
Ignacy Kitowski
Department of Nature Conservation, Institute of Biology, Marla Curie-Sklodowska University, Akademicka 1 9, PL-20-033 Lublin, Poland
International Journal of Zoological Research
Year: 2006 | Volume: 2 | Issue: 2 | Page No.: 169-177







ABSTRACT
Breeding behaviour of Eurasian marsh harriers Circus aeruginosus pairs nesting in marshes, water reservoir and fish pond of SE Poland were compared. Harriers nesting on the marshes were most efficient during aerial prey transfers, which could suggest that the habitat was taken by experienced, better quality birds when set against the ones occupying nests on the ponds and reservoir. Additional evidence that higher quality birds nested on marshes is that egg laying was earlier there. Marsh Harriers performed U-shape undulation, in the nestling period and these appeared more intense for marsh nesters as well.
PDF Abstract XML References
How to cite this article
Ignacy Kitowski, 2006. Breeding Behaviour of Eurasian Marsh Harriers (Circus aeruginosus L., 1758; Aves, Accipitridae) Nesting on Three Habitats in Eastern Poland. International Journal of Zoological Research, 2: 169-177.
DOI: 10.3923/ijzr.2006.169.177
URL: https://scialert.net/abstract/?doi=ijzr.2006.169.177
DOI: 10.3923/ijzr.2006.169.177
URL: https://scialert.net/abstract/?doi=ijzr.2006.169.177
Introduction
Eurasian marsh harrier Circus aeruginosus is typical raptor in that its populations strongly benefited from decrease DDT use and introduction of the wide law protection. The changes resulted in growing the species population in many European countries (Bavoux et al., 1997; Underhill-Day, 1998; Heath et al., 2000; Daroczi and Zeitz, 2003) and expanding into new habitats for breeding. Similar changes have been observed in Poland, where following introduction of the new protection law in the middle 70’s the number of breeding pairs tend to grow and new habitats have been occupied (Buczek and Keller, 1994; Kitowski, 2000a; Tomialojc and Stawarczyk, 2003).
It is thus fairly likely that individual qualities and experience of raptors can significantly impact the choice for different habitats to be preferred for occupation (Lohmus, 2004). In such conditions can exist discrimination in quality of individuals in relation to the type of occupied habitat. All the more so as well documented that individual quality and habitat quality may interact. Body condition or measurements are related to site quality and older birds can occupied better sites (Moore and Henny, 1984; Newton, 1991; Hakkarainen and Kropimaki, 1996). It suggest that birds of better habitat quality can perform better parental care. Marsh Harrier as raptor expanding into new habitats can be good model for study the relation between quality of parental care and quality of occupied territories. It can be supported well known breeding ecology (Witkowski, 1989; Altenburg et al., 1987; Underhill- Day, 1998) and behavioural patterns (Fernandez, 1992; Fernandez and Azkona, 1994). The research has been aimed at comparing patterns of parental care of Eurasian Marsh Harrier breeding pairs in three different habitat conditions in south-east Poland.
MATERIALS AND METHODS
The research was performed in 2001-2002 on three sites of frequent occurrence of the species in south-east Poland within 15 km of Chelm town (51°08 'N, 23°30 'E, SE Poland). The first habitat is in the ”Rozkosz” reserve (in the vicinity of Plawanice village). There 21-44 pairs of Eurasian Marsh Harrier breed yearly depend on 601 ha (Buczek and Keller, 1994; Kitowski I.- unpublished data). This study area consists of 3 main biotopes: marshes covered with Saw Sedges Cladium mariscus), Sedges Carex spp. and Reed beds Phragmites australis extensive meadows and arable fields where mainly cereals and beetroots are cultivated (Buczek and Buczek, 1996; Kitowski I. -unpublished data). The second breeding site was in the area of Strupin Maly a small reservoir (approximately 1.6 km2) where Harriers nested on reedbeds Phragmithes australis and reed maces Typha spp. Apart from Harriers the reservoir is also inhabited by colonies of Whiskered terns Chlidonias hybridus, Black terns Chlidonia niger and Black-head gulls Larus ridibundus (totally aproximately 150 pairs). Every year 3-5 breeding pairs of Marsh Harrier are observed here (Kitowski I. -unpublished data). The third site was on Pobolowice fish ponds, where every year on reedbeds Phragmites australis 3-5 breeding pairs of Marsh Harrier are found (Kitowski I. -unpublished data). Similar breeding situations have been reported by other researchers of other harrier species (Simmons, 1988; Pandolfi and Pino d’Astore, 1994; Wiacek, 2004) adult individuals were not captured to mark since it might have induced them to abandon their broods. Parental individuals were distinguished by individual characteristics including molt and individual differences in plumage. While observing, data on differences in plumage, was recorded by means of drawings and photos of particular individuals in order to clearly identify each one. Both binocular (10x60) and a telescope (12x60) were used for distance watching (250-300 m.). Nine prey categories have been determined, namely: 1) small mammal-typically Voles Mictrotus sp. (m = 20 g), 2) larger mammals such as hares Lepus europeanus juv. (m = 200 g), or water vole Arvicola terrestris (m = 80 g) (Zawadzka 1,999), 3) small birds which included sparrows Passer spp. (m = 20 g), Wagtails Moticilla spp. (m = 20 g), 4), larger birds such as partridge (Perdix perdix) (m = 400 g), or domestic hen Gallus domesticus (m = 1000 g), Coots Fulica atra fledglings (m = 550 g), 5) Amphibia-Anura namely frogs (m =10 g), 6) Reptilia which consisted of lizards Lacerta spp., (m = 10 g), 7) Large insects such Tettigonioidea following (Kitowski, 2000b) mass of grasshoppers Tettigoniidae was assumed as mass great green bush cricket Tettigonia viridissima individual -0.7 g (Kitowski, 2000b), 8) large bits of carrion, namely dead small dogs m = 500 g, 8) dead fish-small carp,Carpinus carpio - 600 g. Totally parental behaviour was observed in 9 nests located in fish ponds (n = 3), marshes(n = 3) and water reservoir (n = 3). A total number of 540 h of watching have been performed on 2, 12, 22, 32 and 42th day after the oldest chick hatching. Following Fernandez (1992) the nestling period of marsh harrier was divided into an early and late nestling period. That covered 300 h of watching performed in the early nestling period and 240 h in the late nestling period. Studies were carried out from middle June till late July, when juveniles started to perform the first flights. At each nest 5 observational sessions (a total of 60 h) were performed.
RESULTS
Prey Transfers
A total of 349 prey was observed to being brought to nest during breeding season. Males hunted 266 (76.2%) prey, while merely 83 (23.8%) prey were hunted by females (χ2 = 95.6, p<0.01). All prey caught by females, were delivered directly to the nest. Prey caught by ales were elivered directly to the nest a 53 (20%, n = 266) items.
| Table 1: | Modes of food pass from male to dependants Marsh Harrier during nestling period when females present in breeding area. A: Direct transfer to the nest, B: Transfer on the ground, C: Aerial transfers between mates |
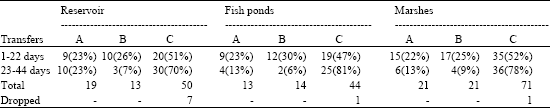 | |
The remaining prey had been transfered to females either on the ground, or in the air in respective numbers of 48 and 165 cases, which contributed to 18%, n = 266 and 62.0%, n = 266, respectively (Table 1). Following the prey transfer, females delivered it to nests. During 9(5%, n = 165) aerial transfers the prey was dropped, but all the prey was retrieved by females and subsequently delivered to nests. Prey (n = 8) was most likely to be dropped in the early nestling period. Seven aerial failed transfers were spotted at Strupin site (6 in the early and 1 in the late nestling period). At Strupin site in all observed nests events of prey drops were observed. In latter sites prey drops concerned to the one particular nest.
All drops during aerial transfers appeared from poor coordination between birds involved. Only one drop (at Plawanice site) resulted from food robber attempt by a male Montagu’s Harrier Circus pygargus. In parallel to the growing nestlings, a distance from the nest at which the aerial transfers were performed (r = 0.743, n = 45, p<0.001) tend to grow, so in the late nestling period nearly 81% of aerial transfers was done over meadows and fields far (d>250 m) from nests. Change in prey transfers techniques applied in the early and late nestling period for ground and aerial transfers were found. The respective numbers and percentages were as follows: ground prey transfer 39(26.7%, n =146) vs 9(7.5%, n = 120): χ2 = 15.2, df = 1, p<0.001), aerial prey transfers 74(50.1%, n = 146) vs 91(75.8%, n = 120): χ2 = 16.3, df = 1, p<0.001. No difference was detected for direct prey transfers to the nest, in early and late nestling period the following number were recorded: 33(22.6%, n = 146) vs 20(16.7 %, n = 120): χ2 = 1.11, df = 1, ns (Table1).
Frequency of Delivered Prey Number and Biomass Delivery to the Nest
In the early nestling period adults delivered 0.53 prey/h, whereas in the late nestling period they delivered 0.80 prey items/h. As fledglings grew parents made more food deliveries daily (r = 0.46, n = 45, p<0.001). Biomass supplied to the nest also increased (r = 0.44, n = 45, p<0.001), though it was accompanied by a decrease in both number (r = - 0.84, n = 45, p<0.001) and biomass (r = -0.77, n = 45, p<0.001) of prey that is delivered by males. On the marsh site 142 prey deliveries at rate of 0.79 prey/h (48,7g biomass prey/h) was recorded. A number of 8 (5.6%, n = 142) of which was disturbed by diving waders (n = 2) and cleptoparasitic attacks of Montagu’s Harriers (n = 5), as well as other Eurasian Marsh Harriers (n = 1), but no prey was lost. On the reservoir 114 prey were supplied to the nest, (0.63 prey/h, 10.5 g biomass prey/h), out of which 31 (27.1%) were disturbed by other birds diving. On fish ponds 93 deliveries to the nest were performed. The respective frequency amounted to 0.52 prey/h, (28.8 g biomass prey/h).
During 7 transfers the birds carrying prey were unsuccessfully attacked by other diving Marsh Harriers adults from neighbour nests. For all 9 nests on average 0.73 prey items/h was delivered to breeding sites, which corresponded to 29.3 g of biomass prey/h. On marsh sites an average of 0.95±0.05 of total daily number prey deliveries were not disturbed and at the reservoir sites an average of 73.0±0.08 of total number of prey deliveries were performed undisturbed; the same value at the fish pond sites amounted to 91.0±0.05.
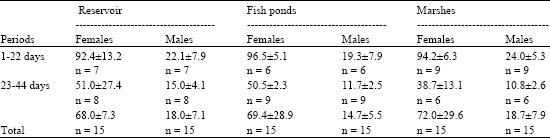
| Table 2: | Time spent in the nesting area (as% of total time of observation) by males and females Harrier during nestling period. n-number observatio nalsessions |
 | |
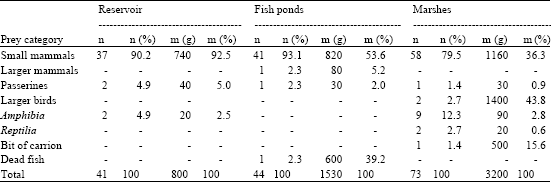
| Table 3: | Prey delivery in the early nestling period |
 | |
| Table 4: | Composition of food delivered to the nest in late nestling period |
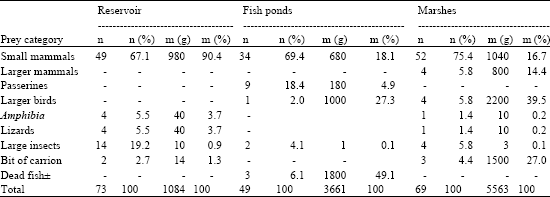 | |
The average over all the nests under observations, a86.0±0.15 of total daily number of prey deliveries were not disturbed. Differences were found in the daily rate of undisturbed food deliveries between the habitats (ANOVA: F3,59 = 280, p<0.001). In the early nestling period, up to the 11th day after hatching, females were rarely involved in providing the nests (Table 1 and 2) and they delivered to the nest only 0.074±0.02 of total number of prey on average. The values ranged from 0.049 to 0.09 of total number of prey. In the late nestling period females were observed to have carried to the nests on average 0.39±0.09 of prey, with the respective range of total number of prey between 0.29 and 0.41. Males carried the prey from places located far away from the nest (d>2 km), whereas females hunted within the areas close to the nests and they managed to hunt successfully only a small number of relatively small mass prey from water environment. For a reservoir - 4 prey of a 40 g mass, which gave 3.7% of biomass, on marshes -1 prey only that weighed 10 g, i.e., 3.7% of the biomass. The ponds were exceptional in that respect, since females managed to retrieve from water 3 dead carps Carpinus carpio of a total mass of 1800 g (Table 3 and 4).
| Table 5: | Frequency (%) of the material supply to the nest by the Marsh Harrier in the study area |
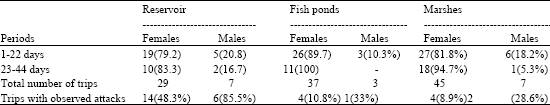 | |
Nest Building
A total number of 128 (n = 128) events delivering to the nest material (Table 5) was observed. After hatching the nestlings, in the early nestling period a need to extend the nest have rised. Thus females performed 111 (88%) nest material supplies trips, while males only a number of 17 (13%) trips. Differences in frequency between sexes were significant (χ2 = 69.03, df = 1, p<0.001). At the fish pond site and marshes in the late nestling period males role in delivering nest material did significantly drop (Table 5). At the reservoir sites males share in delivering nest material was the highest one and it reached respectively 21 and 17% in the early and late nestling period. 31 (24%, n = 128) events of dives on Harriers carrying nest material were recorded. Males were attacked by diving only 9 times, while females 22 (χ2 = 5.45, df = 1, p<0.02). All attacks were undertaken by Terns or Gulls. In the reservoir a high number of 14 (48 %, n = 29) female victims with nest material was observed, while males with nest material were seen to have been attacked only 6 times (86%, n = 7). Four (3.1%, n = 128) trips consisted in delivering twigs with fresh green leaves, or grass to the nest in the late nesting period. Males (n = 3) nesting on the reservoir flew shortest time (18.0±6.0 sec) with nest material in the direct vicinity of nest (d<25 m) when compared to the males (n = 7) nesting on marshes (35.0±7.0 sec) and the ones (n = 7) nesting on the fish ponds (42.0±6.0 sec) (Kruskal - Wallis ANOVA: H = 6.22, df = 2, p<0.05). The building activities were most intense in the morning. Most, n = 63 (49%) of nest material supplies trips were performed in 800 -1100. Other nest material supplies trips were performed later: 1100 - 1400, n = 19 (15%) and 1400 - 1700, n = 14 (11%) and 1700 - 2000, n = 32 (25%). Differences in frequency of daily distribution of nest material supplies trips were significant (χ2 = 54.4, df = 3, p<0.001).
Brood Defences and Aerial Displays
In the early nesting period females spent 0.95±0.09 of total time of observation in the nesting area (Table 2). In the late nestling period this rate amounted to 0.42±0.22. Males were observed to spend there only 0.22±0.06 and 0.134±0.03 of the total observation time in the early and late nestling period, respectively. Overall number of n = 109 events of territorial defences by adults were observed, out of which n = 78 (71.6%) was conspecifics oriented. Males were more frequently involved in harassing conspecifics in comparison to females: 56 vs. 22; χ2 = 14.2, df = 1, p<0.0002. A number of n = 28 sky dance displays sessions were observed, 24 of them performed by males, whereas only 4 by females; (χ2 = 14.3, df = 1, p<0.0002). That yielded intensity of 5.2 display/100 h for the studied birds. Intensity of males displays was higher in comparison to females (4.4 displays/100 h vs. 0.74 display/100 h). On the marshes males performed n = 14 sky dances sessions, at which 9.8±2.0, ranging 6-14, U-shape undulations (Simmons, 1991a) were observed. For fish pond sites and the reservoir a respective n = 6 sessions (with 8.2± 2.6, ranging 6 - 12) and only n = 4 such sessions (with 5.7±2.2, ranging 4 - 9) U-shape undulations were noted. Both for displays (χ2 = 7.0, df = 2, p<0.03) and the number of U-shape undulations (Kruskal-Wallis ANOVA: H = 7.54, df = 2, p<0.02) in particular habitats the differences in frequency were found. Males more frequently displayed in the early than late nestling period: 20 vs 4 sessions (χ2 = 10.7, df = 1, p<0.001). Early nestling period displays contained more U-shape undulations than late nestling period ones (Mann - Whitney U test: Z = -2.48, n1 = 20, n2 = 4, p<0. 0124).
Breeding Phenology and Breeding Effects
Pairs nesting in marshes initiated breeding at the earliest, laying the first eggs on the 116.8±4.0 day of the year (median -116.6 day, all data in Julian days) in 14 clutches (4.6±0.9 eggs, median 5 eggs) found during studies. Fish ponds pairs performed laying on 120±2.2 day (median - 120 day) in 7 clutches (4.1±0.9 egg, median: 4 eggs). Reservoir pairs were the last ones to have laid eggs, i.e., on the 123.5±3.0 day (median: 124 day of year) in 6 clutches with eggs (3.7±0.8, median: 3.5 eggs). The differences in clutch size between the habitats were found insignificant: Kruskal-Wallis ANOVA: H = 4.60, df = 2, ns, but the first egg laying dates were proven to be significant: Kruskal-Wallis ANOVA: H = 10.6, df = 2, p<0.02.
A total number of 19 juveniles were fledged from 9 nests under study. On marshes, ponds and reservoir respectively 8, 7 and 4 youngs were fledged, which corresponded to an average values of 2.7±0.58, 2.3±0.58 and 1.3±0.58.
DISCUSSION
Harriers nesting on the marshes were most efficient during aerial prey transfers, raising the possibility that is habitat was occupied by experienced, better quality birds when set against the ones occupying the nests on the ponds and reservoir. It seems that efficiency of aerial prey transfers can serve as a reliable indicator that helps to determine the quality of mates, since during aerial transfers they have to integrate many factors related to their body condition, flying and catching capabilities, which in turn may reflect the quality of particular individuals. The study confirmed smaller feeding frequency and biomass prey delivered to nestlings by pairs nesting on the fish ponds and reservoir compare to the pairs nesting on marshes. Another evidence of better quality birds that nested on marshes was the earliest date of egg laying. The date of arrival at the breeding area, is considered to be a clear indicator of breeding quality of nesters, since younger, less experienced birds, in poorer condition, tend to arrive late at the nesting area and start breeding late (Stutchbury and Robertson, 1988; Brown and Bomberg- Brown, 1996). High percentage of large insects and that were delivered to fledglings in the late nestling period also contributes to the idea reservoir nesters are lower quality which occupied lower quality areas. Insects are easy to catch and are widely quoted as exemplary prey of inexperienced in foraging abilities individuals (Shrubb, 1982; Toland, 1986; Varland et al., 1991). However, also phenomenon may be partially related summer abundance due to demographic explosion of these insects.
Cases of starting breeding in worse quality habitats by inexperienced younger birds, in comparison to adult individuals, were also recorded in other raptors (Moore and Henny, 1984; Newton, 1991; Wiklund, 1996). Apart from high contribution of insects in food delivered to nests overall food composition of food in studied Eurasian Marsh Harriers did not deviate from the pattern recorded for other well known European populations, supporting strongly the basic role of voles (Microtus sp.) and birds of water habitats in the diet (Hilden and Kalinanen, 1966; Schipper, 1977; Underhill-Day, 1985; Witkowski, 1989). The carrion cases deserve attention since it can be interpreted in terms of opportunistic hunting strategy in the studied species.
The highest number of drops in the course of aerial prey transfers was recorded in the early nestling period. Apart from the individuals quality, it can be contributed to early development of the breeding cycle. Other harriers research (Simmons, 1991b; Kitowski, 2001; Fernadez and Azkona, 1994) also demonstrated more frequent failed transfers in the early parts of breeding when mates just started to learn their habits. Prey availability can be another factor shaping prey transfers patterns between mates of Harriers. In Hen Harrier Circus cyaneus it was observed (Simmons et al., 1987) that males provisioned nest directly only 4.5% of the time of 'high vole year', but 63% of the time in 'poor vole year'. It resulted from greater absence of females from nests in poor years, when they foraged more often. In SE Poland only about 20% of prey was delivered directly to nest by males, which could be contributed to the high abundance of prey in the foraging area.
As reported elsewhere (Witkowski, 1989; Fernandez, 1992) I observed a drop in the role that males play as food providers the course of the nestling period. Still, at nests heavily impacted by other birds (reservoir) the males role did not tend to cease that drastically, which has most likely resulted from better flexibility of their bodies. Witkowski (1989) noted that males flying around with nest material may be also of an informative character to advertise holding of territory. It could also be essential in studied Marsh Harriers, though such behaviour on the reservoir was modified by mobbing gulls and terns.
Nest material provides a microhabitat for bacteria, fungi, insects and mites including pathogenic microorganisms (Hamilton and Zuk, 1982; Singleton and Harper, 1998) and carrying greenery to the nest may have a sanitation function (Clarke and Manson, 1985; Witkowski, 1989). It was observed that intense gathering of nest material by Eurasian Marsh Harriers happened to occur in the mornings. Most likely it results from the necessity to reform and dry the platform structure after night when building activity has been stopped and the nest is considerably wetter (Fernandez, 1992).
It was shown that Eurasian Marsh Harriers perform U-shape undulation flights typical for the pre-nuptial period of Circus species (Simmons, 1991a; Pandolfi and Barocci, 1994), also in the nestling period. Complex and intense aerial sky dances are considered to be a likely factor indicating the quality of males which plays a basic function in choosing mate in Harriers (Simmons, 1991a; Wiacek, 2004). In such terms the observations support also better quality males of Marsh Harriers nesting on marshes when compared to the males from the fish ponds and reservoir. They were definitely of higher intensity. In Harriers sky dances, apart from their nuptial role, can also perform a defensive function (Powers et al., 1984; Kitowski, 2003; Arroyo et al., 2001) that makes them to be likely observed in Eurasian Marsh Harriers in late nestling period and even later in the post-fledging period (Kitowski I.-unpublished data). Still, it was noted in African Marsh Harrier (Simmons, 1991a) and Montagu’s Harrier (Wiacek, 2004) that sky dances intensity, is the highest in the pre-laying period and tends to decrease with breeding cycle development. Nevertheless, their number was significantly lower that the number of performed sessions by females of African Marsh Harriers which during all breeding cycles were found to have performed 30.3% of n = 142 observed sky dances sequences (Simmons, 1991a).
In conclusion, the study on Eurasian Marsh Harriers as the raptor expanding for new breeding habitats showed that there is discrimination in parental care behaviour and breeding effects between habitats what reflects quality of individuals.
ACKNOWLEDGMENTS
I thank Prof. M. Pandolfi (University of Urbino, Urbino Italy), Dr K.Bildstein (Hawk Mountain Sanctuary Assoc, Kempton, USA), Prof. Tryjanowski Piotr (Dept. Behavioural Ecology, University of Adam Mickiewicz, Poznan, Poland) for creative comments on previous drafts of the paper and some English language improvements.
REFERENCES
- Arroyo, B.E., F. Mougeot and V. Bretagnolle, 2001. Colonial breeding and nest defence in Montagu's Harrier (Circus pygargus). Behav. Ecol. Sociobiol., 50: 109-115.
CrossRefDirect Link - Clarke, L. and J.R. Manson, 1985. Use material as insecticidal and antipatogenic agents by the European starling. Oecologia, 67: 169-176.
CrossRefDirect Link - Fernandez, C., 1992. Nest material supplies in the marsh harrier Circus aeru-gi-nosus: Sexual roles, daily and seasonal activity patterns and rainfall influence. Ardea, 80: 281-284.
Direct Link - Hakkarainen, H. and E. Kropimaki, 1996. Competitive and predatory interaction among raptors: An observational and experimental study. Ecology, 77: 1134-1142.
CrossRefDirect Link - Hamilton, W.D. and M. Zuk, 1982. Heritable true fitness and bright birds: A role for parasites. Science, 218: 384-387.
CrossRef - Newton, I., 1991. Habitat variation and population regulation in sparrowhawks. Ibis, 133: 76-88.
CrossRefDirect Link - Wiklund, C.G., 1996. Determinats of dispersal in breeding merlins (Falco columbarius). Ecology, 77: 1920-1927.
CrossRefDirect Link