained results with that reported in the previous studies which are concerned with spermiogenesis in other lizard species.
MATERIALS AND METHODS
Five adult males of the lizard P. hasselquisti were captured during the period of sexual activity (April and May 2005), from old houses in the city of Riyadh (25° 10' N, 46° 50' E), Saudi Arabia. The lizards were dissected and their testes were removed, diced into proper small pieces (1 mm cubes) and immediately fixed by immersion in 3% buffered glutaraldehyde (0.1 M sodium cacodylate buffer at pH 7.2) for 4 h at 4°C. The fixed tissue specimens were thoroughly washed in the same buffer and then post-fixed in 1% osmium tetroxide (OsO4) in 0.1 M sodium cacodylate buffer (pH 7.2) for 2 h. Tissue specimens were subsequently dehydrated in ascending grades of ethanol and transferred to epoxy resin (Epon/Araldite mixture) via propylene oxide. Thin sections (70-80 nm) were cut with a diamond knife on an ultramicrotome (Leica, UCT), double stained with uranyl acetate and lead citrate and examined under a transmission electron microscope (JEOL, JEM1011) operating at 80 KV.
RESULTS
The differentiating spermatids occupied the adluminal compartment of the seminiferous epithelium. At the early stage of spermiogenesis, spermatids of P. hasselquisti displayed prominent juxtanuclear Golgi complex which consisted of flattened cisternae and many associated varied sized vesicles (Fig. 1). The cytoplasm of these early spermatids also contained abundant smooth endoplasmic reticulum, but scarce rough endoplasmic reticulum represented by flattened cisternae studded by tiny ribosomal granules. Mitochondria, ovoid or elliptical with linear cristae, were distributed throughout the spermatid cytoplasm. The content of the spermatid rounded nucleus was mostly a homogenous fine granular chromatin (euchromatin), with the presence of small patches of dense chromatin (heterochromatin) that were restricted to the inner side of the nucleolemma. The early spermatids were still rounded in shape. The first morphological event in the differentiating spermatids was the hypertrophy of Golgi complex and proliferation of its elements as represented by increasing number and size of its cisternae and associated vesicles (Fig. 2). The active Golgi complex was in a close contact with the spermatid nucleus. The associated Golgi complex vesicles continued to increase in size and subsequently a large voluminous vesicle developed, most likely through coalescence of many smaller vesicles (microvesicles) (Fig. 3). The resultant macrovesicle represented a proacrosomal vesicle which contained a flocculent material of low electron density.
With progression of the differentiation process, the proacrosomal vesicle moved to attach the spermatid nuclear membrane (Fig. 4), with the formation of an electron dense material at the site of attachment. This electron dense material probably represented the initial appearance of a subacrosomal material between the proacrosomal vesicle and the nuclear membrane.
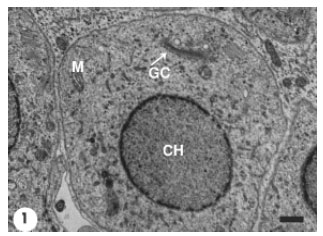 | |
| Fig. 1: | Early spermatid showing prominent Golgi Complex (GC). Nuclear chromatin (CH) is uniformly fine granular (euchromatin), except for small chromatin clumps (heterochromatin) on the inner nuclear membrane. Scale bar = 1 μm |
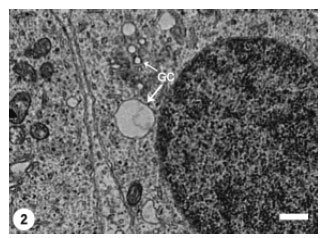 | |
| Fig. 2: | Early spermatid manifesting conspicuous proliferated Golgi Complex (GC) which involves varied sized vesicles at a juxtanuclear location. Scale bar = 1 μm |
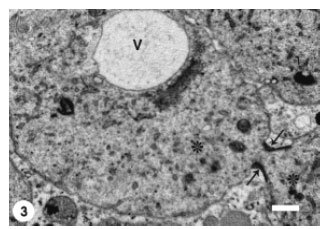 | |
| Fig. 3: | Large voluminous vesicle (V) (macrovesicle) formed as a result of coalescence of smaller Golgi complex vesicles (microvesicles). There is an intercellular bridge (arrows) connecting the two spermatids (*). Scale bar = 1 μm |
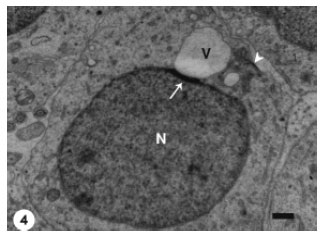 | |
| Fig. 4: | A macrovesicle (V) (proacrosomal vesicle), arised from Golgi complex, attached to the spermatid nucleus (N). An electron dense material (arrow) has been formed at the site of attachment. Golgi complex (arrowhead) is still in a close contact with the spermatid nucleus. Scale bar = 1 μm |
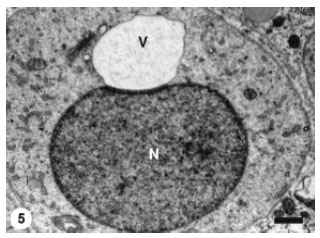 | |
| Fig. 5: | The proacrosomal vesicle (V) has enlarged and exerted more pressure on the spermatid nucleus (N) which manifests a slight concavity. The vesicle (V) contains a flocculent material. Scale bar = 1 μm |
The nuclear surface at the site of attachment to the proacrosomal vesicle was relatively flattened. The proacrosomal vesicle was still in intimate contact with the active Golgi complex. While it was attached to the spermatid nucleus, the proacrosomal vesicle continued to enlarge and Golgi complex microvesicles were discerned joining the proacrosomal vesicle membrane (Fig. 5).
The previously flattened nuclear surface showed slight concavity to accommodate the enlarging proacrosomal vesicle.
At a later stage, the acrosomal vesicle became a large outstanding structure occupying a significant proportion of the spermatid cytoplasm (Fig. 6) and contained more flocculent material as well as remnants of degraded membranous structures. The proximal concavity of the spermatid nucleus was increased obviously and the subacrosomal material became distinct. A large number of microvesicles were in a close association with the attached acrosomal vesicle. With advancing of spermatid differentiation, the nuclear concavity was noticeably increased to the extent that the acrosomal vesicle was completely lodged into the spermatid nucleus (Fig. 7) which was still regularly rounded.
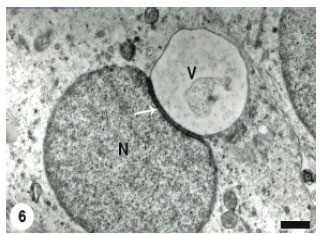 | |
| Fig. 6: | The proacrosomal vesicle (V) has more enlarged and the proximal concavity of the spermatid nucleus (N) has increased. The dense material (arrow), between the vesicle membrane and the spermatid outer nuclear membrane, became more obvious. Scale bar = 1 μm |
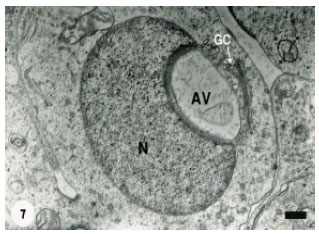 | |
| Fig. 7: | The Acrosomal Vesicle (AV) has been completely lodged into the proximal concavity of the spermatid nucleus (N). Golgi Complex (GC) is in a close contact with the acrosomal vesicle. Scale bar = 1 μm |
At that stage, Golgi complex was remarkably active as evidenced by the presence of numerous released microvesicles. The acrosomal vesicle was progressively expanded to approach the spermatid periphery (Fig. 8) and subsequently it was pushed against the spermatid plasma membrane (Fig. 9).
The higher magnifications revealed the direct apposition of the acrosomal vesicle membrane to spermatid plasmalemma, as well as the compression of the vesicle (Fig. 10). Scattered electron dense varied sized granules appeared within the flocculent material contained in the acrosomal vesicle. The subacrosomal material was discerned as linear dense small patches arranged between the vesicle membrane and the spermatid nuclear membrane.
Compression of the acrosomal vesicle against the spermatid plasma membrane was progressed and the subsequently flattened vesicle was recognized covering a larger area on the proximal spermatid nuclear surface (Fig. 11).
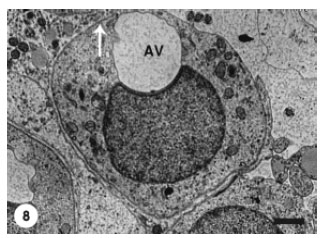 | |
| Fig. 8: | The Acrosomal Vesicle (AV) has expanded to approach the spermatid plasma membrane (arrow). Scale bar = 1 μm |
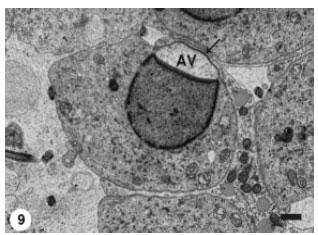 | |
| Fig. 9: | The Acrosomal Vesicle (AV) membrane has become directly apposed to the spermatid plasma membrane (arrow). Scale bar = 1 μm |
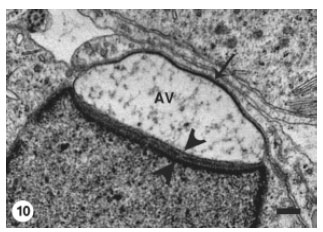 | |
| Fig. 10: | Higher magnification showing the direct apposition of the Acrosomal Vesicle (AV) membrane to the spermatid plasma membrane (arrow). The acrosomal vesicle is apparently compressed. The subacrosomal material (arrowheads) became more distinct and appears as linear dense patches. Scale bar = 1 μm |
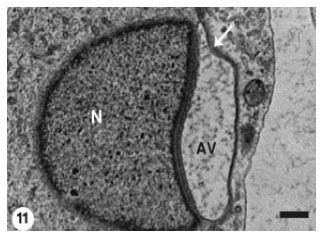 | |
| Fig. 11: | The Acrosomal Vesicle (AV) has been more compressed against the spermatid plasma membrane (arrow). The flattened vesicle covers larger area on the proximal surface of the spermatid nucleus (N). The vesicle still contains flocculent material. Scale bar = 1 μm |
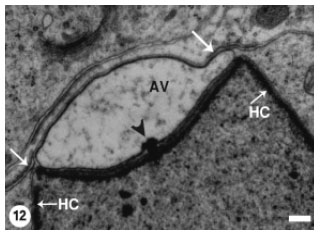 | |
| Fig. 12: | The Acrosomal Vesicle (AV) has been markedly pushed against the spermatid plasma membrane (arrows). A small acrosomal granule (arrowhead) has appeared at the base of the acrosomal vesicle. Heterochromatin (HC) is densely packed as linear patches on the inner nuclear membrane. Scale bar = 1 μm |
This morphological change coincided with the appearance of a small dense granule (acrosomal granule) on the base of the acrosomal vesicle (Fig. 12). Flattening of the acrosomal vesicle was associated with evident increase in the size of the acrosomal granule which was also gradually flattened (Fig. 13). The later stage spermatids disclosed more compression of the acrosomal vesicle and relative elongation of the spermatid nucleus (Fig. 14).
Some spermatids showed a prominent acrosomal granule at the base of a non-flattened lodged acrosomal vesicle (Fig. 15). However, such vesicles were not apposed to the plasma membrane and located free within spermatid cytoplasm. A small number of the growing spermatids disclosed acrosomal vesicles at different morphogenetic stages, for instance some had flattened acrosomal vesicles with prominent acrosomal granule and others possessed less flattened vesicles with absence of the acrosomal granule (Fig. 16). Generally, the growing spermatids displayed similar stage of acrosomal vesicle development, e.g., spermatids showing acrosomal vesicles that were strongly pushed against the spermatid plasmalemma (Fig. 17).
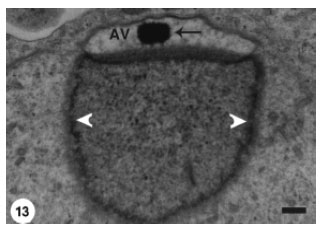 | |
| Fig. 13: | The acrosomal granule (arrow) has enlarged and appears as a dense body. The shape of the acrosomal granule conforms with that of the compressed Acrosomal Vesicle (AV). Heterochromatin (arrowheads) is still condensed on the inner nuclear membrane. Scale bar = 1 μm |
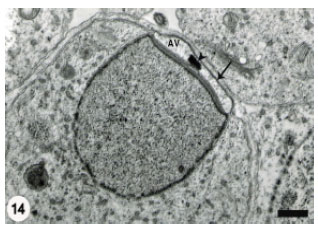 | |
| Fig. 14: | The Acrosomal Vesicle (AV) has been highly attenuated over the proximal nuclear surface. The acrosomal granule (arrowhead) has been also flattened. Scale bar = 1 μm |
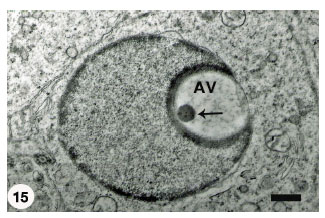 | |
| Fig. 15: | A rounded Acrosomal Vesicle (AV) containing prominent acrosomal granule (arrow) resting on its base and has lodged completely in the proximal nuclear concavity. There is no apposition between the acrosomal vesicle and the spermatid plasma membrane (arrowhead). Scale bar = 1 μm |
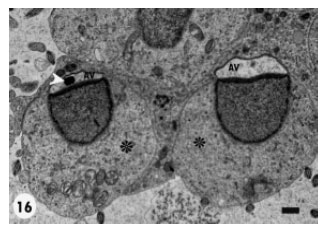 | |
| Fig. 16: | Spermatids (*) showing pushing and apposition of their Acrosomal Vesicle (AV) against the corresponding plasma membranes. The vesicle on the left is more flattened and contains distinct acrosomal granule (arrowhead) and that on the right is less flattened with absence of apparent acrosomal granule. Scale bar = 2 μm |
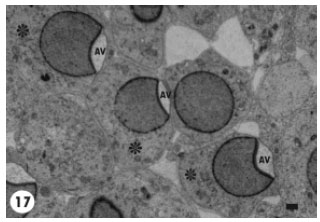 | |
| Fig. 17: | A number of growing spermatids (*) showing Acrosomal Vesicles (AV) at the same morphogenetic stage. The acrosomal vesicles are directly apposed to the spermatids plasma membranes and flattened over the proximal nuclear surfaces. Scale bar = 1 μm |
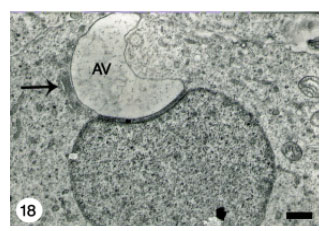 | |
| Fig. 18: | A crescent-shaped Acrosomal Vesicle (AV) in a close contact with Golgi complex (arrow) within a growing spermatid. Scale bar = 1 μm |
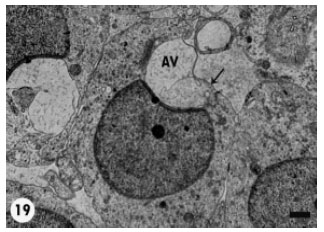 | |
| Fig. 19: | A poorly defined Acrosomal Vesicle (AV) with no distinction between its membrane and the spermatid plasma membrane (arrow). Scale bar = 1 μm |
Relatively few spermatids displayed acrosomal vesicle deformities such as crescent-shaped vesicles and the poorly-defined ones (Fig. 18, 19).
DISCUSSION
The present study describes the developmental morphology of the acrosomal vesicle in the lizard P. hasselquisti. The currently described acrosomal vesicle morphogenesis is in accordance with the general events of sperm head differentiation reported in other lizard species (Landim and Hoffling, 1977; Butler and Gabri, 1984; Courtens and Depeiges, 1985; Al-Hajj et al., 1987; Mubarak, 2004; Al-Dokhi, 2006). Acrosomal vesicle has been reported as an initial feature of sperm head differentiation in reptiles (Butler and Gabri, 1984; Courtens and Depeiges, 1985; Dehlawi and Ismail, 1990, 1991; Dehlawi, 1992).
The initial event during the currently described acrosomal vesicle morphogenesis was the hypertrophy and proliferation of Golgi complex elements which are known to be integrated in packaging and transporting of the synthesized intracellular materials (Stevens and Lowe, 1997). In case of acrosomal vesicle, the accumulated substances within the vesicle are of an enzymatic nature, mainly hydrolytic enzymes (Young and Heath, 2002), since the acrosomal vesicle, which is considered a giant lysosome, is the precursor of the sperm acrosomal cap. The latter structure mainly acts during fertilization to disaggregate corona radiate cells and dissolve the zona pellucida by mean of its hydrolytic enzymes, principally hyaluronidase.
The noticed close association between the acrosomal vesicle and Golgi complex elements obviously points to the origin of the vesicle from Golgi complex. It was evident in the present morphogenetic process that the proacrosomal vesicle had been formed as a result of fusion of the smaller vesicles released from the trans face of Golgi (exit face). Golgi complex cisternae and the associated vesicles were in a continuous intimate contact with the resultant proacrosomal vesicle and the subsequent acrosomal one. This observation confirms that the development of pro- and acrosomal vesicles requires continuous supply of the substances released from Golgi complex. This seems to be essential to increase the vesicle size and also to accentuate the concentration of the vesicle contents.
Attachment of the proacrosomal vesicle to the spermatid nucleus coincided with the appearance of the proximal nuclear concavity, which was most probably developed as a result of the pressure exerted by the proacrosomal vesicle on the spermatid nucleus. According to the present observation, the spermatid nuclear concavity is crucial for lodgment of the acrosomal vesicle.
The present results demonstrated that the spermatid nucleus and the lodged vesicle had been transported from the cell center toward the cell periphery to appose the vesicle membrane to the spermatid plasmalemma. This intracellular translocation process was probably mediated by the cytoskeletal system, especially the microtubular component. Microtubules have the ability to attach to membranous organelles (e.g., mitochondria, vesicles), providing a mean by which such organelles can be moved about within the cytoplasm (Young and Heath, 2002). In other words, the microtubular system is concerned with the transportation processes within the cell cytoplasm. The present results revealed that flattening of the lodged acrosomal vesicle on the spermatid nuclear surface had taken place while the vesicle was pushed against the spermatid plasmalemma. Flattening of the acrosomal vesicle has been demonstrated as the initial event in shaping of the acrosomal cap during spermiogenesis in the lizard species (Landim and Hoffling, 1977; Butler and Gabri, 1984; Courtens and Depeiges, 1985; Al-Hajj et al., 1987; Mubarak, 2004; Al-Dokhi, 2006).
It has been noticed in the present study that changing of the electron density of the acrosomal vesicle occurs after flattening of the vesicle. In mammalian spermatids, the electron translucent acrosomal vesicle enlarges and then condenses and its content is transformed to an electron dense material (Lin and Jones, 2000). We suppose that the nuclear pressure on the acrosomal vesicle shares greatly in changing the vesicle electron density through reducing its size with subsequent appearance of the acrosomal granule.
Presently, the initial appearance of the acrosomal granule has been noticed while the acrosomal vesicle was flattened on the spermatid nucleus. The appearance of the acrosomal granule is most probably the end result of a continuous concentration process of the vesicle flocculent material. It has been reported that acrosomal granule represents the condensed hydrolytic enzymes previously dispersed in the acrosomal vesicle (Hiatt and Gartener, 1997). Currently, a single large acrosomal granule was observed at the base of the acrosomal vesicle. Presence of such single granule is a common finding in reptiles (Clark, 1967; Furieri, 1974; Del Conte, 1976; Butler and Gabri, 1984). However, multiple acrosomal granules have been recognized in some reptilian species such as Scincus mitranus (Al-Dokhi, 1996) and Agama adramitana (Dehlawi et al., 1992). The single acrosomal granule gives rise to an acrosomal medulla of uniform density in the sperm head, while the multiple granules after dissolution result in acrosomal medulla of varied density. In accordance with the present data, many other reptiles develop the acrosomal granule after lodgment of the acrosomal vesicle to spermatid nucleus (Dehlawi et al., 1990; Al-Dokhi, 2006). However, the lizard Stenodactylus selvini contradicts this finding since it manifests the acrosomal granule before the mentioned lodgment (Dehlawi et al., 1991).
Similarly, autherian mammals develop the acrosomal granule within the early acrosomal vesicle before its lodgment to the spermatid nucleus (Holstein and Roosen-Runge, 1981; Ploen and Courtens, 1986).
According to the present observations, most of the differentiating spermatids disclosed a similar morphogenetic phase. This finding indicates that the spermatids differentiation proceeds simultaneously. The presently illustrated intercellular bridges between the growing spermatids play a role in such phenomenon, since the information (cellular communication) can be transmitted between the spermatids via these bridges (Hiatt and Garetner, 1997). Similar intercellular bridges were described in some other lizard species (Landim and Hoffling, 1977; Al-Hajj et al., 1987; Dehlawi et al., 1990; Dehlawi, 1992; Mubarak, 2004; Al-Dokhi, 2006).
The intercellular cytoplasmic bridges is a sequence of incomplete meiotic division due to a modified cytokinesis (Weiss and Greep, 1977; Fawcett, 1991). Finally, the spermatids are connected together to form a syncytium and the spermatids communicate through the bridges and thus their activities are synchronized (Hiatt and Gartner, 1997). Additionally, some ectoplasmic specialization such as dense plates between Sertoli cells share to establish a synchronous differentiation of spermatids (Junqueira and Carreiro, 1980).
The noticed electron dense material between the acrosomal vesicle membrane and nuclear membrane, represents the subacrosomal material which latter develops into the rod shaped structure, the perforatorium, characteristic for reptiles (Del Conte, 1976; Lin and Jones, 1993).
The currently illustrated acrosomal vesicle deformities were possibly developed as a result of motility of the spermatid cytoplasmic contents. The currently described morphogenetic events are involved in the Golgi and cap phases of spermiogenesis (Hiatt and Gartener, 1997) and considered essential morphological transformations during acrosome development (Sprando and Russel, 1988; Lin et al., 1997).
The present data undoubtedly contribute in establishing the basic knowledge necessary for the researches concerned with phylogenesis in reptilian species.
ACKNOWLEDGMENT
This study was supported by a grant from Research Center-College of Science, King Saud University, Project No. (Zoo/2008/65). We gratefully acknowledge this support.
REFERENCES
- Al-Dokhi, O.A., 2004. Electron microscopic study of sperm tail differentiation in the lizard Bunopus tuberculatus (Squamata, Reptilia). Saudi J. Biol. Sci., 11: 111-117.
Direct Link - Al-Dokhi, O.A., 2006. Ultrastructure of sperm head differentiation in the lizard, Acanthodactylus boskinus (Squamata, Reptilia). Int. J. Zool. Res., 2: 60-72.
CrossRefDirect Link - Da Cruz-Landim C. and M. da Cruz-Hoffling, 1977. Electron microscope study of the lizard spermiogenesis in Tropidurus torquatus (Lacertilia). Caryologia, 30: 151-162.
Direct Link - Del Conte, E., 1976. The subacrosomal granule and its evolution during spermiogenesis in a lizard. Cell Tiss. Res., 171: 483-498.
CrossRef - Ferreira, A. and H. Dolder, 2002. Ultrastructural analysis of spermiogenesis in Iguana iguana (reptilia sauria iguanidae). Eur. J. Morphol., 40: 89-99.
Direct Link - Ferreira, A. and H. Dolder, 2003. Cytochemical study of spermiogenesis and mature spermatozoa in the Tropidurus itambere (reptilia squamata). Acta Histoch., 105: 339-352.
CrossRef - Jamieson, B.G.M., 1995. The Ultrastructure of Spermatozoa of the Squamata (Reptilia) with Phylogenetic Considerations. In: Advances in Spermatozoal Phylogeny and Taxonomy, Jamieson, B.G.M., J. Ausio and J.L. Justine (Eds.). Memooires du Museum National dHitoire Naturelle, Paris, France, pp: 359-383.
- Lin, M. and R.C. Jones, 1993. Spermiogenesis and spermiation in japanese quail (Coturnix coturnix japanonica). J. Anat., 183: 525-535.
PubMed - Lin, M. and R.C. Jones, 2000. Spermiogenesis and spermiation in a monotreme mammal, the platypus, Ornithorhynchus anatinus. J. Anat., 196: 217-232.
PubMed - Lin. M., A. Harman and J.C. Rodger, 1997. Spermiogenesis and spermiation in a marsupial, the tammar wallaby (Macropus eugenii). J. Anat., 190: 377-395.
PubMed - Ploen, L. and J. Courtens, 1986. Comparative aspects of mammalian spermiogenesis. Scann. Electron Mic., 2: 639-652.
PubMed - Mubarak, M., 2004. Ultrastructure of sperm head and its differentiation in the lizard, Bunopus tuberculatus (Squamata, Reptilia). Saudi J. Biol. Sci., 11: 143-151.
Direct Link - Sprando, R.L. and L.D. Russel, 1988. Spermiogenesis in the red-ear turtle (Pseudomys, Scripta) and the domestic fowl (Gallus domesticus): A study of cytoplasmic events including cell volume changes and cytoplasmic elimination. J. Morphol., 198: 95-118.
PubMedDirect Link - Teixeira, R.D., G.H.C. Vieira, G.R. Colli and S.N. Bjo, 1999. Ultrastructural study of spermatozoa of the neotropical lizards Tropidurus semitaeniatus and Tropidurus torquatus (Squamata, tropiduridae). Tissue Cell, 31: 308-317.
CrossRef - Vieira, G.H.C., G.R. Coli and N. Bao, 2004. The ultrastructure of the spermatozoon of the lizard Iguana iguana (Reptilia, Squamata, Iguanidae) and the variability of sperm morphology among. J. Anatomy, 204: 451-464.
Direct Link