
Mohsen Abdel- Tawwab
Department of Fish Ecology, Suez Canal University, Ismailia, Egypt
Mamdouh A. A. Mousa
Department of Fish Ecology, Suez Canal University, Ismailia, Egypt
Safaa M. Sharaf
Department of Animal Production, Faculty of Agriculture, Suez Canal University, Ismailia, Egypt
Mohammad H. Ahmad
Department of Fish Niltrition, Central Laboratory for Aquaculture Research, Abbassa, Abo-Hammad, Sharqia, Egypt
International Journal of Zoological Research
Year: 2005 | Volume: 1 | Issue: 1 | Page No.: 41-47







ABSTRACT
This study was carried out to evaluate the response of Nile tilapia, Oreochromis niloticus (L.) Fed dietary protein levels to chronic stress induced by doubling fish density. In this study, Nile tilapia (15±1 g) was randomly distributed into the aquaria at a rate of 15 or 30 fish/100 L. The temperature was adjusted at 27±1°C. Fish of each density were fed either a diet containing 25, 35 or 45% Crude Protein (CP) with a feeding rate of 3% of life body weight twice daily for two weeks. The results of this study showed that Hb and Ht were slightly reduced and no significant effect of protein level on both of them. The RBCs count in chronically stressed fish by high fish density showed such elevation and was increased significantly with increasing protein level in the diet. Glucose level in plasma was increased significantly due to high fish density especially at 25 and 35% CP. No marked changes in glucose level were observed at both fish densities at 45% CP. Protein level in the diet significantly induced plasma glucose and protein especially at low fish density. Chronic stress induced by high fish density insignificantly affected protein level in plasma. Similarly, a decrease in plasma cortisol levels due to high densities and high dietary protein levels was verified. The aspartate amninotransferase (AST) activity in liver was insignificantly reduced at high fish density and significantly increased by increasing protein level in the diet. AST activity in plasma was only significantly reduced at 45% CP, however, AST activity in fish muscle is not significantly affected by either fish density or protein level in the diet. Alanine aminotransferase (ALT) activity in plasma, liver and muscle was insignificantly affected by fish density or protein level. It could be concluded that Nile tilapia may quickly adapted to high rearing density by enhancing feed quality especially protein level in the diet to prevent the deleterious effect in fish farm.
PDF Abstract XML References
How to cite this article
Mohsen Abdel- Tawwab, Mamdouh A. A. Mousa, Safaa M. Sharaf and Mohammad H. Ahmad, 2005. Effect of Crowding Stress on Some Physiological Functions of Nile Tilapia, Oreochromis niloticus (L.) Fed Different Dietary Protein Levels. International Journal of Zoological Research, 1: 41-47.
DOI: 10.3923/ijzr.2005.41.47
URL: https://scialert.net/abstract/?doi=ijzr.2005.41.47
DOI: 10.3923/ijzr.2005.41.47
URL: https://scialert.net/abstract/?doi=ijzr.2005.41.47
INTRODUCTION
Nile tilapia, a native fish species of Egypt, grows faster in warm months. So, tilapia culturing in Egypt has become more popular because it is relatively easy in a variety of aquaculture systems and because tilapia are favorable food fishes. The maximum growth of tilapia depends upon the dietary protein quality, energy content of the diet, the physiological state of the fish, age, reproductive state and the environmental factors such as temperature, salinity etc.[1]
In aquaculture, control of fish size and production are two important tasks to meet the market demands and increasing the stocking density is a way of dealing with problem of land shortage. In many cultivated fish species, growth is inversely related to stocking density and this is mainly attributed to social interactions[2-4]. The social interactions through competition for food and/or space represent a type of chronic stressor that could negatively affect fish growth. On the other hand, fish in intensive rearing facilities are continuously exposed to management practices such as handling, transportation or confinement, which are often confined in high fish densities[5]. These practices are potential stressors to the fish[6].
Fishes respond to stress throughout many physiological changes to maintain homeostasis after stress including hematology[7], osmolaty[8], hormone release and energy metabolism[9,10]. The hematological and biochemical examinations of intensively farmed fish are an integral part of evaluating their health status. However, the diet composition, metabolic adaptations and variations in fish activity are the main factors responsible for the seasonal changes in hematological parameters of fish[11,12]. The alteration of blood biochemistry and hormones status might be indicative of unsuitable environmental conditions or the presence of stressing factors as toxic chemicals, excess of organic compounds, crowding and even usual procedures in aquaculture[13]. Thus, determining the basal parameters of blood biochemistry and hormones might be of great importance in order to monitor the health status for commercial purposes.
Crowding is a common husbandry practice in aquaculture, as is reducing the water level or increasing the fish stocking density. By establishing the relationship between dietary protein level and crowding stress, enhanced monitoring of fish stocks and prediction of their physiological needs may be possible. These situations are of obvious concern within the aquaculture industry, as they often related to the need for decreasing mortality, increasing feed efficiency and minimizing food wastes and the concomitant environmental pollution[14]. Therefore, this study was carried out to evaluate the physiological changes of Nile tilapia, Oreochromis niloticus (L.) subjected to different stocking densities, which represent usual stress in aquaculture practice and to which extent the protein nutrition may reduce this effect.
MATERIALS AND METHODS
Experimental procedures: Healthy fish of Nile tilapia, Oreochromis niloticus (L.) were collected from the nursery ponds of Central Laboratory for Aquaculture Research, Abbassa, Abo-Hammad, Sharqia. Fish (15±1 g) were acclimated indoor tanks for 2 weeks to laboratory conditions. The fish of mixed sex were distributed randomly in glass aquaria (130 L capacity containing 100 L of aerated water) with either 15 and 30 fish/aquarium. Each aquarium was supplied with compressed air via air-stones from air pumps. The temperature was adjusted at 27±1°C by using thermostatically controlled heaters. Half of the aquaria water was siphoned every day for excreta removing and an equal volume of well-aerated water replaced it. Dead fish were removed and recorded daily.
A semi-moist diet was prepared from purified ingredients and was used to formulate three identical diets in all the nutrient contents except for the protein levels (Table 1). The formulated diets contained 25, 35 or 45 crude protein. Fish were fed frequently to satiation level three times daily one week before the experiment start for food adaptation. Then, the experiment was conducted for two weeks after which fish harvested, counted and weighed. Three aquaria were assigned for each treatment in each fish stocking density. During the experiment, fish fed one of the tested diets at a rate of 3% of live body weight twice daily. The used diets were analyzed using standard methods of the Association of Official Analytical Chemists[15] for determination of moisture, crude protein, total lipids and ash.
Preparation of blood samples: Fish were not fed for 24 h before sampling. Fish were anaesthetized with buffered MS222 (50 mg L-1) and blood was collected with a hypodermic syringe from the caudal vein. The blood collection lasted less than 3 min in order to avoid cortisol rise induced by the manipulation during sampling. The extracted blood was divided in two sets of eppendorf tubes. One set contained heparin, used as anticoagulant, for hematology (hemoglobin, haematocrit and red blood cells counting). The second set, without anticoagulant, was left to clot at 4°C and centrifuged at 5000 rpm for 10 min at room temperature. The collected serum was stored at –20°C for further assays (glucose and cortisol). After decapitation of fish, samples of liver and muscle were taken and frozen for further biochemical analysis.
Physiological measurements: Haematocrit values (Ht %) were immediately determined after sampling by placing fresh blood in glass capillary tubes and centrifuging for 5 min in a microhematocrit centrifuge. Hemoglobin levels (Hb, g dL-1) were determined colorimetrically by measuring the formation of cyanomethaemoglobin after using a commercial kit. Red Blood Cells (RBCs, cells μL-1) were counted under the light microscope using a Neubauer haemocytometer after blood dilution with phosphate-buffered saline.
Glucose was determined colorimetrically using glucose kits according to Trinder[16]. Total protein content in plasma, muscle and liver was determined colorimetrically according to Henry[17]. Activities of aspartate aminotransferase (AST) and alanine aminotransferase (ALT) in plasma, liver and muscle were determined colorimetrically according to Reitman and Frankel[18]. Kits reagents used for these measurements are supplied by Egyptian American Co. for Laboratory Services, Egypt.
Serum cortisol levels were measured by radioimmunoassay with a commercially available 125I and the radioactivity was quantified using a liquid scintillation counter as previously validated by Barcellos et al.[19].
Statistical analysis: The obtained data were subjected to in two-way ANOVA and the differences between means probability level using Duncan’s New Multiple Range Test.
| Table 1: | Ingredients and chemical composition and the experimental diet (on DM basis) |
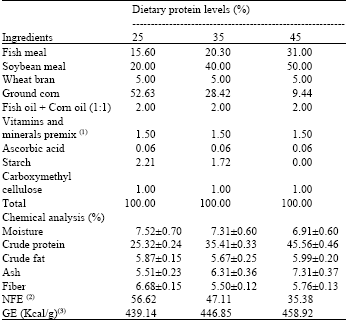 | |
| [1]Vitamin and minerals premix: each 2.5 kg contain vitamin A 12 MIU; D3 2 MI U, E 10 g; K 2 g; B1 1 g; B2 4 g; B6 1.5 g; B12 10 mg; Pantothenic acid 10 g; Nicotinic acid 20 g; Folic acid 1 g; Biotin 50 mg; Choline chloride 500 mg; copper 10 g; iodine 1 g; iron 30 g; manganese 55 g; zinc 55 g and selenium 0.1 g (2)NFE (Nitrogen Free Extract) = 100–(protein + lipid + ash + fiber) (3)GE (Gross Energy) was calculated as 5.64, 9.44 and 4.11 Kcal g-1 for protein, lipid and NFE, respectively | |
The software SPSS, version 10 (SPSS, Richmond, USA) was used as described by Dytham[20].
RESULTS
At high fish density, groups showed no elevation in the RBCs counting, while it was significantly affected with increasing protein level in the diet (p<0.05). Besides RBCs count increased and Ht decreased insignificantly (Table 2). At high fish density, blood parameters (Hb and Ht) were not reduced significantly, further no significant effect of protein level was observed (Table 2). Glucose level in plasma was significantly increased due to high fish density at 25 and 35% CP (Table 2). No significant changes in glucose level were observed between fish densities at 45% CP. On the other hand, protein level in the diet significantly induced plasma glucose at low fish density, while at high fish density, no changes of glucose level was observed due to the increment in protein level in the diet. Protein level in the diet significantly induced the level of plasma protein (p<0.05), while high fish density insignificantly affected protein level in plasma (Table 2). Plasma protein in this study ranged from 2.07 to 3.58 g dL-1.
| Table 2: | Changes in RBC, Hb, Ht, plasma glucose and plasma protein of Nile tilapia fed different protein levels at two stocking densities |
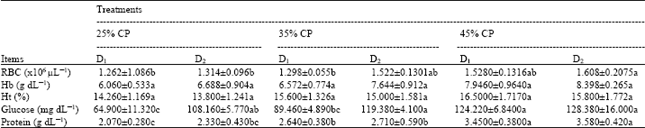 | |
| Table 3: | Changes in AST activity in plasma, liver and muscles of Nile tilapia fed different protein levels at two stocking densities |
 | |
| Table 4: | Changes in ALT activity in plasma, liver and muscles of Nile tilapia fed different protein levels at two stocking densities |
 | |
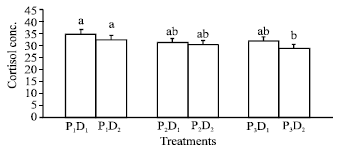 | |
| Fig. 1: | Changes in plasma cortisol level (ng mL-1) of Nile tilapia fed different protein levels at two stocking densities. Bars assigned with the same small letter(s) is not significantly differed at p<0.05 D1 = 15 fish/aquarium, D2 = 30 fish/aquarium P1 = 25% CP, P2 = 35% CP, P3 = 45% CP |
A significant decrease in plasma cortisol levels was observed at the two high protein levels, but it decreased insignificantly due to the high stocking density (p<0.05; Fig. 1). The AST activity in plasma was only significantly increased at 45% CP and insignificantly changed at 25 and 35% CP. AST activity in liver was insignificantly reduced at high fish density and significantly increased by increasing protein level in the diet. Moreover, AST activity in fish muscle is insignificantly affected by either fish density or protein level in the diet (Table 3). The obtained data revealed that ALT activities in plasma, liver and muscle were insignificantly altered due to either fish density or protein level in the diet (Table 4).
DISCUSSION
The blood parameters of teleost fish may be affected by acute and/or chronic stress[6,10]. In this regard, Hb, Ht and RBC values of Nile tilapia were insignificantly increased by doubling fish density in limited water volume and space. This could be related to the low level of activity of the fish in this experiment. Moreover, any chronic stress induced by high fish density here may be limited and it is supposed that if the fish density increased over that used here and/or the fish growth was faster, the stress effect will be higher. McFarlane et al.[21] found that fish stress via crowding by decreasing water level for one minute had no influence on growth, but did induce obvious changes in swimming behavior. This result is due to the short stress time while the stress time here is longer. It is supposed that if the stress time is longer the obvious growth reduction will be obtained especially the higher fish density did lead to higher swimming activity[22] that spent more energy leading to reduced growth. Schippers et al.[23] reported that time has a positive effect on the physiological functions as a result of crowding-related social stress due to increase in biomass with time.
There is an increase in plasma glucose levels induced by high fish density (p<0.05). The mobilization of glucose in response to stress is generally accepted as a means of providing extra energy resources, enabling the fish to overcome the disturbance[9,24]. Alterations in glucose metabolism are a common response to stress by capture in fish[10] including other species such as the golden perch, M. ambigue[25]. On the other hand, the direct effect of the higher metabolic demands during stress is driven by an increase in oxygen transport efficiency by elevation of the haematocrit and red cell numbers[26].
Plasma protein in this study reflected the protein feeding levels. Similar results were obtained by Verdegem et al.[27]. Working with freshwater-reared rainbow trout, Schippers et al.[23] and Zeitoun et al.[28] did not find significant differences in total plasma protein levels in fish fed different protein levels. Plasma proteins contribute significantly to the maintenance of the blood volume and the water content of the fluid in tissue[29,30]. The colloidal plasma protein can not diffuse through capillary membranes towards the surrounding, relatively protein-free tissues. In this way, they exert an osmotic pressure allowing a minimum liquid volume to be maintained in the capillary blood vessels[27].
Concerning the plasma cortisol levels, the obtained results indicated that the dietary protein level inhibited the plasma cortisol level at high density, meanwhile high stocking density has no effect on the cortisol plasma level. This result may be due to that the nature of the chronic stressor can play an important role on how the fish may respond to an additional disturbance. Tripathi and Verma[31] reported that cortisol induces catabolic and aerobic but inhibits anabolic and anaerobic processes in freshwater catfish (Clarias batrachus). They also found an inverse relationship between cortisol level and protein synthesis capacity in fish. Also, Gaslin and Auperin[32] suggested that the particular activation of pre-existing proteins, govern cortisol production.
Exposure of fish to chronic stressors such as toxicants[33] and parasites[34], was not always elevating basal plasma cortisol levels. Of the relatively few studies that have examined the influence of chronic crowding stress on the response to an acute stressor, most have found no effect or a reduction in post-stress cortisol elevations. Kebus et al.[35] and Rotllant and Tort[36] found no effect of increased stocking density on the cortisol response of rainbow trout, Oncorhynchus mykiss (Walbaum) and gilthead seabream Sparus aurata (L.) after rearing periods of 8 days and 3 weeks. Similarly, Procarione et al.[37] observed no difference in post-stress cortisol levels in rainbow trout after 3, 10 and 17 days of crowding. Also, Urbinati et al.[38] found that the concentration of plasma cortisol in matrinxã (Brycon cephalus) decreased inversely to the densities. On the other hand, Ruane et al.[39] observed a transient elevation in plasma cortisol levels in the higher density of common carp Cyprinus carpio (L.) after 15 h before values returned to control levels after 39 h. The lack of an effect on the acute stress response in the previous studies may be due to an acclimation of the internal tissue to chronic stress over time[40].
The cortisol response seen in Ruane et al.[39] is most likely due to the sudden increase in density, as water quality did not deteriorate in the crowded tanks during the experiment and there was no increased competition for food as the fish numbers stayed the same. The plasma cortisol level in this study is less than previously reported for chronically crowded carp[39,41]. This difference may be due to the difference in fish species and the degree of adaptation to the stressor. Previous work on the mechanisms of cortisol release in the sea bass demonstrated a continuous production of cortisol under control conditions but lower reactivity to adrenocorticotropin hormone (ACTH) pulses[42]. In this regard, when a chronic stressor by high density was applied for the yellow perch Perca flavescens L., cortisol levels increased to 46 ng mL-1 after 3 days and 20 ng mL-1 after 2 weeks[43].
AST activities in muscle and liver were higher than that of plasma and ALT activity in liver was higher than those of muscles and plasma. Both enzymes belong to the non-plasma-specific enzymes, which are located within tissue cells and have unknown physiological function in plasma[44]. In rainbow trout, Gaudet et al.[45] showed that AST was concentrated in the heart and ALT in the liver and in the kidney. In English sole, Parophrys vetulus Girard, Casillas et al.[46] found that liver has been shown to contain high AST activity and the liver and kidney are the potential source of ALT. The increase in AST activity in this study is probably related to high activity of enzyme-producing hepatocytes and slightly released to the fish blood. When cell membranes are intact, these enzymes are present in serum in low concentrations, as the cell membranes are impermeable to enzymes when the cells are metabolizing normally [47].
In conclusion, the overall results presented here indicate that Nile tilapia may quickly adapted to high rearing density, however, the high density could not impair the health status of Nile tilapia. It also indicates to the importance of the corrected procedure of fish culture stock by enhancing feed quality especially protein level in the diet to prevent the deleterious effects in farmed fish.
REFERENCES
- Canario, A.V.M., J. Condeca, D.M. Power and P.M. Ingleton, 1998. The effect of stocking density on growth in gilthead sea-bream, Sparus aurata (L.). Aquac. Res., 29: 177-181.
Direct Link - Bonga, S.E.W., 1997. The stress response in fish. Physiol. Rev., 77: 591-625.
CrossRefPubMedDirect Link - Dethloff, G.M., D. Schlenk, S. Khan and H.C. Bailey, 1999. The effects of copper on blood and biochemical parameters of rainbow trout (Oncorhynchus mykiss). Arch. Environ. Contam. Toxicol., 36: 415-423.
Direct Link - Carragher, J.F. and C.M. Rees, 1994. Primary and secondary stress responses in golden perch, Macquaria ambigua. Comp. Biochem. Physiol. Part A: Physiol., 107: 49-56.
CrossRef - Barton, B.A. and G.K. Iwama, 1991. Physiological changes in fish from stress in aquaculture with emphasis on the response and effects of corticosteroids. Ann. Rev. Fish Dis., 1: 3-26.
CrossRefDirect Link - Rehulka, J., B. Minaiik and E. Rehulkova, 2004. Red blood cell indices of rainbow trout Oncorhynchus mykiss (Walbaum) in aquaculture. Aquacult. Res., 35: 529-546.
CrossRefDirect Link - Barcellos, L.J.G., L.C. Kreutz, C. de Souza, L.B. Rodrigues and I. Fioreze et al., 2004. Hematological changes in jundia (Rhamida quelen Quoy and Gaimard Pimelodidae) after acute and chronic stress caused by usual aquacultural management, with emphasis on immunosuppressive effects. Aquaculture, 237: 229-236.
- Reitman, S. and S. Frankel, 1957. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol., 28: 56-63.
CrossRefPubMedDirect Link - Arends, R.J., J.M. Mancera, J.L. Munoz, S.E. Wendelaar Bonga and G. Flik, 1999. The stress response of the gilthead sea bream (Sparus aurata L.) to air exposure and confinement. J. Endocrinol., 163: 149-157.
Direct Link - Verdegem, M.C.J., A.D. Hilbrands and J.H. Boon, 1997. Influence of salinity and dietary composition on blood parameter values of hybrid red tilapia, Oreochromis niloticus (Linnaeus) x O. mossambicus (peters). Aquacult. Res., 28: 453-459.
CrossRef - Tripathi, G. and P. Verma, 2003. Pathway-specific response to cortisol in the metabolism of catfish. Comp. Biochem. Physiol. Part B: Biochem. Mol. Biol., 136: 463-471.
CrossRefPubMedDirect Link - Kebus, M.J., M.T. Collins, M.S. Brownfield, C.H. Amundson, T.B. Kayes and J.A. Malison, 1992. Effects of rearing density on the stress response and growth of rainbow trout. J. Aquat. Anim. Health, 4: 1-6.
CrossRefDirect Link - Urbinati, E., J. Abreu, A. Camargo and M. Parra, 2004. Loading and transport stress of juvenile matrinxa (Brycon cephalus, Characidae) at various densities. Aquaculture, 229: 389-400.
CrossRefDirect Link - Ruane, N.M., E.C. Carballo and J. Komen, 2002. Increased stocking density influences the acute physiological stress response of common carp Cyprinus carpio (L.). Aquacult. Res., 33: 777-784.
CrossRefDirect Link - Gaudet, M., J.G. Racicot and C. Leray, 1975. Enzyme activities of plasma and selected tissues in rainbow trout Salmo gairdneri Richardson. J. Fish Biol., 7: 505-512.
CrossRef - Sandnes, K., O. Lie and R. Waagbo, 1988. Normal ranges of some blood chemistry parameters in adult farmed Atlantic salmon, Salmo salar. J. Fish Biol., 32: 129-136.
CrossRef - Geslin, M. and B. Auperin, 2004. Relationship between changes in mRNAs of the genes encoding steroidogenic acute regulatory protein and P450 cholesterol side chain cleavage in head kidney and plasma levels of cortisol in response to different kinds of acutestress in the rainbow trout (Oncorhynchus mykiss). Gen. Comp. Endocrinol., 135: 70-80.
Direct Link