
S. Dasari
Department of Biology, University of Kentucky, Lexington, KY 40506-0225, United States of America
L. Wang
Department of Biology, University of Kentucky, Lexington, KY 40506-0225, United States of America
D.A. Harrison
Department of Biology, University of Kentucky, Lexington, KY 40506-0225, United States of America
R.L. Cooper
Department of Biology, University of Kentucky, Lexington, KY 40506-0225, United States of America
International Journal of Zoological Research
Year: 2009 | Volume: 5 | Issue: 3 | Page No.: 101-114







ABSTRACT
The misexpression as well as a knock down of the 5-HT2dro receptor produces slower body movements in larvae and altered development. When 1st instar larvae are raised with altered expression levels a high degree of death occurred. Exposure of the CNS to 5-HT in control larva increases motor unit excitability; however, when the 5-HT2dro expression is decreased the relative sensitivity to exogenously applied 5-HT is enhanced. This is likely a function of reduced basal CNS activity in this line. No change was observed for the strain with an over-expression compared to controls. Evoked sensory-CNS-motor circuits as well as spontaneous motor neuronal activity are also reduced. Like CNS activity, Heart Rate (HR) in larva is sensitive to 5-HT. When the 1st instar to early 3rd instar were chronically reduced or misexpressed in the 5-HT receptor no alteration to 5-HT sensitivity on HR occurred, although the initial HR was lower in both strains as compared to wild type. Thus, a normal expression of the 5-HT2dro is required for development and CNS responsiveness to 5-HT, but this receptor subtype might not function in acute responsiveness of the heart to 5-HT, although the receptor has some effect on basal heart rate.
PDF Abstract XML References Citation
How to cite this article
S. Dasari, L. Wang, D.A. Harrison and R.L. Cooper, 2009. Reduced and Misexpression of 5-HT2 Receptors Alters Development, Behavior and CNS Activity in Drosophila melanogaster. International Journal of Zoological Research, 5: 101-114.
DOI: 10.3923/ijzr.2009.101.114
URL: https://scialert.net/abstract/?doi=ijzr.2009.101.114
DOI: 10.3923/ijzr.2009.101.114
URL: https://scialert.net/abstract/?doi=ijzr.2009.101.114
INTRODUCTION
Serotonin (5-HT) is a major neurotransmitter and neuromodulator in both vertebrates and invertebrates. It has been shown to have a role in development and behaviors of various vertebrates and invertebrates (Lucki, 1998; Monastirioti, 1999; Osborne, 1996; Whitaker-Azmitia, 2001). Recently, it was demonstrated that 5-HT increases activity in a sensory-CNS-motor circuit in larvae (Dasari and Cooper, 2004; Dasari et al., 2007). When reducing the production of 5-HT, by pharmacological means through feeding p-CPA to larva, there is a delay in the rate of development and slowed motor behaviors (body wall contractions and eating) (Dasari et al., 2007). Likewise, exposure to MDMA (ecstasy), a drug of abuse, which impacts the serotonergic system acutely as well as chronically, also alters larval development and bodily movements (Dasari et al., 2007). Thus, altering the production or the level of 5-HT has broad ranged effects on larval Drosophila, however, to understand the mechanisms by which these alterations occur one needs to address the effectors of 5-HT which relay the signals.
The 5-HT acts through multiple receptor subtypes. There are 14 known 5-HT receptors in vertebrates (Barnes and Sharp, 1999) whereas four are known to occur in the Drosophila genome (Tierney, 2001). Hence, Drosophila melanogaster is an attractive model organism to study the role of 5-HT mediated by these receptors. The four known receptors are: 5-HT7do, 5-HT1Adro, 5-HT1Bdro, and 5-HT2dro (Colas et al., 1994; Saudou et al., 1992; Tierney, 2001; Witz et al., 1990).
The 5-HT2dro receptor subtype is of particular interest because in humans the 5-HT2 receptor is known to be associated with many diseases such as schizophrenia, depression and anxiety (Leonard, 1994; Fuller, 1991). Drosophila offers some advantages in the ability to address the developmental and mechanistic understanding of neuromodulators, particularly 5-HT because this model organism is being increasingly used to address physiologically related as well as the genetic diseases that afflict humans (Kendler and Greenspan, 2006).
Colas et al. (1999a, b) showed that 5-HT2dro is essential during embryo genesis and that it is the major receptor subtype in 3rd instar larvae (Colas et al., 1994). Thus, we hypothesized that there would be behavioral and physiological alterations in larva if the receptor is over-or under-expressed. One approach to determine the role of 5-HT2dro is to selectively knock down its expression. In this study, a heat shock inducible antisense transgenic line, Y32 was designed to specifically reduce the functional expression of the 5-HT2dro receptor (Colas et al., 1999b). Another heat shock inducible transgenic strain, HM2, misexpresses 5-HT2dro in all tissues. The Y32 transgene uses direct expression of an antisense fragment of 5-HT2dro driven by hsp70 gene promoter. The HM2 strain carries a hsp70-driven GAL4 transgene plus a UAS-full-length 5HT2dro cDNA (Brand and Perrimon, 1993). It is expected that both transgenic lines would have low basal expression at low temperature (18°C or less), but would have chronic temperature-dependent expression at elevated, but sub-heat shock temperatures (below 37°C) (Brand and Perrimon, 1994). To determine the effects of reduced and misexpressed 5-HT2dro on behavior and on CNS receptivity to 5-HT, studies at low and high temperatures were compared in wild type controls and the two strains (HM2 and Y32).
Johnson et al. (1997) have shown heart rate to be modulated by 5-HT in Drosophila. We have also shown direct effects of various doses of 5-HT on the function of the Drosophila larval heart (Dasari and Cooper, 2006). We measured heart rate in the fly strains to investigate if the 5-HT2dro receptor regulates its function in larvae.
These studies are significant because they demonstrated that the 5-HT2dro expression is important in overall development, normal behavior and heart function. It is feasible now to address mechanisms of how these functions are altered. This will help to determine if the same mechanisms are translated to mammals that exhibit similar consequences with perturbed serotoninergic systems.
MATERIALS AND METHODS
Stock and Staging of Larvae
The common wild-type laboratory strain of Drosophila melanogaster, Canton S, was used in these studies as controls. HM2 and Y32 are used as experimental lines. The HM2 strain contains both an hsp70 promoter-GAL4 transgene and a UASGAL4-5-HT2dro transgene, permitting misexpression of the receptor in a temperature-dependent manner. Y32 permits knock-down by directing a 5-HT2dro antisense RNA under direct control of the hsp70 promoter (Colas et al., 1999a,b). Both lines were gifts from Dr. L. Maroteaux, INSERM U839, Institut du Fer a Moulin, Paris, France. The methods used to stage fly larvae have been described earlier (Campos-Ortega and Hartenstein, 1985; Li and Cooper, 2001). All animals were maintained in vials partially filled with a cornmeal-agar-dextrose-yeast medium. All animals were kept on a 12:12 light-dark cycle. The fly stocks were maintained and experiments were carried out from 2005-2008, in Department of Biology, University of Kentucky, Lexington, KY., USA.
Behavioral Assays
Early 3rd instar larvae were used for behavioral assays. Feeding and locomotor behaviors were assessed as described in Neckameyer (1996) and Li et al. (2001). In brief, single animals were placed on a 2% agar surface and the number of body wall contractions was counted for 1 min, after which an animal was placed in a 2% yeast solution overlaid on an agar plate (just covering the larvae allowing the spiracles to reach out of the solution). In this condition, Drosophila larvae immediately start feeding, initiating a pattern of repetitive mouth hook movements. The number of full mouth hook contractions in 1 min was counted (Sewell et al., 1975). Early 3rd instars were given a 4 h heat pulse at 33°C and then the locomotor and feeding behaviors were measured at room temperature (21°C) within 5 min of taking them out of 33°C. Each larva was taken from the incubator and placed on a test plate. This allowed the larvae to become familiar with the test plate for a minute. Then body wall and mouth hook movements were measured.
Dissection and Electrophysiological Recordings
Wandering 3rd instar larvae were dissected as described earlier (Cooper et al., 1995). In brief, larvae were dissected ventrally removing heart and viscera which left a filleted larvae containing only a body wall, body wall muscles and the neural circuitry for the sensory, CNS and body wall (i.e., skeletal) motor units. The HL3 saline was prepared in the lab from component reagents (Sigma) and contained: 1.0 mM CaCl2•2H2O, 20 mM MgCl2, 70 mM NaCl, 5 mM KCl, 10 mM NaHCO3, 5 mM trehalose, 115 mM sucrose and 5 mM BES (N,N-bis[2-Hydoxyethyl]-2-aminoethanesulfonic acid) (Stewart et al., 1994). The recording arrangement was essentially the same as priviously described (Neckameyer and Cooper, 1998; Stewart et al., 1994). Intracellular recordings in muscles were made with 3 M KCl-filled microelectrodes (3-60 MΩ). The amplitudes of the excitatory postsynaptic potentials (EPSP) elicited by Is and Ib motor nerve terminals in the various segments of muscles m6 were monitored. Intracellular responses were recorded with a 1xLU head stage and an Axoclamp 2A amplifier.
Sensory-CNS-Motor Circuit
To induce a sensory-CNS-motor circuit the tail segmental nerves were cut and stimulated using the suction electrode. Detailed procedures have been outlined priviously (Dasari and Cooper, 2004). Stimuli were given in short bursts at a frequency of 40 Hz (10 stimuli at 40 Hz). The sensory stimulation leads to activation of interneurons and motor neurons, thus inducing a response in muscle 6 which is monitored. A change in the evoked response is used to examine the effects of 5-HT on the circuit.
In cases when it was desirable to measure the effect of 5-HT on intrinsic activity all the segmental nerves were left intact. The intrinsic CNS activity probably is induced by the fact the animal has been cut down the dorsal midline and stretched out on a dish by pinning four corners of the animal down. Thus, there is a background sensory drive. The burst frequency and frequency within a burst were measured for experimental comparisons. All experiments were performed at room temperature (20-22°C).
Heart Rate (HR) Measures
The same microscopic method as for behavioral movements was used to record HR but with the exception of a 2 x base objective to obtain a higher resolution of the heart and trachea. Early 3rd instar larvae were dissected ventrally and pinned on 4 corners. The heart and trachea are exposed in these semi-intact preparations. The movements of the trachea or heart were used for direct counts (Dasari and Cooper, 2006). Effects of 5-HT were observed in CS, HM2 and Y32 larvae, which were grown at 18, 21 and 31-32°C.
Developmental Assays
Eggs were collected after a 15 min prepulse (i.e., clearing the oviduct of stored eggs) for 2 h. The eggs were incubated at 21°C until hatching. At which time 18, 21 and 31-32°C were used to determine effects of the anti-sense activation on development. To control if room temperature might be a temperature at which a low expression level of the heat shock promotor might be active, 18°C was used as well as room temperature (21°C).
First instars (about 15) were transferred to vials containing food and placed at the given temperatures. The food was made with a standard corn meal diet (Neckameyer, 1996; Li and Cooper, 2001). When wandering 3rd was seen, the vials were checked every 4 h and each individual pupa was marked on the walls of the tube to record the time to pupation. This was done throughout the day and night. The time to pupation and the time spent as a pupa, until eclosion, were indices for development.
Quantification of Transgene Expression by Real Time RT-PCR
The heat-dependent knock-down of 5-HT2dro receptor expression by Y32 antisense has been previously demonstrated (Colas et al., 1999a, b). To assess the degree of misexpression directed by the HM2 line at high temperature, real time RT-PCR was performed. Total RNAs of heat-shocked (6 h at 37°C) HM2 third instar larvae and control wild type third instar larvae were isolated with Invitrogen TRIzol® solution. Obtained total RNAs were kept in RNAsecureTM suspension solution (Ambion Inc) and treated with DNase for removing genomic DNA (TURBO DNA-freeTM, Ambion Inc). One microgram of DNase treated total RNA was used for cDNA synthesis with iScriptTMcDNA synthesis kit (BIO-RAD). Twenty microliter of cDNA was obtained for each sample and then diluted in 20 μL DEPC H2O to make a 40 μL cDNA pool. One microliter of diluted cDNA was subjected to real time PCR amplification analysis using iQTM SYBR Green Supermix (BIO-RAD) on a BIO-RAD iCycler. Amplification was performed with two experimental primer sets for 5-HT2dro and one control for the housekeeping gene Rp49: 5-HT2dro primer pair 1: catgggcgcgataccagcttt, ccaaactgatgaagcacatgtgca; 5-HT2dro primer pair 2: gcaaaaggtgaagccgacgg, ccttctgctccgtagcaacg; rp 49 (control for normalization): cagcatacaggcccagatcgt, cttactcgttctcttgagaacgcag. Polymerase chain reaction cycles were set up as follows: Cycle 1(1x) 94°C for 01:30; Cycle 2 (40x): 94°C for 00:15, 60°C for 01:00, 72°C for 01:00; Cycle 3 (1x): 95°C for 01:00; Cycle 4 (80x): 60°C for 00:10 and then increase temperature by 0.5°C after the second cycle for melt curve data collection and analysis; Cycle 5 (1x): hold at 4°C. The data were analyzed according to the 2-ÄÄCT method (Livak and Schmittgen, 2001; Pfaffl, 2001; Schmittgen and Livak, 2008). As compared with wild type larvae, the HM2 larvae at high temperature showed a 504±26 fold increase in 5-HT2dro expression.
Statistical Analysis
The ANOVA test is used where appropriate. For developmental studies we use a Kolmogorov-Smirnov (KS) test to determine if the distribution of pupation and eclosion times of the 5-HT2dro transgenics differ from the controls (Cesani et al., 2006; Mentre and Escolano, 2006).
RESULTS
Behavior-Mouth Hook and Body Wall Movements
Locomotion (body wall) and feeding (mouth hook) movements were measured for CS, HM2 and Y32 strains that were raised from 1st instar to 3rd instar under low-expressing conditions (21°C) and higher-expressing conditions (33°C) to induce the heat shock antisense or over production of the 5-HT2dro receptor. There was no significant difference seen among the three lines in mouth hook movements when larvae were raised at room temperature (21°C) or high temperature (33°C) (ANOVA, N >10, Fig. 1b). But, the HM2 strain showed reduced body wall movements compared to CS at both the temperatures (Fig. 1a, ANOVA, p<0.01, N = 25). Similarly, Y32 also showed slower body wall movements than CS at both the temperatures (Fig. 1a, ANOVA, p<0.05, N = 25).
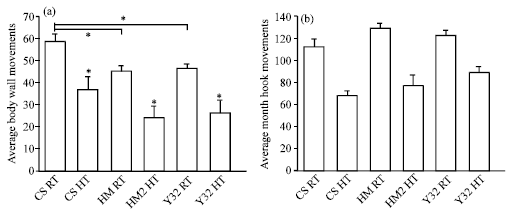 | |
| Fig. 1: | Body wall and mouth hook movements for CS, HM2 and Y32 in 3rd instar larva. (a) Larvae are grown at room temperature (RT, 21°C) or high temperature (HT, 32°C) from 1st to 3rd instar stage and body wall movements are counted in early wandering 3rd instar at room temperature. The HM2 strain showed reduced body wall movements compared to CS (ANOVA, p<0.01, N = 25). Y32 also showed slower body wall movements than CS (ANOVA, p<0.05, N = 25). There is a significant difference in each of the three strains with the heat shock pulse as compared to 21°C (* p<0.05). (b) Mouth hook movements in a 3rd instar larvae for the same treatment paradigm. No significant difference is present among the 3 groups at RT or at HT (N >10, ANOVA) |
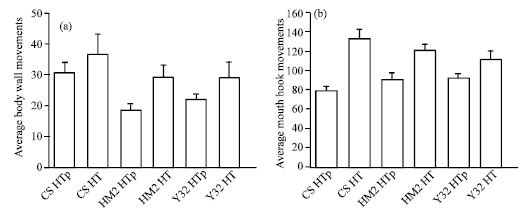 | |
| Fig. 2: | Larvae are grown at 21°C and each group was given a heat pulse at 33°C for 4 h. The average (a) body wall movements and (b) mouth hook movements are shown. The larva from the 4 h heat pulse and assayed at 21°C is indicated by p (pulse) and the other heat shock group was monitored at 33°C following the 4 h heat shock. No significant difference is observed among 3 groups (N >10, ANOVA) for body wall or mouth hook movements |
To examine effect a shorter heat pulse would have on acute behaviors, the early 3rd instars were examined after a 4 h heat shock at 33°C and monitored at room temperature (21°C). Also, to control for the temperature variation by taking the larva out of the incubator to conduct the behavioral assays, the feeding and locomotor behaviors were measured at 33°C after a 4 h heat pulse at the same temperature. Again no significant difference in the mouth hook movements occurred (Fig. 2b, N = 10) but this time no significant difference for body wall movements occurred among the CS, HM2 and Y32 with heat shock (Fig. 2a, N = 10). There is a significant difference in each of the three strains with the heat shock pulse as compared to 21°C. These results have important implications in the manner in which one conducts experimental paradigms involving heat shocking and behavioral assays at a different temperature.
Larval Development
Developmental assays were carried out at 18°C, room temperature (21-22°C) and 31-32°C for CS, HM2 and Y32. Time was assessed for 50% of population to reach pupation or eclosion. The time in which eggs were collected is defined as zero time.
At low temperature, where the 5-HT2dro transgenes are not being expressed, we did not anticipate to see any major difference in the development patterns among the strains. But a small variation was observed between the 3 groups for time to pupation (Fig. 3). This small variation continued for the time from pupation to eclosion. For calculating the amount of time taken for eclosion for each pupa, the time point at which pupa formed was set to 0 (zero) and number of hrs is calculated to eclosion.
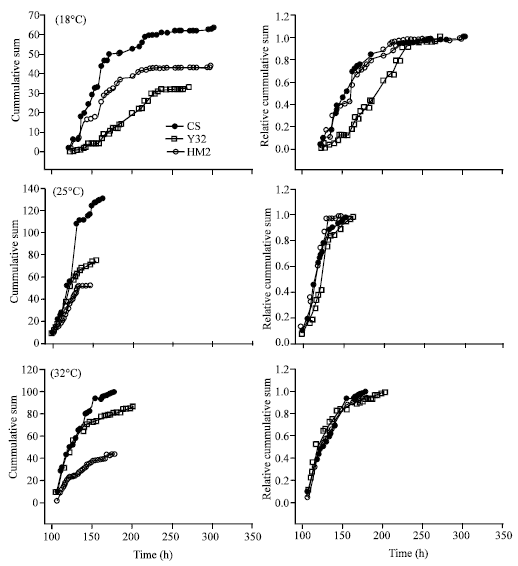 | |
| Fig. 3: | Development curves from egg to pupa formation in CS, HM2 and Y32 strains grown at three different temperatures (18, 21 and 31-32°C). Cumulative sum (left side) and relative cumulative sum (right side) plots are shown for each. There are no significant differences among the groups within a particular experimental temperature paradigm |
There are insignificant differences for CS, HM2 and Y32 in both time to pupation and eclosion. To statistically test for a difference in the distributions a Kolmogrov-Smirnov (K-S) test was performed. D and P values for pupation among all groups were not significant. Similarly eclosion at room temperature (21°C) is not significant. Also, for eclosion time a K-S test was used to assess for statistically significant differences in the distributions. No significant difference was seen for pupation in any groups. Chronic expression of the 5-HT2dro transgenes at 31-32°C from 1st instar stage also had no pronounced effects. Examining the effects on eclosion proved to be futile since many of Y32 and HM2 pupa died. We conclude that either reduction or misexpression of 5-HT2dro is detrimental to the health of the developing pupa.
Spontaneous Activity
The intrinsic motor activity, driven by spontaneous CNS events, was measured from m6 or m7 muscles as both these muscles receive the same innervation by two motor neurons (Is and Ib). The spontaneous motor neuron activity is present when the larva is dissected, stretched and pinned. This procedure likely keeps sensory neurons active which drives the motor neurons. This could account for the variation in activity patterns observed from preparation to preparation. Two sets of experimental paradigms were used here; (1) larvae grown at room temperature and neural activity assessed at room temperature and (2) larvae grown at 31-32°C and intrinsic activity was measured in saline at room temperature. The sensitivity to 100 nM 5-HT was examined for all groups in these two conditions. Intrinsic activity was recorded for 2 min in saline and then another 6-10 min in the presence of 5-HT. For quantifying, the average frequency per second, all the peaks within each burst was measured. Also, the time for each burst was recorded. The average frequency per second in saline and in presence of 5-HT was used to calculate the percent difference in activity from saline. At room temperature (21°C) 5-HT (100 nM) sensitivity of the CS increased by approximately 15%, whereas HM2 and Y32 showed a decreased responsiveness (Fig. 4a). There is no statistical significance seen among CS, HM2 and Y32 (N = 5, ANOVA, Fig. 4a). However, the CS and HM2 larvae grown at 31-32°C and exposed to 5-HT (100 nM) showed the opposite effect to Y32 (Fig. 4b). CS and HM2 showed a decreased response whereas Y32 showed a significant increase in activity from saline (N = 5, ANOVA, p<0.013 and Tukey post hoc test, p<0.05, Fig. 4b). Y32 did not have as much activity in saline initially when compared to that of CS and HM2. The basal activity for Y32 that were grown at 31°C was 8.75±2.41, where as for CS and HM2, the basal activity was higher 17.17±2.24 and 27.42±2.14.
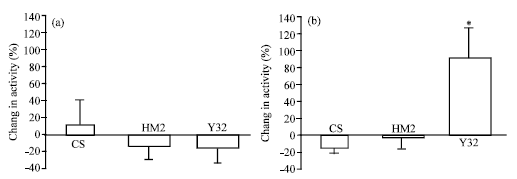 | |
| Fig. 4: | The percent change in frequency per second (activity) from saline to 5-HT (100 nM) is (a) measured in CS, HM2 and Y32 larvae grown at permissive temperature and (b) non-permissive temperature. (a) At room temperature no significant difference in sensitivity for 5-HT (100 nM) was observed among the 3 groups (N = 5, ANOVA). (b) CS and HM2 larvae that were grown at high temperature showed a decrease in activity in presence of 5-HT (100 nM) but not significantly (N = 5, ANOVA). However, Y32 showed a significant increase in activity (N = 5, ANOVA, p<0.01) |
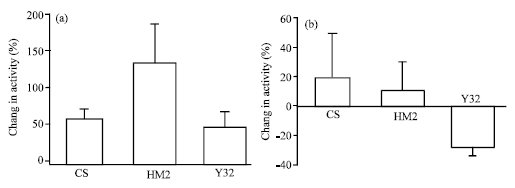 | |
| Fig. 5: | Sensory-CNS-motor circuit. (a) A percent difference in the activity of motor units to muscle 6 before and after application of 5HT (100 nM) in larvae grown at permissive temperature. All the 3 groups showed an increase in activity with no statistical difference seen (N=5, ANOVA). (b) A percent difference in activity exposed to 5-HT (100 nM) for larvae that were grown at non-permissive temperature. CS and Y32 showed an increase in activity but Y32 showed a decrease in activity. No statistical difference is present among the 3 groups (N=5, ANOVA) |
The increased activity of Y32 seen in presence of 5HT could be due to a low initial rate of activity.
Sensory-CNS-Motor Circuits
The sensitivity of central circuits to 5-HT in mutant lines was examined by stimulating sensory nerves and monitoring motor units before and during exposure to the compounds. CS showed an increase in motor activity in presence of 5-HT (100 nM) at room temperature. Similarly HM2 and Y32 showed an increase in activity at room temperature in response to 5-HT (100 nM). No statistical difference was seen among the 3 groups (N = 5, ANOVA, p<0.15, Fig. 5a). The basal activity seen in saline for CS and HM2 was similar (10.65±2.15 and 10.77±1.90, respectively), but Y32 showed a slightly higher basal activity with an average activity of 18.00±2.73. Possibly at room temperature (21°C) a low level of receptor misexpression occurs in the HM2 line which produces an increase in responsiveness but with a large degree of variability. However, under heat shock an excessive expression of the receptor somehow reduces the sensitivity to exogenous application of 5-HT. For both 5-HT2dro knock-down and misexpression, sensitivity of CNS-circuit to 5-HT (100 nM) changed. The average basal activity for both HM2 (4.63±1.08) and Y32 (6.26±1.02) was seen to be very small as compared to CS (10.01±2.98). The CS and HM2 that were grown at high temperature (31-32°C) showed a very small increase in motor activity. On the other hand, Y32 grown at high temperature showed a decrease in activity. Y32 larvae grown at 31-32°C have a lower activity in saline and there was a fast run down in the activity. The frequency of stimulation had to be increased in some preparations to 60 Hz from the 40 Hz, as used for all the other preparations, in order to evoke motor nerve activity. Again no statistical significance is present among the 3 groups (N = 5, ANOVA, p<0.23, Fig. 5b).
Heart Rate (HR)
5-HT is known to modulate heart rate in Drosophila larvae (Dasari and Cooper, 2006; Johnstone and Cooper, 2006). Hence, we measured the sensitivity of the heart to 5-HT in the HM2 and Y32 larvae with altered expression in the 5-HT2dro receptor to determine if this receptor subtype had any role on HR. To quantify the responsiveness a percent difference is calculated for each preparation from saline to 5-HT and the average percent change for the group is compared (Dasari and Cooper, 2006). CS, HM2 and Y32 larvae grown at room temperature (21°C) did not show any significant difference in sensitivity to 5-HT. All 3 groups showed an increase in the HR in presence of 5-HT, though HM2 showed a little higher increase, but not statistically significant (N = 10, ANOVA, p<0.1), than both CS and Y32 (Fig. 6a).
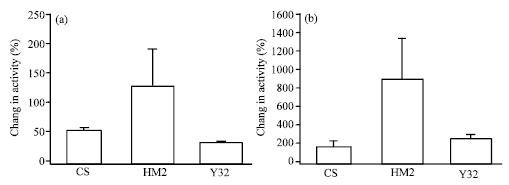 | |
| Fig. 6: | The percent change in heart rate upon exposure to 5-HT (100 nM). (a) CS, HM2 and Y32 larvae that were grown at room temperature showed an increase in heart rate with no statistical difference (N = 10, ANOVA). (b) CS, HM2 and Y32 larvae that were grown at high temperature showed an increase in heart rate with no statistical difference (N=10, ANOVA). But the initial heart rate in HM2 and Y32 are lower giving an abnormal percentage change in BPM (BPM-beats min-1) |
In a different set of experiments, larvae that were grown at 31-32°C were taken out one at a time right before the experiment and heart rate counted at room temperature (21°C) for one min in saline and another 3 min in presence of 5-HT. The total time taken for each preparation is about 5 min. The average initial HR in saline was low for all three strains CS (46±5), HM2 (20.88±4.62) and Y32 (25.3±3.55). The CS larvae showed a small variation in the initial HR as compared to that of room temperature (21°C). Y32 had a very low HR with some of preparations not beating at all. In all cases, when the 5-HT containing saline was introduced HR increased. Because of the low initial HR, a percent difference gives a large difference as compared to preparations starting at a higher HR with less of a change. No statistical difference occurred among the 3 groups (N = 10, ANOVA, p<0.41, Fig. 6b). The HM2 shows a greater mean, as well as a large degree of variability, in response to 5-HT for larvae raised at room temperature (21°C) as well as higher temperature. This would not be surprising because amplified expression of the binary expression system creates high sensitivity and there is likely some basal expression at 21°C.
DISCUSSION
In this study, two Drosophila lines were used to alter the expression of the 5-HT2dro receptor. The knock down in Y32 is accomplished by anti-sense 5-HT2dro under a heat shock promoter. The misexpressing line, HM2, is ubiquitously expressing 5-HT2dro in all cells at elevated temperatures. These two extremes in the presence of the 5-HT2dro receptor were used to assess developmental, behavioral and physiological significance of this receptor subtype. The development of Y32 and HM2 at room temperature (21°C), 18°C and chronic heat induction (31-32°C) were not significantly different from controls. However, at 31-32°C most of the CS formed pupa but a lower percentage of Y32 and HM2 larvae pupated. So, some larvae are dying in these mutant strains, but pupal development was the real limiting factor in survival. None of the Y32 or HM2 strains eclosed at these higher temperatures. However, no significant difference is present in simple locomotor movements (body wall and mouth hook movements) in wandering larvae that are grown at 33°C. On the other hand, the physiological responses of the CNS to 5-HT application in Y32 and HM2 strains grown at 31-32°C had a decreased responsiveness compared to controls. Even though the initial HR was lower in Y32 and HM2 raised at 31-32°C as compared the same strains raised at 21°C, there is no effect on sensitivity to 5-HT in relation to acute cardiac function. Thus, these results show that the 5-HT2dro receptor plays an important role during the metamorphosis from larval to a adult stage but the lack of any effect on the locomotor movements or cardiac function suggest that these functions are not modulated by 5-HT2dro receptor or other receptors are compensating for the misfunction of the 5-HT2dro.
Studies altering synaptic transmission of adult vertebrate motor nerve terminals have provided evidence that established synapses and nerve terminals maintain a high degree of plasticity. Many of the same regulating factors in synaptic maintenance in adult animals are likely utilized during developmental stages, so understanding how synapses develop have ramifications for mature synapses. Sokolowski (1980) showed that the behavioral repertoire of Drosophila larvae requires flexible synaptic inputs. The development in the larval Drosophila CNS is a topic that has not been intensively studied but has been gaining interest in recent years (Bossing and Brand, 2002, 2006). Since, the larval CNS contains substantially fewer neurons ~2,000 (Nassif et al., 2003) as compared to the 100,000 (Iyengar et al., 2006) or more in the adult, the regulatory factors might be easier to assess and quantify in the larval brain. The strains we examined in this study open the field for further investigation into CNS development.
In general, very little research has been conducted on the role of the Drosophila 5-HT receptors in development and CNS function. The 5-HT2dro is 40% homologous over the transmembrane domain of 5-HT2 receptor of mammals. 5-HT2dro is present on 3rd chromosome and right arm. Colas et al. (1999a, b) reported that 5-HT2dro mRNA is expressed during embryogenesis and gastrulation. With the use of these 5-HT2dro mutants it has been shown that the receptor must have some functional role in embroygenesis since ectoderm extension, during embryogenesis is delayed and hence embryonic lethality is observed (Colas et al., 1999a, b). In these studies, the mutation was activated at a 1st instar stage which reduced the number that reached the pupal stage. The ones which did pupate, however did not eclose. Like in other animals, 5-HT is thought to have a major role during development for various tissues. 5-HT is detected in sea-urchins (Buznikov et al., 2005), chicken and frogs during zygotic divisions in which gastrulation and neurulation takes place. Nebigil et al. (2000) had shown that mice deficient in 5-HT2B receptor even have a defective heart.
There are few studies in mammals or other animals that addressed the developmental and behavioral effects of misexpression of 5-HT2 receptors. 5-HT1A receptors are known to be increased in the cortex for some schizophrenic patients (Kasper et al., 2002). In another study with mice, a genetic deletion or over expression of 5-HT1A-receptors impaired learning and memory in certain tests but not others (Bert et al., 2008). This suggests in mice that the 5-HT1A receptor is modulatory on neuronal function. Since, we showed lethality in development, we assume 5-HT2 receptors in larval Drosophila serve a greater role in survival. Oddly enough, over or under expression has been shown to increase 5-HT synthesis and turnover in the postnatal mice (Deng et al., 2007). We did not measure 5-HT levels in these larvae with altered receptors levels; however, such a follow up to this study would be useful for comparative purposes. The Drosophila genome contains only four 5-HT receptor subtypes, so it would be feasible to dissect the role of each one in development and behavior.
Since, 5-HT modulates locomotive and behaviors in insects and that 5-HT containing neurons are known to innverate guts, pharyngeal muscles and ring gland in Drosophila (Valles and White, 1988), one would assume that the knock down of the 5-HT2dro receptor would produce very pronounced effects on eating behaviors. It was surprising that no significant effect on locomotor or feeding behavior were obvious in our studies. The rationale for explaining the lack of an effect is probably that these behaviors are regulated by other 5-HT receptors and not the 5-HT2dro. Recently the 5-HT1 receptor was shown to have a role in sleep behavior in Drosophila (Yuan et al., 2006). Unlike Drosophila, the 5-HT2A in mammals is involved in feeding behavior and 5-HT2C in regulating satiety (Lucki, 1998).
Other neuromodulators that are known to be in the hemolymph and released into the nuropile of the larval CNS, such as octopamine and dopamine are likely to have key roles in shaping the development of the neural circuits (Monastirioti, 1999). Since, there is such a pronounced effect of octopamine on the neural activity (Dasari and Cooper, 2004), one would expect a larva deficit in the production or in the receptors to result in substantial changes in CNS development. As far as we are aware, such studies have yet to be conducted. The effects of octopamine and a related compound, tyramine, have been examined in relation to honey bee behavior and flight (Fussnecker et al., 2006). It is likely that similar developmental cues such as activity and local retrograde signaling factors hold for synapse formation and maintenance in the larval CNS as established in other model preparations, however detailed studies on defined tracts are necessary to substantiate the various possibilities. Combining physiological measures of larval CNS circuits with the anatomical profiles along with behavioral assays will undoubtedly be an area of significant interest in the next few years.
The 5-HT and its receptors have a role in cardiovascular physiology (Nebigil et al., 2001; Hoyer et al., 2002). Loss of Tph1 gene (peripherally expressed TPH) in mice results in abnormal cardiac function (Cote et al., 2003) and knock out of the 5-HT2B receptor in mice results in lethality due to defects in heart development (Nebigil et al., 2000). A follow up on the function in adult hearts in these Drosophila lines with altered 5-HT2 receptors as well as understanding the mechanism for a slower heart rate in the larvae could help shed some light on the heart and aortic defects noted in human and other mammals with over expression of 5-HT1B/1C and 5-HT2A/2BR subtypes (Xu et al., 2002; Fitzgerald et al., 2000; Roy et al., 2000). In present studies, both 5-HT2dro mutant lines have a lower initial heart rate when compared to that of room temperature or controls that were grown at high temperature, but sensitivity to 5-HT is not altered. Probably Drosophila cardiac function is not modulated only by the 5-HT2dro receptor subtype. Detailed pharmacological studies with agonists and antagonists of all four 5-HT receptors of Drosophila would give more insight to this issue.
Clinically in humans, in addition to the role of 5-HT2 receptors have in embryogenesis, the receptors may be important as determinants in migraine, hypertension, heart failure, neurodegenerative diseases and brain maturation disorders such as schizophrenia or autistic behaviors. It is now known that the presence of 5-HT decreased interneuron migration in mice and 5-HT6 activation mimicked the effect, whereas blockade of 5-HT6 prevented migration (Riccio et al., 2009). The Present study is significant since it provides pertinent information, which addresses the degree in synaptic performance at the release sites over developmental stages in a model system of Drosophila when a defined receptor subtype is altered in expression. The findings can be correlated to the underlying factors of the serotoninergic system during development which provides a frame work for future studies in knocking down the other three 5-HT receptor subtypes or even sensitivity to other neuromodulators such as dopamine or octopamine. Thus, there is a premise that understanding the fundamental basics of synaptic transmission in this model system will be directly relevant to all neural systems, including humans.
ACKNOWLEDGMENTS
Funding was provided in part by NSF grants NSF-IBN-0131459 (RLC) and a G. Ribble Fellowship in the School of Biological Sciences at the University of Kentucky (SD). We appreciate the help in the developmental studies by Ms. Blaire Cullman-Clark (undergraduate student at Univ. KY).
REFERENCES
- Barnes, N.M. and T. Sharp, 1999. A review of central 5-HT receptors and their function. Neuropharmacology, 38: 1083-1152.
PubMedDirect Link - Bert, B., H. Fink, J. Rothe, J. Walstab and H. Bonisch, 2008. Learning and memory in 5-HT(1A)-receptor mutant mice. Behav. Brain Res., 195: 78-85.
CrossRefDirect Link - Bossing, T. and A.H. Brand, 2002. Dephrin, a transmembrane ephrin with a unique structure, prevents interneuronal axons from exiting the Drosophila embryonic CNS. Development, 129: 4205-42018.
Direct Link - Bossing, T. and A.H. Brand, 2006. Determination of cell fate along the anteroposterior axis of the Drosophila ventral midline. Development, 133: 1001-1012.
PubMed - Brand, A.H. and N. Perrimon, 1993. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development, 118: 401-415.
PubMedDirect Link - Brand, A.H. and N. Perrimon, 1994. Raf acts downstream of the EGF receptor to determine dorsoventral polarity during Drosophila oogenesis. Genes Dev., 8: 629-639.
PubMed - Cesani, M.F., A.B. Orden, E.E. Oyhenart, M. Zucchi, M.C. Mune and H.M. Pucciarelli, 2006. Growth of functional cranial components in rats submitted to intergenerational undernutrition. J. Anat., 209: 137-147.
PubMed - Colas, J.F., J.M. Launay and L. Maroteaux, 1999. Maternal and zygotic control of serotonin biosynthesis are both necessary for Drosophila germband extension. Mech. Dev., 87: 67-76.
PubMed - Colas, J.F., J.M. Launay, J.L. Vonesch, P. Hickel and L. Maroteaux, 1999. Serotonin synchronises convergent extension of ectoderm with morphogenetic gastrulation movements in Drosophila. Mech. Dev., 87: 77-91.
PubMed - Cooper, R.L., B.A. Stewart, J.M. Wojtowicz, S. Wang and H.L. Atwood, 1995. Quantal measurement and analysis methods compared for crayfish and Drosophila neuromuscular junctions and rat hippocampus. J. Neurosci. Methods, 61: 67-78.
CrossRefDirect Link - Cote, F., E. Thevenot, C. Fligny, Y. Fromes and M. Darmon et al., 2003. Disruption of the nonneuronal tph1 gene demonstrates the importance of peripheral serotonin in cardiac function. PNAS., 100: 13525-13530.
PubMed - Dasari, S. and R.L. Cooper, 2004. Modulation of sensory-CNS-motor circuits by serotonin, octopamine and dopamine in semi-intact Drosophila larva. Neurosci. Res., 48: 221-227.
CrossRefPubMedDirect Link - Dasari, S. and R.L. Cooper, 2006. Direct influence of serotonin on the larval heart of Drosophila melanogaster. J. Comp. Physiol. B, 176: 349-357.
CrossRefDirect Link - Dasari, S., K. Viele, A.C. Turner and R.L. Cooper, 2007. Influence of p-CPA and MDMA (ecatasy) on physiology, development and behavior in Drosophila melanogaster. Euro. J. Neurosci., 26: 424-438.
PubMed - Deng, D.R., S. Djalali, M. Holtje, G. Grosse and T. Stroh et al., 2007. Embryonic and postnatal development of the serotonergic raphe system and its target regions in 5-HT1A receptor deletion or overexpressing mouse mutants. Neuroscience, 147: 388-402.
PubMed - Fitzgerald, L.W., T.C. Burn, B.S. Brown, J.P. Patterson and M.H. Corjay et al., 2000. Possible role of valvular serotonin 5-HT(2B) receptors in the cardiopathy associated with fenfluramine. Mol. Pharmacol., 57: 75-81.
PubMed - Fuller, R.W., 1991. Role of serotonin in therapy of depression and related disorders. J. Clin. Psych., 52: 52-57.
PubMed - Fussnecker, B.L., B.H. Smith and J.A. Mustard, 2006. Octopamine and tyramine influence the behavioral profile of locomotor activity in the honey bee (Apis mellifera). J. Insect Physiol., 52: 1083-1092.
CrossRefPubMedDirect Link - Hoyer, D., J.P. Hannon and G.R. Martin, 2002. Molecular, pharmacological and functional diversity of 5-HT receptors. Pharm. Biochem. Behav., 71: 533-554.
PubMed - Iyengar, B.G., C.J. Chou, A. Sharma and H.L. Atwood, 2006. Modular neuropile organization in the Drosophila larval brain facilitates identification and mapping of central neurons. J. Comp. Neurol., 499: 583-602.
PubMedDirect Link - Johnson, E., J. Ringo and H. Dowse, 1997. Modulation of Drosophila heartbeat by neurotransmitters. J. Comp. Physiol. B, 167: 89-97.
CrossRefPubMedDirect Link - Johnstone, A.F.M. and R.L. Cooper, 2006. Direct innervation of the Drosophila melanogaster larval aorta. Brain Res., 1083: 159-163.
CrossRefDirect Link - Kasper, S., J. Tauscher, M. Willeit, M. Stamenkovic and A. Neumeister et al., 2002. Receptor and transporter imaging studies in schizophrenia, depression, bulimia and Tourette's disorder�implications for psychopharmacology. World J. Biol. Psychiatry, 3: 133-146.
PubMedDirect Link - Kendler, K.S. and R.J. Greenspan, 2006. The nature of genetic influences on behavior: Lessons from simpler organisms. Amer. J. Psych., 163: 1683-1694.
PubMed - Leonard, B.E., 1994. Serotonin receptors--where are they going?. Internat. Clinic Psychopharmcol., 1: 7-17.
PubMed - Li, H. and R.L. Cooper, 2001. Effects of the ecdysoneless mutant on synaptic efficacy and structure at the neuromuscular junction in Drosophila larvae during normal and prolonged development. Neuroscience, 106: 193-200.
PubMed - Li, H., D. Harrison, G. Jones, D. Jones and R.L. Cooper, 2001. Alterations in development, behavior, and physiology in Drosophila larva that have reduced ecdysone production. J. Neurophysiol., 85: 98-104.
CrossRefPubMedDirect Link - Livak, K.J. and T.D. Schmittgen, 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods, 25: 402-408.
CrossRefDirect Link - Lucki, I., 1998. The spectrum of behaviors influenced by serotonin. Biol. Psychiatry, 44: 151-162.
CrossRefPubMedDirect Link - Mentre, F. and S. Escolano, 2006. Prediction discrepancies for the evaluation of nonlinear mixed-effects models. J. Pharmacokinet. Pharmacodyn., 33: 345-367.
PubMed - Monastirioti, M., 1999. Biogenic amine systems in the fruit fly Drosophila melanogaster. Microscopy Res. Tech., 45: 106-121.
PubMed - Nebigil, C.G., D.S. Choi, A. Dierich, P. Hickel, M. Le Meur, N. Messaddeq, J.M. Launay and L. Maroteaux, 2000. Serotonin 2B receptor is required for heart development. PNAS, 97: 9508-9513.
PubMed - Nebigil, C.G., P. Hickel, N. Messaddeq, J.L. Vonesch and M.P. Douchet et al., 2001. Ablation of serotonin 5-HT(2B) receptors in mice leads to abnormal cardiac structure and function. Circulation, 103: 2973-2979.
PubMed - Neckameyer, W.S., 1996. Multiple roles for dopamine in Drosophila development. Dev. Biol., 176: 209-219.
CrossRefPubMedDirect Link - Neckameyer, W.S. and R.L. Cooper, 1998. GABA transporters in Drosophila melanogaster: : molecular cloning, behavior and physiology. Invert. Neurosci., 3: 279-294.
CrossRef - Osborne, R.H., 1996. Insect neurotransmission: Neurotransmitters and their receptors. Pharmacol. Ther., 69: 117-142.
PubMedDirect Link - Pfaffl, M.W., 2001. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res., 29: e45-e45.
CrossRefPubMedDirect Link - Riccio, O., G. Potter, C. Walzer, P. Vallet and G. Szab� et al., 2009. Excess of serotonin affects embryonic interneuron migration through activation of the serotonin receptor 6. Mol. Psychiatry, 14: 280-290.
PubMed - Roy, A., N.J. Brand and M.H. Yacoub, 2000. Expression of 5-hydroxytryptamine receptor subtype messenger RNA in interstitial cells from human heart valves. J. Heart Valve Dis., 9: 256-260.
PubMed - Saudou, F., U. Boschert, N. Amlaiky, J.L. Plassat and R. Hen, 1992. A family of Drosophila serotonin receptors with distinct intracellular signaling properties and expression patterns. J. EMBO, 11: 7-17.
PubMed - Schmittgen, T.D. and K.J. Livak, 2008. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc., 3: 1101-1108.
CrossRefPubMedDirect Link - Sokolowski, M.B., 1980. Foraging strategies of Drosophila melanogaster: A chromosomal analysis. Behav. Genetics, 10: 291-302.
PubMed - Stewart, B.A., H.L. Atwood, J.J. Renger, J. Wang and C.F. Wu, 1994. Improved stability of Drosophila larval neuromuscular preparation in haemolymph-like physiological solutions. J. Comp. Physiol. A, 175: 179-191.
CrossRefPubMedDirect Link - Tierney, A.J., 2001. Structure and function of invertebrate 5-HT receptors: A review. Comp. Biochem. Physiol. [A], 128: 791-804.
CrossRef - Witz, P., N. Amlaiky, J.L. Plassat, L. Maroteaux, E. Borrelli and R. Hen, 1990. Cloning and characterization of a Drosophila serotonin receptor that activates adenylate cyclase. PNAS, 87: 8940-8944.
PubMed - Xu, J., B. Jian, R. Chu, Z. Lu and Q. Li et al., 2002. Serotonin mechanisms in heart valve disease II: The 5-HT2 receptor and its signaling pathway in aortic valve interstitial cells. Am. J. Pathol., 161: 2209-2218.
PubMed