
Muhammad Akbar Khan
Department of Zoology, University of the Punjab, Lahore, Pakistan (54590)
Muhammad Umer Farooq
Department of Zoology, University of the Punjab, Lahore, Pakistan (54590)
International Journal of Zoological Research
Year: 2006 | Volume: 2 | Issue: 2 | Page No.: 100-109







ABSTRACT
During the Miocene, the Indian plates moved closer to Eurasia causing the further elevation of the Alps, Himalayas and other mountain belts. The Neogene terrestrial rocks generally the Siwaliks, form the Sub-Himalayas. Early pecarons in Africa, Pakistan and India are poorly known. Ruminant fossils are numerous in the Siwaliks. The cervoid are found in Eurasia and presumably evolved there but some fragmentary fossils are found in Bugti hills and two specimens are collected from the Chinji Formation. Tragulids are found in the Lower Miocene of Egypt, East Africa and Nambia and Giraffoids evolved in Africa. The Bovid (Eotragus) appeared in Europe and the same time in the lower Siwalik of Pakistan. Changes in global climate had a direct impact on the distribution of species. The global climate also permitted Asian animals to disperse into Europe. Faunal exchange between Europe and the Siwaliks occurred through SE Europe and Anatolia. While extensive landmasses were slowly being formed out of a number of smaller fragments, sea level changes connected and disconnected these areas and allowed for, or inhibited faunal exchange. The effects of the sea level changes were strongest during the Early Miocene, whereas from the Middle Miocene onward, the land sea distribution seems to have been less affected and less important in controlling the geographical distribution of mammals.
PDF Abstract XML References
How to cite this article
Muhammad Akbar Khan and Muhammad Umer Farooq, 2006. Paleobiogeography of the Siwalik Ruminants. International Journal of Zoological Research, 2: 100-109.
DOI: 10.3923/ijzr.2006.100.109
URL: https://scialert.net/abstract/?doi=ijzr.2006.100.109
DOI: 10.3923/ijzr.2006.100.109
URL: https://scialert.net/abstract/?doi=ijzr.2006.100.109
INTRODUCTION
The Neogene sediments of the Siwalik outcrop belt stretches along the southern foothills of the Himalayas from south central Pakistan through India to Nepal. The Neogene non-marine rocks of the Himalayas and adjacent ranges have generally been ascribed as Siwalik; a term well entrenched in South Asian geologic usages. These sediments are best known for their abundant Neogene mammalian fauna. The best record and most studied region is the Potwar Plateau in northern Pakistan. The Potwar plateau of the Punjab Province (72°30’ E, 33°00’ N) is an elevated area of some 20,000 km2 bounded to the north by the Kala Chitta and Margala Hills, south by the Salt Range, east by the Jhelum River and west by the Indus River. A series of faunas were described from Pakistan, later the names of these faunas were applied to the formations from which they were collected and still later, the names have been applied as stages; Murree, Kamlial, Chinji, Nagri, Dhok Pathan, Soan, most of which have been palaeomagnetically sampled and which detailed range charts. The Siwalik formations have always been cryptic chronostratigraphic units and from the paleontological point of view recognition of the formations and their boundaries has been a crucial step in dating the fossils (Colbert, 1935; Johnson et al., 1985; Tauxe and Opdyke 1982; Tauxe and Badgley, 1988). This practice has in the past produced much confusion and sterile debate, but it is now possible to assume that with the contribution of magnetostratigraphy as a means of dating the rocks this era of confusion is past (Bary et al., 2002). Between 7.5 and 6.5 ma the Siwalik fauna for the first time becomes closely similar to contemporary ones in northern and western Eurasia. This pattern of Eurasian connections contrasts to previous similarities to modern Southeast Asian mammals and indicates a new phase had begun. Extinctions include Sivapithecus, archaic carnivores, Deinotherium, Brachypotherium, dormice and possible tree shrews and lorisids. Appearances include leporids, Hystrix, a giraffine, several bovids and eventually cercopithecids.
As a consequence, the middle and late Miocene Siwalik faunas are a mixture of endemic Asian taxa and immigrant-derived taxa. After 18 million years, there have been several turnover events, some of which are more abrupt and of greater magnitude than others. However, throughout its history and particularly just prior to 7.5 million years, the Siwalik fauna continues to have a strong taxonomic similarity to the modern fauna of tropical Southeast Asia and it is likely that many extant Southeast Asian species have derived from or closely related to Siwalik forms.
The Siwalik faunas had their origin in a major turnover, which occurred prior to 18 million years and possibly as early as 22 million years. That turnover event is manifest by the appearance of many immigrant taxa in the Siwaliks (Bary et al., 1995) and can be approximately correlated to oceanographic events, including a short-term fall in sea-level (Haq et al., 1987). The Paratethys and Tethys became smaller and extensive areas in the Middle East became shallow seas and finally became land. The area of SE Europe became connected to Eurasia, while the collapse of the area north of Crete separated it from Anatolia. The Red Sea formed and Arabia became disconnected from Africa. During much of the Miocene India may have had a contact with Asia at the place of the present Himalayas, but there is little evidence of faunal exchange through this area. The Paratethys was another barrier. Faunal exchange between northern Asia and the southern continents may have occurred through the areas between Himalayas and Paratethys.
While plate tectonics gradually changed the geography, sea level changes were, geologically speaking, instantaneous and had a magnitude of up to a hundred meters and occasionally successive events had accumulative effects. Between 15 and 10.5 ma, sea level dropped in three successive events over two hundred meters. Sea level changes were related to global climate; ice accumulation on Antarctica was responsible for sea level drops (Miller and Mountain, 1996). In the Indian Subcontinent there is virtually no Oligocene and earliest Miocene record. However, in Pakistan most of Bugti fauna extends from the Early Early Oligocene upto the Late/Latest Oligocene (Marivaux et al., 1999; Marivaux et al., 2001; Antoine et al., 2003; Antoine et al., 2004; Marivaux and Welcomme, 2003)
The fossil ruminants are quite abundant in the Neogene of the Siwaliks and the Mediterranean. They are very well known and are represented by a variety of tragulids, cervids, giraffids and bovids. The ruminants are well represented taxonomically and are ecologically diversified, ranging in size from the small tragulid (Dorcatherium minus) to the large giraffid, (Bramatherium). The Miocene faunal turnover events introduced immigrants into South Asia mainly from Africa, whereas the Pliocene events record mammalian faunas closely similar to contemporary ones in northern and western Eurasia (Bary and Flynn, 1989). The latest faunal turnover events during the late Pliocene around 2-9 ma introduced many Eurasian mammals in the Siwalik faunal province (Bary et al., 1982).
Ruminants
The ruminants (infraorder Ruminantia) are a diverse group of mammals, which belong to order Artiodactyla. Artiodactyls contain a large number of taxa falling into three major groups; the Suiformes, Tylopods and Ruminants. Artiodactyls, despite their great diversity, share the same diagnostic leg structure. They have generally 4 digits with the axis of the foot passing between the third and fourth digits and have trilobed upper fourth milk premolar (dp4). The most primitive representative of ruminants had a full complement of teeth but during evolution there has been a strong trend toward reduction of the upper incisors which in most persisting species have been replaced with a horny pad. By both its shape and position, the lower canines often assume the role of lower incisors. The upper canines may be absent or, in some of the most primitive forms, long and dagger- like. The first artiodactyls appear in the early Eocene from condylarth-like ancestors (Janis and Scott, 1987). The ruminant radiation gave rise to six modern families with nearly 200 extant species and it also produced an abundance of extinct species and higher-level taxa. However, most of what we know of this radiation is based on taxa from the temperate Oligocene and Miocene of North America, Europe and central Asia, with little being known of contemporaneous ruminants from tropical southern Asia or Africa, which are now the sites of the greatest species diversity (Bary et al., 2005). This suborder of the artiodactyls reached its zenith late in the Neogene although its origin lies ca. 45 million years ago in the Middle Eocene. Concomitant with their abundance in extant faunas and their wide geographical distribution ruminants show a broad range in ecological specialization: nearly all kinds of available terrestrial habitats from the coast to 6000 meters above the sea level and from tropical rainforests to polar tundras are populated by ruminants. Dietary preferences extend from some omnivory (including carrion) to strictly herbivorous with adaptations to browsing and grazing herbivory. Their natural zoogeographical distribution differs clearly from family to family and excludes Australia and Antarctica; Bovidae inhabit Africa, Eurasia and North America, Cervidae Eurasia, North and South America, Giraffidae Africa, Moschidae Asia, Antilocapridae North America and Tragulidae Asia and Africa. Since the beginning of humankind they have possessed a significant place in our economic and cultural history (Gentry et al., 1999). In the Old World the suborder Ruminantia at the present day comprises the family Tragulidae (chevrotains) plus the infraorder Pecora (Cervidae, Giraffidae and Bovidae to which some would add Moschidae for the musk deer of Asia). The suborder also includes extinct families and the New World Antilocapridae (Gentry and Heizmann, 1996).
Family Tragulidae
Traguloids have occurred in both Eurasia and North America since the Middle Eocene (Vislobokova, 2001). Three groups, archaeomerycids, lophiomerycids and bachitheriids, seem to exist in only Eurasia. Gelocids and tragulids occurred in Eurasia and Africa. In America, traguloids were represented by leptomerycids and gelocids. Hypertraguloids, mainly North American groups, were spread from the late Middle Eocene through the beginning of the Oligocene in both the Old and New Worlds and up to the Early Miocene in North America. The presence of Hypertraguloidea gen. et sp. is reported from the Middle Eocene Khaichin Ula II Fauna from Mongolia (Badamgarav and Reshetov, 1985). In the Late Eocene to the Early Oligocene of Mongolia, hypertraguloids were represented by Praetragulus (Vislobokova, 1998). Although there is no direct evidence on the place of origin of tragulines, Asia seems to be very plausible as the site of their early development. During the Late Cretaceous, Asia was a very large continent with very diverse biotopes. Possible ancestors of artiodactyls had already existed there. The primitive structure and a large diversity of Eocene tragulines in Asia also confirm the Asian origin of this group (Vislobokova, 1997). Many traguline genera occurred in the Middle Eocene of Asia. The most ancient of them were represented by the archaeomerycids Archaeomeryx and Xinjiangmeryx.
Archaeomeryx is known in the Irdinmanhan and Sharamurunian mammal ages of China, which are correlated to most of the Rhenanian (Late Lutetian to Early Bartonian) of Western Europe and to the Uintan of North America (Matthew and Granger, 1925; Tong et al., 1995). This genus was widespread in Central Asia, including Mongolia and Kazakhstan (Gabounia, 1977; Vislobokova, 1990). Xinjiangmeryx is known from only the Sinkiang fauna of the Tufan Basin, China (Zheng, 1978). The four other genera appeared at the end of the Middle Eocene. Among them, the archaeomerycids, Miomeryx and Notomeryx and the lophiomerycid, Lophiomeryx were found in China (Matthew and Granger, 1925; Qiu, 1978) and the lophiomerycid Indomeryx was reported from Burma and China (Qiu, 1978; Pilgrim, 1928). At the Middle-Late Eocene boundary (about 38 ma), the diversity of tragulines in Asia sharply increased: the gelocid Gobiomeryx, the tragulid Krabimeryx and the hypertraguloid Praetragulus spread in Asia (Vislobokova, 2001; Matthew, 1925; Trofimov, 1957; Ducrocq, 1992; Wang, 1992 ).
The extant genus Tragulus is found in low hilly areas of India. The extinct genus Dorcatherium is known from Asia (Lydekker, 1876; Matthew, 1929; Corbet and Hill, 1980), Europe and Africa. Until now many species of the genus Dorcatherium have been reported from Africa, Europe and Asia. The three Pakistani species of the genus Dorcatherium are D. minimus, D. minus and D. majus was founded by Lydekker (1876) on the basis of two upper molars and a maxilla from the Middle Siwaliks of Khushalgarh, district Attock, Punjab, Pakistan. Colbert (1935) has reported a number of the species from Chinji, district Chakwal and from Hasnot, district Jhelum, Punjab, Pakistan. Akhtar (1996) described the fourth upper premolar in the genus Dorcatherium and the species Dorcatherium majus from the Siwaliks. Tragulids are well known from the Siwaliks of Pakistan and reported from the Chinji, Nagri and Dhok Pathan Formations. The Tragulids seem to be appeared in the Chinji Formation. Both genera of the family Tragulidae are reported, which are Dorcatherium and Dorcabune. Two species of the Dorcatherium are known from the Formation, which are, Dorcatherium majus and Dorcatherium minus. Whereas only one species of Dorcabune i.e., Dorcabune anthracotherioides is reported. The Nagri Formation reveals two genera i.e., Dorcatherium and Dorcabune. The genus Dorcatherium is known by one species, which is Dorcatherium majus. The second genus Dorcabune is also known by one species i.e., Dorcabune nagrii. The Dhok Pathan Formation is richly fossilerous and both genera Dorcatherium and Dorcabune are reported from the Formation. The Tragulids entered in Asia and Europe in Miocene but survived to date in Asia where as they extinct in Europe in late Pliocene. In Africa they appeared in Pleistocene to date.
At least three adaptive radiation events preceded the diversity of tragulines in Asia. The two first of them happened earlier than 49 million years ago. The first radiation led to the appearance of the traguloid and hypertraguloid branches. During the second radiation, traguloids were divided into archaeomerycids, lophiomerycids and tragulids. The third radiation was in the late Middle Eocene and gave origin to gelocids and, probably, leptomerycids. At the same time or later, at the Eocene/Oligocene boundary, bachitheriids branched out from archaeomerycids. Hypertraguloids and ancestors of leptomerycids dispersed into North America apparently at the end of the Middle Eocene (not later than 41 million years ago). The most ancient hypertraguloid Simimeryx is known from the Late Uintan of California and the central Great Plains (Stock, 1934; Webb, 1998). The first appearance of the leptomerycids Leptomeryx and Hendryomeryx happened at the Early-Late Uintan boundary (Leidy, 1853; Black, 1978). The adaptive radiation of hypertraguloids, with the appearance of Parvitragulus, Hypertragulus and Hypisodus, occurred at the end of the Middle Eocene at the Late Uintan-Duchesnean boundary (about 39.5 million years ago) (Webb, 1998). During the Oligocene, tragulines developed independently in Asia and America. In the Late Eocene, gelocids penetrated into Europe (Pavlow, 1900), which was almost completely isolated from Asia. The first adaptive radiation of gelocids in Europe evidently occurred in the middle of the Late Eocene (later 35.5 ma) Gelocus and Phaneromeryx are found in France.
Family Giraffidae
Giraffids have a rich fossil history consisting of approximately thirty species throughout the Neogene of the Old World (Bohlin, 1926; Hamilton, 1978; Geraads, 1986; Gentry and Hooker; 1988). The family Giraffidae represents only two living species, the Okapi (Okapi johnstoni) and the Giraffe (Giraffa cameloparadalis) (Solounias et al., 2000). Both the species are present in Africa. The Okapi is present in a limited area of heavy forest in the West Africa, whereas the Giraffe in the more open parts of the Africa below the Sahara. The giraffids started their history in the Early Miocene. They appeared in the Early Miocene sediments of central Asia from where they dispersed to Europe most probably in the Middle Miocene. The first appearance of the giraffids in Africa is reported in the Miocene sediments (Romer, 1974; Gentry, 1993). The giraffids record in South Asia begins from the Early Miocene times, from where they later dispersed to the Southeast Asia. The Giraffids made their appearance in the lower Chinji strata and relatively abundant in quantity (Fig. 1). No larger or Sivatherine giraffid was present in Chinji strata. As indicated by the fossil record, the Sivatherines were definitely present in the Nagri. Since the Nagrian record is highly fragmentary, nothing can be stated with certainty about the quality of the group. Dhok Pathanian was the age of gigantic giraffids.
Seven species of the Sivatherine giraffes are known from the beds of this age. Tatrot, the part of Upper Siwalik in Pakistan is still without any record of the giraffids but the Pinjor, the part of Upper Siwalik in India is characterized by the presence of large giraffids such as Sivatherium. The Giraffa continued to survive till the upper most limits of the Pinjor.
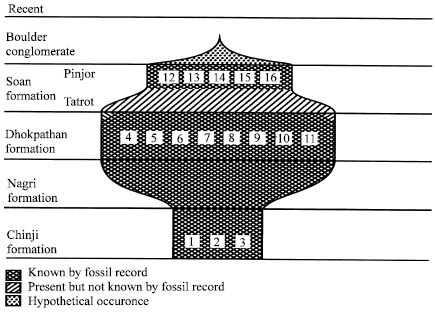 | |
| Fig. 1: | Distribution of the Siwaliks giraffids (1. Bramatherium perimense. 2. Giraffokeryx chinjiensis. 3. Giraffokeryx punjabiensis. 4. Giraffa sivalensis. 5. Giraffa punjabiensis. 6. Giraffa priscilla. 7. Helladotherium grande. 8. Hydaspitherium birmanicum. 9. Hydaspitherium grande. 10. Hydaspitherium megacephalum. 11. Hydaspitherium magnum. 12. Bramatherium geraadsi. 13. Sivatherium giganteum. 14. Vishnutherium iravaticum) |
The presence of Sivatherium and Giraffa just below the Boulder Conglomerates indicates that some members of these genera were probably present when Boulder Conglomerates were accumulating above the Soan Formation. Paleobiogeographically the giraffids are distributed as follows; Late Miocene -Pleistocene in Asia, Late Pliocene-Pleistocene in Europe and Pliocene to date in Africa.
Family Bovidae
Bovids comprise a family of ruminant artiodactyls that are the dominant mammalian herbivores in Old World terrestrial communities. The family is highly diverse, with numerous extant species and has an extensive fossil record with species in Africa, Eurasia and North America. Bovids appear to have had three major adaptive radiations at 14, 7.5 and 2 million years ago (Gentry, 1970; Gentry, 1978; Solounias, 1982; Thomas, 1984; Ye, 1989) but few fossils are known from sediments older than 14 million years ago (Solounias et al., 1995). Citations of very early Bovidae and the great diversity in the primitive forms in Central Asia, Mongolia and China and the sudden appearance of bovids belonging to different subfamilies in Europe, the Indian Subcontinent and Africa, suggest that bovids their origin in north or central Asia. The oldest bovids outside north or central Asia are the 18 ma old boselaphine Eotragus from the Indian Subcontinent (Solounias et al., 1995) and the Eotragus from MN4e in Europe, both probably coming from north or central Asia. Eotragus ranged well beyond Europe. Besides the early Pakistan example, putative Eotragus horn cores have been found in North Africa, Israel, China and East Africa. Protragocerus first appeared in Europe, in the Indian Subcontinent in the Lower Chinji Formation and in Africa. Miotragocerus of Eurasian origin entered some 6 million years ago in Africa (Gentry, 1978) and also very common in the Middle Siwalik of Pakistan. Sivoreas entered the Indian Subcontinent 13.8 ma (Barry and Flynn, 1989) and is found in the Vallesian of Africa. Another middle Miocene bovid, Hypsodontus is very different from Boselaphini, Eotragus, or Tethytragus and indicates an early dichotomy in the Bovidae, or possibly a pecoran parallelism with them. It and some related genera, if validly separate from one another, achieved a broad Old World distribution in the middle Miocene: China, Russia, India, Turkey, Southwestern Asia, South eastern Europe and Africa. In Arabia and China it seems to occur prior to the middle Miocene and perhaps as early as Eotragus does in Pakistan and Europe (Gentry, 1990; Azanza, 1994). Later Miocene bovids fall into several subfamily and tribal groups and are said to be modern bovids. The Bovinae contain Boselaphini (living Indian nilgai) and Bovini (cattle) and their relatives. The Antilopinae contain Gazella, its relatives and the spiral-horned Antilope of India. The Caprinae contain Caprini (sheep and goats), Ovibovini and diverse related forms. The earliest known boselaphine, if Eotragus be excluded, may come from Pakistan (Solounias et al., 1995) at 17.6 ma. In Europe boselaphines only appeared in the later middle Miocene. Miocene genera of boselaphines nearly all had keels on their horn cores. Around the end of the upper Miocene Tragoportax vanished. In its place Bovini appeared in Eurasia, especially India and in Africa.
The Neotragini are a probably paraphyletic African tribe (Gentry, 1992). The middle Miocene Homoiodorcus sp. from Al Jadidah, Saudi Arabia (Thomas, 1984) is more primitive and less obiviously neotragine than the much later type species H. tugenicum in Kenya. Homoiodorcus is probably a close relative of antilopini. There are early citations of the antilopine Gazella from Africa (Set IIIa, IV), the Middle Chinji Formation in Pakistan and possibly from MN6, but certainly MN9 in Anatolia (Gentry, 1978; Thomas, 1984; Hamilton, 1973; Kohler, 1987). Miocene gazelles differ from living ones by their generally smaller size, longer nasals, lower teeth, longer premolar rows and little or no enlargement of the rear lobe of m3. They are present in Spain, France, Greece, Ukraine and Georgia from the Turolian. In the European upper Miocene Caprinae are represented by Palaeoryx, Protoryx, Pachytragus, Pseudotragus, Aragoral, Norbertia and perhaps Procobus but in the Siwaliks it is not known from Pakistan but three species (Sivacapra sivalensis, Sivacapra crassicornis, Pantholops hundesiensis) of it are known from Pinjor (India) (Pilgrim, 1939). Near the end of the Miocene and shortly afterwards modern tribes of African antelopes became recognizable in that continent and in Pakistan. Some bovids of probable African origin spread into southern Europe very late in the Miocene (Seguenza, 1902).
Family Cervidae
Cervids appeared in Europe in the late Early Miocene (MN3) with small size and lack of antlers but did not reach the Siwaliks during the Miocene. During MN3 and MN4 the genus Procervulus became common in Europe evolving to a medium sized cervid with few changes in dental morphology (Gentry and Heizmann, 1999). Early small cervids, e.g., Eumeryx and Iberomeryx, appeared in the Middle Oligocene sediments of Central Asia from where they dispersed to Europe and North America, most probably, in the early Miocene. The first appearance of cervids in South America and Africa has been reported from the Pleistocene (Romer, 1974). Cervids entered in the Siwaliks during Plio - Pleistocene time and they showed the similarity in fossils record of Pliocene deer in Europe. In the Siwaliks there are no deer in pre-Hipparion levels but now recently a specimen of Cervus sivlensis is collected from the Chinji Formation of the Middle Siwalik (Ghaffar et al., 2005). Late Oligocene and early Miocene indeterminate large cervoid (fragmentary mandible) is found from the Zinda Pir sequences in Pakistan (Barry et al., 2005). Several species of the family Cervidae have been described mainly from the Upper Siwalik rocks of the Western Himalayas including the Siwalik Hills and adjoining ranges in India and southern Kashmir, Potwar and Trans-Indus Hill ranges of Pakistan.
REFERENCES
- Antoine, P.O., S.M.I. Shah, I.U. Cheema, J.Y. Crochet and D. De Franceschi et al., 2004. New remains of the baluchithere Paraceratherium bugtiense (Pilgrim, 1910) from the late/latest oligocene of the Bugti hills, Balochistan, Pakistan. J. Asian Earth Sci., 24: 71-77.
CrossRefDirect Link - Barry, J., M. Morgan, L. Flynn, D. Pilbeam and A.K. Behrensmeyer et al., 2002. Faunal and environmental change in the Late Miocene Siwaliks of Northern Pakistan. Paleobiology, 28: 1-71.
CrossRefDirect Link - Barry, J.C., S. Cote, L. MacLatchy, E.H. Lindsay, R. Kityo and A.R. Rajpar, 2005. Oligocene and early miocene ruminants (Mammalia, Artiodactyla) from Pakistan and Uganda. Palaeontol. Elect., 8: 1-29.
Direct Link - Colbert, E.H., 1935. Siwalik mammals in the American museum of natural history. Trans. Am. Philosophical Soc., 26: 1-401.
Direct Link - Corbet, G.B. and J.E. Hill, 1980. A World List of Mammalian Species. 2nd Edn., Facts on File Publications, British Museum, Natural History, New York, London, ISBN-10: 0816015481.
Direct Link - Hamilton, W.R., 1973. The lower Miocene ruminants of Gebel Zelten, Libya. Bull. British Museum (Natural History) Lond. Geol., 21: 73-150.
Direct Link - Hamilton, W.R., 1978. Fossil giraffes from the Miocene of Africa and a revision of the Giraffoidea. Phylosophical Trans. Royal Soc. London, 283: 165-229.
Direct Link - Haq, B.U., J. Hardenbol and P.R. Vail, 1987. Chronology of fluctuating sea levels since the triassic. Science, 235: 1156-1167.
CrossRefDirect Link - Janis, C.M. and K.M. Scott, 1987. The interrelationships of higher ruminant families with special reference on the members of the Cervoidea. Am. Mus. Novitates, 2893: 80-85.
Direct Link - Johnson, N.M., J. Stix, L. Tauxe, P.F. Cerveny and R.A.K. Tahirkheli, 1985. Paleomagnetic chronology, fluvial processes and tectonic implications of the Siwalik deposits near Chinji village, Pakistan. Geology, 93: 27-40.
Direct Link - Miller, K.G. and G.S. Mountain, 1996. The leg 150 shipboard party and members of the New Jersey coastal plain drilling project 1996: Drilling and dating New Jersey oligocene-miocene sequences: Ice volume, global sea level and exxon records. Science, 271: 1092-1095.
CrossRefDirect Link - Marivaux, L., J.L. Welcomme, P.O. Antoine, G. Metais and M. Benammi et al ., 2001. A fossil lemur from the Oligocene of Pakistan. Science, 294: 587-591.
CrossRefDirect Link - Marivaux, L. and J.L. Welcomme, 2003. Diatomyid and baluchimyine rodents from the oligocene of Pakistan (Bugti Hills Balochistan): Biochronologic and paleobiogeographic implications. J. Vertebrate Paleontol., 23: 420-434.
CrossRefDirect Link - Pilgrim, G.E., 1928. The artiodactyla of the eocene of Burma. Palaeontol. Ind., N. Ser., 13: 1-39.
Direct Link - Solounias, N., J.C. Barry, R.L. Bernor, E.H. Lindsay and S.M. Raza, 1995. The oldest bovid from the siwaliks, Pakistan. J. Vertebrate Paleontol., 15: 806-814.
Direct Link - Stock, C., 1934. A hypertragulid from the sespe uppermost eocene, California. Proc. Natl. Acad. Sci., 20: 625-629.
PubMedDirect Link - Tauxe, L. and C. Badgley, 1988. Stratigraphy and remanence acquisition of a paleomagnetic reversal in alluvial Siwalik rocks of Pakistan. Sedimentology, 35: 697-715.
CrossRef