
Ashraf Ahmed Montasser
Department of Zoology, Faculty of Science, Ain Shams University, Cairo, Egypt
International Journal of Zoological Research
Year: 2006 | Volume: 2 | Issue: 3 | Page No.: 204-212







ABSTRACT
The present Scanning Electron Microscopic (SEM) study includes morphological observations on male Echinolaelaps echidninus with particular attention to the undescribed details of its gnathosoma, palpal organ and pulvilli that have rarely described in males of superfamily Dermanyssoidea. The holoventral plate had 23 setae, including 5 pairs in sternogenital region, 5 pairs in preanal region and 3 anal setae. The peritreme extended anteriorly behind coxa 2 from an oval pit situated ventro-laterally between coxae 3 and 4. Gnathosoma consisted of long basis capituli carrying median hypostome and 2 lateral pedipalps. Hypostome had median labium terminated with 2 lobulated plates, cheliceral digits forming pincers-like claw and 2 pointed corniculi. Each pedipalp terminated with tarsus that carried 9 sensillae forming the palpal organ. Small conical sensillae assumed to be chemoreceptors while long broad ones were probably mechanoreceptors. Pulvillis 1 had smooth surface and terminated with 2 small claws and integumental folds. Pulvilli 2-4 had smooth dorsal surface, ventral longitudinal ridges and 2 lateral plates, each with minute processes.
PDF Abstract XML References
How to cite this article
Ashraf Ahmed Montasser, 2006. Scanning Electron Microscopic Observations on Male Echinolaelaps echidninus (Berlese, 1887) (Dermanyssoidea: Laelapidae) with Emphasis on its Gnathosoma, Palpal Organ and Pulvilli. International Journal of Zoological Research, 2: 204-212.
DOI: 10.3923/ijzr.2006.204.212
URL: https://scialert.net/abstract/?doi=ijzr.2006.204.212
DOI: 10.3923/ijzr.2006.204.212
URL: https://scialert.net/abstract/?doi=ijzr.2006.204.212
INTRODUCTION
Dermanyssoidea includes 15 families of mites; most of them are parasites and disease carriers. Of these families, species of Dermanyssidae, Laelapidae and Macronyssidae are proven transmitters of diseases in birds, reptiles and mammals including man (Krantz, 1978).
The spiny rat mite, Echinolaelaps echidninus occurs throughout the world as a parasite of the Norway rat (brown rat), Rattus norvegicus and occasionally of other rodents (Ugbomoiko and Obiamiwe, 1991; Soliman et al., 2001). E. echidninus received the attention of several authors since the start of twentieth century. It is the intermediate host of the protozoan Hepatozoon perniciosum, a parasite of rats (Miller, 1908). Junin virus, the cause of Argentinian hemorrhagic fever, has been isolated from E. echidninus in South America (Parodi et al., 1959).
Studies on E. echidninus were mainly concerned with surveys and host-parasite relationships (Linardi et al., 1984, 1985; Aamir et al., 1985; Ugbomoiko and Obiamiwe, 1991; Soliman et al., 2001). Internal anatomy of E. echidninus through serial sections has investigated by Jakeman (1961). Physiological studies were also conducted on this mite including the oxygen uptake in relation to water balance (Kanungo, 1965), fluctuating humidities and frequency of blood meals on mite survival (Knulle, 1967) and kinetics of water exchange between the mite and the surrounding air (Wharton and Devine, 1968; Devine and Wharton, 1973). Despite the above studies carried out on E. echidninus, detailed description of its mouthparts, sense organs and pulvilli or those of other dermanyssoids, remain sparse due largely to the minute size of these parts.
Substantial literatures have encountered in redescription of mite species with SEM (Desch, 1987; Bukva, 1995; Pence and Gray, 1996; Green and Baker, 1996; Tanskul and Linthicum, 1999). Description of E. echidininus has only investigated with light microscopy in former studies of Keegan (1956) and Tipton (1960).
Description of the gnathosoma of male E. echidninus or that of other dermanyssoids has rarely examined. Detailed morphology of this part is an important step towards elucidation of the feeding behavior of this mite and hence its pathology. SEM has used in describing the gnathosoma of some astigmatic mite species (Dusbabek et al., 1975; Shatrov, 1981; Paran, 1982; Mapstone et al., 2002).
Palpal organ, one of the important sense organs in mites, has a primary significance in relation to orientation responses to humidity, temperature and chemical stimuli. SEM description of palpal organ has previously examined in some gamasid mites infecting nasal cavities of birds (Leonovich, 1998; Leonovich and Staniukovich, 2002).
Detailed morphology of pulvilli is a vital step towards explanation of the attachment mechanism of the mite to its host. Examination of pulvilli in mites was generally rare while that of insects, particularly flies, recognized the interest of some workers (Kupp et al., 2000; Niederegger and Gorb, 2003).
The present study represents a complimentary study to a recent one carried out in female E. echidninus (Montasser, 2006). It includes SEM morphological observations in the male with particular attention to the undescribed details of its gnathosoma, palpal organ and pulvilli.
MATERIALS AND METHODS
E. echidininus of the present study have recovered from R. norvegicus collected from Bilbeis area, about 60 km northeast of Cairo, Egypt. The study area and method of collection have described by Soliman et al. (2001). Mites have separately processed for mounting and identified according to Keegan (1956), Tipton (1960) and Krantz (1978).
E. echidininus were washed several times using saline solution to remove debris. Specimens have fixed in 2.5% glutaraldehyde mixed in phosphate buffer solution (PBS) at a pH of 7.4 at 4°C for 24 h. They have then rinsed twice with PBS at 10 min intervals. Specimens have next treated with 1% osmium tetraoxide at room temperature for 1 day for post-fixation. This has followed by rinsing twice with PBS and dehydrating with alcohol. To replace water in mites with alcohol, they were subjected to increasing concentrations of ethanol as follows 30, 50, 70, 80, 90 and 95% for 15 min each. They have then placed in absolute alcohol for 10 min for 2 changes. Finally, they have subjected to critical point drying in order to complete the dehydration process (Junkum et al., 2004). In order to view specimens, they have first attached with double-sided carbon tape to aluminum stubs, so that they have coated with gold in a sputter-coating apparatus (JEOL JFC-1200). The surface topography of specimens has viewed at 25 kV in a JEOL-JSM5600 scanning electron microscope (Japan) at Central Laboratories Unit, United Arab Emirates University, United Arab Emirates during May 2005.
RESULTS
Male E. echidninus was oval and measured ≈450 μm in length and ≈290 μm in width. The holoventral plate extended anteriorly from the oval transverse genital opening behind the gnathosoma to the rear of the oval longitudinal anal opening (Fig. 1 and 2).
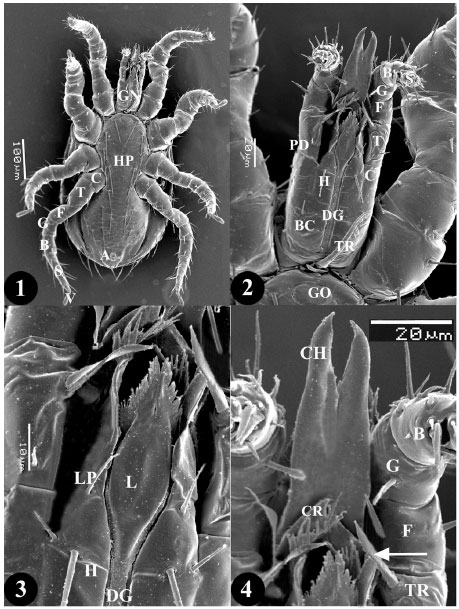 | |
| Fig. 1-4: | Scanning electron micrographs of male E. echidninus. 1. Whole ventral side showing gnathosoma (GN), holoventral plate (HP), anus (A) and leg segments; coxa (C), trochanter (T), femur (F), genu (G), tibia (B), tarsus (S) and pulvillus (V). 2. Whole gnathosoma showing the basis capituli (BC) carrying median hypostome (H) and 2 lateral pedipalps (PD). It also showed deutosternal groove (DG), genital opening (GO), tritosternum (TR) and pedipalp segments; coxa (C), trochanter (T), femur (F), genu (G), tibia (B) and tarsus (S). 3. Higher magnification of lower part of gnathosoma showing hypostome (H) carrying median labium (L), in front of the deutosternal groove (DG), and 2 lateral flattened plates (LP). 4. Higher magnification of upper part of gnathosoma showing cheliceral digits (CH) forming a pincers-like claw, pointed corniculi (CR) and leaf-like seta (arrow) on the trochanter (TR). B, tibia; F, femur; G, genu |
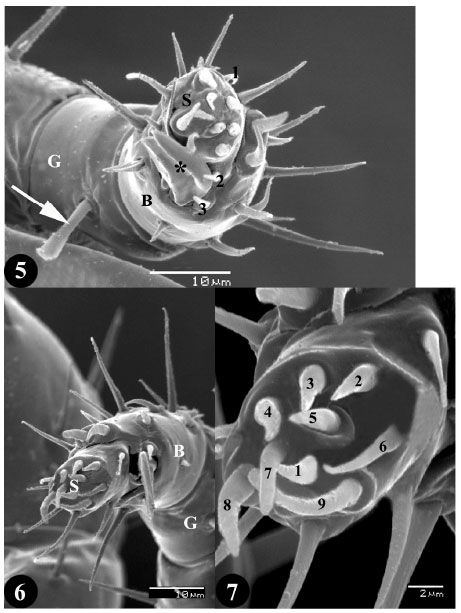 | |
| Fig. 5-7: | Scanning electron micrographs of palpal organ of male E. echidninus. 5. Terminal segments of right pedipalp showing club-like process (arrow) on Genu (G) and 2 extremely robust (*) and 3 minute setae (1-3) terminal to tibia (B) beside tarsus (S). 6. As in Fig. 5 but for the left pedipalp showing complete set of terminal setae of tarsus (S) forming the palpal organ. B, tibia; G, genu. 7. Higher magnification of palpal organ formed of 5 small conical (1-5) and 4 long broad setae (6-9) |
 | |
| Fig. 8, 9: | Scanning electron micrographs of pulvilli of male E. echidninus. 8. Pulvillus of 1st leg showing its smooth surface and few terminal integumental folds and 2 small claws. 9. Pulvillus of 3rd leg showing its dorsal smooth surface, ventral longitudinal ridges and minute processes on lateral plates shown by arrow |
It laterally expanded covering most of the opisthosomal area and faintly ornamented with wavy striations. The holoventral plate had 23 setae, including 5 pairs in sternogenital region, 5 pairs in preanal region and 3 anal setae (Fig. 1). The first five pairs of setae were distinctly longer and stouter than other setae on the plate. Adanal setae were much thinner and one third as long as the postanal seta. The peritreme extended anteriorly behind coxa 2 from an oval pit situated ventro-laterally between coxae 3 and 4 (Fig. 1).
Gnathosoma, or capitulum, consisted of long basis capituli carrying median hypostome and 2 lateral pedipalps (Fig. 2). Short tritosternum noticed at the ventral side of the gnathosomal base. It was bifurcated into two sparsely feathered laciniae. Deutosternal groove was located on the ventral side of the basis capituli (Fig. 2). Oval shaped labium observed in front of deutosternal groove (Fig. 2 and 3). Labium carried two fronto-lateral and medially separated plates, each had at least 15 lobules (Fig. 3). Lateral to labium, hypostome carried two long flattened plates, each terminated with 12-15 setiform processes (Fig. 3). Cheliceral digits were edentate and exceedingly elongated. They have united at the base and terminally formed a pincers-like claw (Fig. 4). The corniculi were ventrally located to chelicerae. They were well sclerotized, horn-shaped and pointed. Leaf-like processes originated at the base of each corniculus (Fig. 4).
Each pedipalp consisted of six segments. Beginning with the most proximal, they named as coxa, trochanter, femur, genu, tibia and tarsus (Fig. 2 and 4). Tarsus located at about the terminal half of tibia (Fig. 5). Pedipalp segments mostly carried small and pointed setae. However, trochanter carried leaf-like seta directed towards labial processes (Fig. 4) and genu carried club-like seta near chelicera (Fig. 4 and 5). In the tibial terminal area beside tarsus, tibia carried two extremely robust and elongated setae with common base (Fig. 5). Three minute setiform sensillae with pointed tips and broad bases were observed on the tibial terminal; 2 adjacent to tarsus and 1 at the base of robust setae (Fig. 5). Tarsus had a terminal cluster of nine setiform sensillae that might characterized according to their location, shape, size, tip, socket and surface of the wall (Fig. 6 and 7). According to size, sensillae almost divided into 2 main groups; small-sized 1-5 and large-sized 6-9 (Fig. 7). Sensillae 1-5 had conical shapes with broad bases and pointed tips. Sensillae 3-5 had slightly elevated sockets. Sensillae 1 and 5 were centrally located while those 3-5 were located in part of the periphery. Sensilla 6 had finger shape, broad base, no socket and double the length of sensillae 1-5. Sensilla 7 appeared broader than sensilla 6. Sensilla 8 was robust with pointed tip and slightly elevated socket. Sensilla 9 was the longest and had apparent socket. Sensilla 7 situated medially while sensillae 6, 8 and 9 were peripherally located. All sensillae had smooth surfaces (Fig. 7).
Four pairs of walking legs were located on the anterior half of the ventral side; each carried small-medium sized pointed setae and divided into 6 segments (Fig. 1). Beginning with the most proximal, they were coxa, trochanter, femur, genu, tibia and tarsus. The latter hanged terminal pulvillus through a narrow neck (Fig. 1 and 8). The respective lengths of legs (including pulvilli) were as follows: 1- 344 μm, 2- 272 μm, 3- 268 μm and 4- 392 μm. Pulvillus of 1st leg had a rectangular shape, smooth surface and 2 terminal curved claws and few integumental folds (Fig. 8). Each pulvillus of 2-4 legs had dorsal smooth surface, ventral longitudinal ridges and terminated with integumental folds and 2 lateral plates, each had 4 or 5 min processes (Fig. 8 and 9).
DISCUSSION
Morphological characters of male E. echidninus including coalescing of the ventral plates into one holoventral plate greatly matched with that described in Laelaps (Tipton, 1960) or Echinolaelaps species (Tenorio, 1975). In the present study, holoventral plate carried 23 setae. This number resembled that noticed in male E. joaquini and E. papagomyis described by Tenorio (1975).
In the present study, cheliceral digits have united at their base forming pincers-like claw. This modification might point to grasping habit of male E. echidninus that result in abrasion to the host skin. This pattern not observed in female (Montasser, 2006). Cheliceral modification has considerably noticed among gamasid taxa where there was a correlation between cheliceral structure and the type of food taken (Karg, 1961). Fusion of cheliceral digits has previously mentioned in actinedid mite Brevipalpus cardinalis (Krantz, 1978). Ventral labium and 2 lateral lips appear to enclose the mouth opening. Similar pattern has noticed in Psoroptes ovis (Mapstone et al., 2002). According to the above structures, chelicerae may share in cutting the host skin and labium and 2 lateral lips share in transferring the blood into the mouth. Supporting the latter suggestion is the presence of many processes terminating the labial palp and lateral lips. The tritosternum below deutosternal groove probably function in fluid transport to the buccal region. This supports an earlier suggestion of tritosternum function by Wernz and Krantz (1976).
Pedipalp structure of male in the present study with 6 segments, position of the tarsus on part of the tibial terminal and presence of robust setae on the tibia were greatly similar with that described in female E. echidninus (Montasser, 2006). Presence of leaf-like seta on the trochanter towards the labial processes and club-like seta on the genu near the chelicera were specific features for male E. echidninus rather than female. Presence of these setae near mouth parts probably share in the feeding process such as removing solid material from entering the oral cavity.
Acarine pedipalps predominantly terminated with sensory sensillae, or palpal organ, which aid in locating their food sites (Krantz, 1978). In the present study, palpal organ consisted of 9 setiform sensillae. Ten sensillae have observed in case of female E. echidninus (Montasser, 2006). This might be due to the smaller size of male that probably needs fewer sensillae for food acquisition rather than in females. Five palpal sensillae of male E. echidninus were small in size and with broad base and pointed tips. These features support the olfactory or chemosensory function described in some gamasid species (Leonovich, 1998; Leonovich and Staniukovich, 2002; Montasser, 2006). On the other hand, 4 long sensillae were observed in palpal organ of male E. echidninus instead of 2 in case of female (Montasser, 2006). This probably supports a faster mechanical response in case of male rather than in female. Long palpal sensillae have suggested to possessing a mechanoreceptor function by the above authors.
Pulvilli of male E. echidninus greatly resembled those described in female (Montasser, 2006) with some exceptions. Pulvillus 1 in the present study had a rectangular shape with fewer terminal integumental folds and minute claws. As in female, this identity probably helps the mite to slide over and adhere to the host skin particularly during feeding process where the forelegs have anteriorly directed beside mouthparts. This supports the function of leg 1 in most acarines in which it makes the initial contact with food substrate (Krantz, 1978).
Pulvilli 2-4 appeared as spatula-like which may provide a sliding motion on the smooth surfaces as previously described in the gecko (Autumn et al., 2000) or in the grasshopper (Gorb and Scherge, 2000). The latter authors returned this motion to the presence of distally sloped microfilaments of the pulvillus material. Niederegger and Gorb (2003) added another function of the pulvilli in flies included the secretion of a liquid for adhesion to smooth surfaces. In general, presence of microfilaments or liquid secretion by mite pulvilli needs further histological or physiological studies. In male E. echidninus, longitudinal ridges on the ventral side of pulvilli 2-4 probably share in firm attachment with the host skin.
ACKNOWLEDGMENTS
The author wishes to express his deep appreciation and gratitude to Prof. Aleya S. Marzouk, Prof. Rashid M. Rashid, Zoology Department, Faculty of Science, Ain Shams University, Egypt and Mr. Essam Shaaban, Central Laboratories Unit, United Arab Emirates University, United Arab Emirates for their invaluable assistance and outstanding technical support throughout the study.
REFERENCES
- Autumn, K., Y.A. Liang, S.T. Hsieh, W. Zesch and C.P. Wal, 2000. Adhesive force of a single gecko foot-hair. Nature, 405: 681-685.
CrossRef - Desch, C.E.Jr., 1987. Redescription of Demodex nanus (Acari demodicidae) from Rattus norvegicus and R. rattus (rodentia). J. Med. Entomol., 24: 19-23.
Direct Link - Devine, T.L. and G.W. Wharton, 1973. Kinetics of water exchange between a mite Laelaps echidnina and the surrounding air. J. Insect Physiol., 19: 243-254.
PubMed - Dusbabek, F., W. Sixl and L. Scherr, 1975. Scanning micrographs of the first legs and gnathosoma of two species of myobiidae (Acarina trombidiformes). Folia Parasitol., 22: 375-378.
PubMed - Green, E.D. and C. Baker, 1996. Observations on the micromorphology of the tropical rat mite Ornithonyssus bacoti (hirst) as revealed by scanning electron microscopy. J. S. Afr. Vet. Assoc., 67: 128-132.
PubMed - Junkum, A., A. Jitpakdi, N. Komalamisra, N. Jariyapan, P. Somboon, P.A. Bates and W. Choochote, 2004. Comparative morphometry and morphology of anopheles aconitus from B and C eggs under scanning electron microscope. Rev. Inst. Med. Trop. de Sao Paulo, 46: 257-262.
PubMedDirect Link - Kanungo, K., 1965. Oxygen uptake in relation to water balance of a mite (Echinolaelaps echidninus) in unsaturated air. J. Insect Physiol., 11: 557-568.
PubMed - Knulle, W., 1967. Significance of fluctuating humidities and frequency of blood meals on the survival of the spiny rat mite Echinolaelaps echidninus (berlese). J. Med. Entomol., 4: 322-325.
Direct Link - Kupp, P.G., J.I. Phillips and E. Garton, 2000. The pulvillus and empodium in Culex quinquefasciatus visualization with the light microscope and a study of fine structure with the scanning electron microscope. J. Am. Mosq. Control Assoc., 16: 166-170.
Direct Link - Leonovich, S.A., 1998. The palpal receptor organ of gamasid mites (Mesostigmata gamasina). Parazitologiia, 32: 258-263.
PubMed - Leonovich, S.A. and M.K. Staniukovich, 2002. Comparative study of the sensory system in gamasid mites Rhinonyssus rhinolethrum, Rh. Subrhinolethrum and Ptilonyssus motacillae (mesostigmata gamasina rhinonyssidae) parasites in the nasal cavity of birds. Parazitologiia, 36: 390-395.
PubMed - Linardi, P.M., J.R. Botelho, H.C. Cunha and N.D. Moreira, 1984. Ectoparasites of rodents of the urban region of belo horizonte mg I interaction between ectoparasites and hosts. Mem. Inst. Oswaldo. Cruz., 79: 239-247.
PubMed - Linardi, P.M., J.R. Botelho and H.C. Cunha, 1985. Ectoparasites of rodents of the urban region of Belo Horizonte, MG. II. Variations of the infestation indices in Rattus norvegicus norvegicus. Mem. Inst. Oswaldo. Cruz., 80: 227-232.
Direct Link - Mapstone, S.C., A. Beasley and R. Wall, 2002. Structure and function of the gnathosoma of the manage mite Psoroptes ovis. Med. Vet. Entomol., 16: 378-385.
PubMed - Montasser, A.A., 2006. The spiny rat mite Echinolaelaps echidninus (Berlese, 1887) (Dermanyssoidea: Laelapidae): Redescription of the female with emphasis on its gnathosoma, sense organs, peritreme and pulvilli. Int. J. Zool. Res., 2: 1-13.
CrossRefDirect Link - Niederegger, S. and S. Gorb, 2003. Tarsal movements in flies during leg attachment and detachment on a smooth substrate. J. Insect Physiol., 49: 611-620.
CrossRef - Paran, T.P., 1982. Functional morphology of the gnathosoma in Myobia musculiSchrank 1781 trombidiformes-acari. Acarologia, 23: 348-357.
Direct Link - Shatrov, A.B., 1981. Morphofunctional characteristics of the larval mouth apparatus of the chigger Neotrombicula pomeranzevi (trombiculidae). Parazitologiia, 15: 10-20.
PubMed - Soliman, S., A.J. Main, A.S. Marzouk and A.A. Montasser, 2001. Seasonal studies on commensal rats and their ectoparasites in a rural area of Egypt the relationship of ectoparasites to the species locality and relative abundance of the host. J. Parasitol., 87: 545-553.
Direct Link - Tanskul, P. and K.J. Linthicum, 1999. Redescription of Leptotrombidium (Leptotrombidium) imphalum (Acari trombiculidae) with observations on the bionomics and medical importance in Northern Thailand. J. Med. Entomol., 36: 88-91.
Direct Link - Tenorio, J.M., 1975. Three new species of Laelaps echinolaelaps from Indonesia acari Laelapidae. J. Med. Entomol., 12: 77-86.
Direct Link - Ugbomoiko, U.S. and B.A. Obiamiwe, 1991. Distribution and incidence of ectoparasites on small mammals in a rainforest belt of Southern Nigeria. Angew. Parasitol., 32: 143-148.
PubMed