
Savaas Umar Mohammed Riyaz
Plant Molecular Virology and Nanobiotechnology Laboratory, Department of Biotechnology, University of Madras,
Guindy, 600025, Chennai, Tamil Nadu, India
LiveDNA: 91.14710
Denison Michael Immanuel Jesse
Plant Molecular Virology and Nanobiotechnology Laboratory, Department of Biotechnology, University of Madras,
Guindy, 600025, Chennai, Tamil Nadu, India
Sundarajan Deepan
Plant Molecular Virology and Nanobiotechnology Laboratory, Department of Biotechnology, University of Madras,
Guindy, 600025, Chennai, Tamil Nadu, India
Thangavelu Raja Muthuramalingam
Plant Molecular Virology and Nanobiotechnology Laboratory, Department of Biotechnology, University of Madras,
Guindy, 600025, Chennai, Tamil Nadu, India
Ganapathy Rajendiran
Plant Molecular Virology and Nanobiotechnology Laboratory, Department of Biotechnology, University of Madras,
Guindy, 600025, Chennai, Tamil Nadu, India
Krishnan Kathiravan
Plant Molecular Virology and Nanobiotechnology Laboratory, Department of Biotechnology, University of Madras,
Guindy, 600025, Chennai, Tamil Nadu, India
LiveDNA: 91.12205
International Journal of Virology
Year: 2018 | Volume: 14 | Issue: 1 | Page No.: 1-12







ABSTRACT
Background and Objective: Whitefly-transmitted begomoviruses are the most important limiting factor in ash gourd cultivation in India. This work enlightened about the adaptability and evolution of new begomovirus complexes by interactions between begomoviruses and satellites by mixed infection and recombination. Materials and Methods: Full length genome of bipartite Squash leaf curl China virus (SLCCNV) was amplified by RCA, subsequently its cloned in pBSKII+ vector and sequenced, with universal beta-satellite primers. Croton yellow vein mosaic beta-satellite (CroYVMB), β-satellite and recombination analysis was done through RDP4. Results: Benincasa hispida was a new host for begomovirus, the presence of two different begomovirus species was identified as SLCCNV isolate KP1 and CroYVMB in B. hispida infection was observed. Recombination analysis would serve as a possible evidence for interspecies recombination between the three begomoviral entities-ToLCNDV, SLCCNV and CroYVMB that had been suspected for SLCCNV-KP1infection in the collected B. hispida. Conclusion: Novel recombination was found in DNA-A and DNA β while DNA B was not recombinant; which helped to reduce the host range and severity of the infestation. This frequency of emerging new SLCCNV strains in ash gourd indicating that the virus species had undergone recombination.
PDF Abstract XML References Citation
Received: September 08, 2018;
Accepted: October 22, 2018;
Published: December 01, 2018
How to cite this article
Savaas Umar Mohammed Riyaz, Denison Michael Immanuel Jesse, Sundarajan Deepan, Thangavelu Raja Muthuramalingam, Ganapathy Rajendiran and Krishnan Kathiravan, 2018. Natural Infection and Recombination Analysis of Bipartite Begomovirus and its Cognate Beta-satellite in Benincasa hispida. International Journal of Virology, 14: 1-12.
DOI: 10.3923/ijv.2018.1.12
URL: https://scialert.net/abstract/?doi=ijv.2018.1.12
DOI: 10.3923/ijv.2018.1.12
URL: https://scialert.net/abstract/?doi=ijv.2018.1.12
INTRODUCTION
Gemini viruses (Geminiviridae) possess a circular, single-stranded DNA genome encapsulated within the twinned quasi-isometric geminate icosahedral virions 18×30 nm size. Geminiviruses caused great loss in major crops worldwide which in turn abased the economy of the country1,2. It is classified into seven genera viz., Mastrevirus, Topocuvirus, Curtovirus, Begomovirus, Becurtovirus, Eragrovirus and Turncurtovirus3,4. The genus Begomovirus (type species: Bean golden mosaic virus) is the largest family among the Geminiviridae. Begomoviruses are grouped into Old World (OW) Begomovirus [Europe, Africa, Asia and Australia] and New World (NW) Begomovirus [America] based on genetic diversity and the geographical distribution5. These viruses tend to infect a wide range of eudicot plants and was transmitted by whitefly-Bemisia tabaci (Hemiptera: Aleyrodidae). These viruses have single-stranded, circular DNA genome, of ∼2800 nt in size, encapsidated in twinned icosahedra, transmitted by ubiquitous whitefly and are associated with satellite molecules referred to as alpha and beta-satellite6.
Begomo viruses have been grouped into monopartite (with one circular single stranded (ss) DNA component; DNA-A) and bipartite (with two circular ssDNA components; DNA-A and DNA-B). DNA-A components possess seven open reading frames (ORFs): AV1, AV2, AV3, AC1, AC2, AC3 and AC4. The DNA-B encodes two ORFs: BC1and BV1. The CR possesses the following components: sequence repeats (Iterons), TATA box and conserved hairpin loop structure, which having a nonanucleotide sequence, TAATATT↓AC, the specific site for the binding of Rep protein and initiation of replication7. Both DNA-A and DNA-B components are mandatory for systemic infection in plants. DNA-A maintains the replication and transcription functions, whereas DNA-B maintains the cell to cell movement of viruses.
In addition to DNA-A and DNA-B components in OW begomo viruses have another satellite molecule which is called as beta satellites; it involved to determination of pathogenicity in plants, interspecies recombination is significant in the diversity of geminiviruses8. Mutation, recombination, reassortment and de novo gene acquisition are the important factors for determine of virus diversity. The genetic variation has been known to occur viz., within species, within genera and within family9-11. Molecular events are natural recombinations, homologous and non-homologous recombinations, point mutations are involving a small insertions or deletions of nucleotide which is main source of diversity12. A new Tomato Apical Leaf Curl Virus (ToALCV): a novel, monopartite geminivirus detected in tomatoes in Argentina, ToALCV genome appears to have “modular organization” supported by its recombination origin13 .
Therefore, there is a possibility of intra-species recombination in gemini viruses between their DNA-A and DNA-B, also inter-species recombination between the DNA-A of one species and beta-satellite component from another species. Beta-satellites are pathogenicity determinant molecules that are strictly dependent on the helper virus for its replication, encapsidation and transmission14. Based on the sequence information, the beta-satellites shared 81-86% identity with Croton yellow vein mosaic beta satellite-Bangalore; CroYVMB-[Bang-Cr1] species. Croton yellow vein mosaic virus (CYVMV) has been known to occur with yellow vein mosaic disease in a predominantly growing weed Croton bonplandianum since 196314-17.
This study provides certain valuable information about the host range of virus and the evolution of virus. In view of the above, the present investigation was undertaken with the following objectives on comprehensive and systematic study on characterization of SLCCNV by full genome sequence analysis. The complete nucleotide sequence, the genome organization and the recombination events taken place between distinct bipartite begomo viruses and with another beta-satellite component.
MATERIALS AND METHODS
Isolation of viral DNA from infected ash gourd leaves: Symptomatic leaves of ash gourd plants were collected during December, 2012 to March, 2013 from three different fields of Perambalur district of Tamil Nadu. The plants showing symptoms such as severe interveinal chlorosis, upward leaf curling, shortened internodes, leaf thickening, shortening and growth stunting were considered for plant sample collection. Total DNA extraction was done by Dellaporta et al.18. The extracted DNA was suspended in sterile distilled water and stored at -20°C.
PCR amplification for detection of DNA-A and DNA-B by specific primers: The DNA extracted from the symptomatic ash gourd plants were screened for infection using Polymerase Chain Reaction (PCR) with the red dye master mix commercially obtained from Ampliqons. Degenerate primer pairs (SLCCNVFP/SLCCNVRP) that is specific to partial CP region (~500 bp) was used for its amplification. The PCR was carried out in a Peltier thermal cycler L196GGD (Lark Innovative Technologies, Pvt., Ltd) using the following PCR conditions: 94°C for 5 min, followed by 25 cycles of denaturation at 94°C for 1 min, annealing at 60°C for 1 min for AV1 gene (partial) and 52°C for 1 min for AC1 gene (partial), extension at 72°C for 1 min and a final extension at 72°C for 5 min. Similar way, movement protein (MP; BC1) of DNA B was amplified by using following conditions: initial denaturation at 94°C for 5 min, followed by 25 cycles of denaturation at 94°C for 1 min, annealing at 52°C for 1 min for BC1 gene (partial), extension at 72°C for 1 min and a final extension at 72°C for 5 min.
Detection of DNA beta satellite by universal β-satellite primers: Primer pair Beta01/Beta02 was used to assess the presence of beta-satellites in the samples. PCR was carried out in a Peltier thermal cycler L196GGD (Lark Innovative Technologies, Pvt., Ltd) using the following PCR conditions: 94°C for 5 min, followed by 25 cycles of denaturation at 94°C for 1 min, annealing at 62°C for 1 min for, extension at 72°C for 1 min and a final extension at 72°C for 5 min. The result was further analyzed by agarose gel electrophoresis.
Full-length genome amplification by RCA: The full-length genome was amplified from extracted DNA by using Illustra TempliPhiTM DNA amplification kit (GE healthcare, UK). In this reaction mixture, 0.5 μL (10-20 ng) of DNA was mixed with 5 μL of sample buffer, denatured at 95°C for 5 min and cooled to room temperature; add 5 μL of reaction buffer and 0.2 μL of enzyme mix were incubated for 18-20 h at 30°C. The reaction was finally stopped by incubating at 65°C for 10 min.
Cloning and sequencing of viral genome: Amplified products were run on 0.8% agarose gel to assess the quality of product. The concentration of DNA was checked on Nano drop (Thermo scientific, UK). Finally, 2 μg of RCA product was digested with 5U of BamHI and HindIII (Fermentas Inc.,) to obtain a fragment of approximately 2.7 kb which was cloned into the pBluescript II KS+ vector and transformed into E. coli DH5α. The clones were sequenced in both orientations using primer walking (Eurofins MWG Inc.). Further, all the positive DNA samples were also checked for the potential presence of satellite DNA-β using standard PCR conditions.
Viral genome sequence analysis: The nucleotide sequences of DNA-A (Genbank Acc.no.KF188433), DNA-B (Genbank Acc.no. KJ004521) of SLCCNV-[IN:Coi:Ash:KP1] and CroYVMB (Genbank Acc. No. KM588256) were aligned with the sequences obtained from GenBank using ClustalW program19. The complete nucleotide sequences of both the isolates were initially taken into account for similarity search by using BLASTn search program. Based on the alignment score, percentage of sequence identity and e-value, the nucleotide sequences were chosen for study.
Construction of phylogenetic tree and analysis: Multiple sequence alignments were produced using the MUSCLE algorithm available20 in MEGA v.6. Phylogenetic relationships were inferred using the neighbor joining method21 with bootstrapping22. Evolutionary distances were computed by using the P-distance5, using MEGA v.6. Bootstrap values (1000 iterations) were calculated based on the >70% majority rule and confidence limits were placed on the major nodes of the tree.
Recombination analysis in viral genomes by RDP4: Recombination analysis was done by using Recombination Detection Program Version 4 (RDP4)23. It is a window based program which detects and analyzes recombination signals in a set of aligned DNA sequences. The following three sets of DNA component-DNA-A, DNA-B and CroYVMB were taken into account. RDP4 identifies possible recombinants and also parental sequences in a number of sequences. It uses six different automated methods, namely RDP, GENECONV, MAXIMUM χ2, BOOTSCAN, CHIMERA and SISTER SCANNING. The highest acceptable probability value p = 0.05 based on the standard Bonferroni correction was used for the analysis of RDP. Bootscan analysis was carried out, with 200 replicates with a cut-off value of 95% and the g-scale value was 1 for GENECONV analysis, which is the lowest penalties, means that mismatches are prohibited.
RESULTS
Viral genome sequence and analysis: The BamHI and HindIII digested, cloned ~2.7 kb fragment genome of SLCCNV KP1 DNA-A (2739nt, KF188433) with 8 ORFs (AV1-AV3, AC1-AC5), DNA-B (2683nt, KJ004521) has 2 ORFs (BV1 and BC1). BLASTn analysis for SLCCNV DNA-A sequence (KF188433) showed 98 and 91-96% nucleotide sequence identity with DNA-A components of SLCCNV-India (AY184487) and other Cucurbitaceae-infecting SLCCNV-India isolates and SLCCNV DNA-B sequence (KJ004521) showed 93% sequence identity to DNA-B (AY184488) components of SLCCNV-India [IN: Coi: Pum]. The SLCCNV-KP1 DNA-A ORFs shared >93% sequence identity and DNA-B ORFs shared >91% sequence identity with all the other SLCCNV and ToLCNDV isolates in the database respectively (Table 1) and low percentage identity (71-79%) with ToLCNDV DNA-B was observed it shows that KP1 isolate DNA-B sequence as recombination (Table 2).
| Table 1: | Nucleotide (%) and amino acid sequence identities (%) of DNA-A sequences of SLCCNV-KP1isolate with other SLCCNV and ToLCNDV isolates |
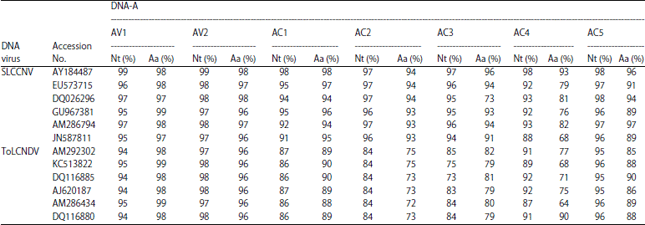 | |
| >93%-maximum sequence identity of KP1 isolate genes of DNA-A of SLCCNV with Acc.No. AY184487 | |
| Table 2: | Nucleotide (%) and amino acid sequence identities (%) of DNA-B sequences of SLCCNV-KP1 isolate with other SLCCNV and ToLCNDV isolates |
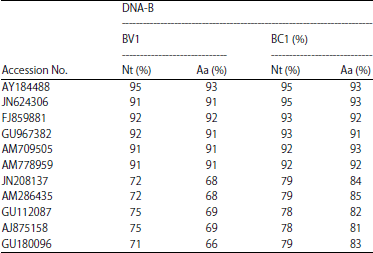 | |
| >91%-maximum sequence identity of KP1 isolate genes of DNA-B with Acc.No. AY184488 | |
| Table 3: | Nucleotide (%) and amino acid sequence identities (%) of CroYVMB-KP1isolate sequences with other CroYVMB and ToLCV isolates |
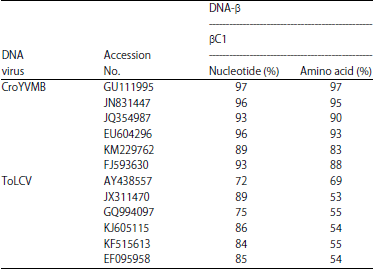 | |
| >90%-maximum sequence identity of KP1 isolate genes of CroYVMB DNA-β with Acc.No. GU111995 and >85% ToLCV DNA-β with Acc.No. JX311470 | |
Sequence comparison and analysis: The isolated DNA-β component of 1350 nt in length (KM588256) shared 88% identity with CroYVMB (Table 3). The sequence alignment of common region (CR) of DNA-A and DNA-B of SLCCNV-KP1 isolate, shows 54% identity, this low identity between the sequences was due to the difference between the TATA box and stem-loop region of the sequences. The upstream of the AC1 TATA box and CR of DNA components contained three conserved iterons (GGGGT) and identical downstream stem loop region contains the conserved TAATATTAC nonanucleotide sequence (Fig. 1). The sequence contained all the features typical of beta-satellites; a single conserved gene (known as βC1; coordinates 221-577) in the complementary-sense with the capacity to encode 118 amino acid protein. The iterons GGGGT-AT-GGGGT had been found in both the DNA-A and DNA-B of SLCCNV-KP1 (Table 4) and in beta-satellite iterons are distinct to DNA-A and DNA-B, it also possesses the nonanucleotide sequence TAATATT↓AC which is a characteristic rep-binding site, conserved with the other two DNA components (DNA-A and DNA-B) (Fig. 2).
Phylogenetic tree construction and analysis: Constructed phylogenetic tree showed, that the species were separated into the groups like SLCCNV, ToLCNDV and SLCPV (Fig. 3a-c). In accordance with the sequence analysis, the SLCCNV-KP1 was included in the SLCCNV group with 93-98% sequence identity, while ToLCNDV and SLCPV was found in distinct groups.
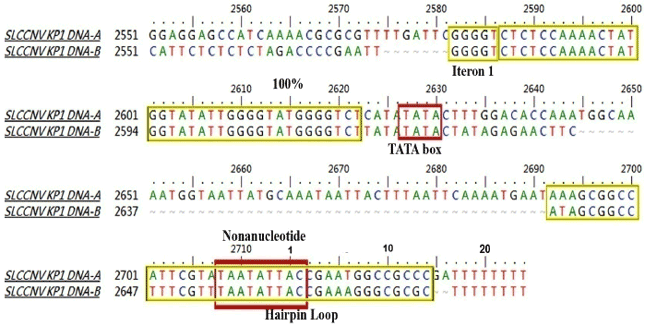 | |
| Fig. 1: | Sequence alignment of common region (CR) of DNA-A and DNA-B of SLCCNV-KP1 isolate |
| Table 4: | Selective comparison of iterons found in DNA-A and DNA-B sequences of SLCCNV-KP1 isolate with other SLCCNV and ToLCNDV isolates |
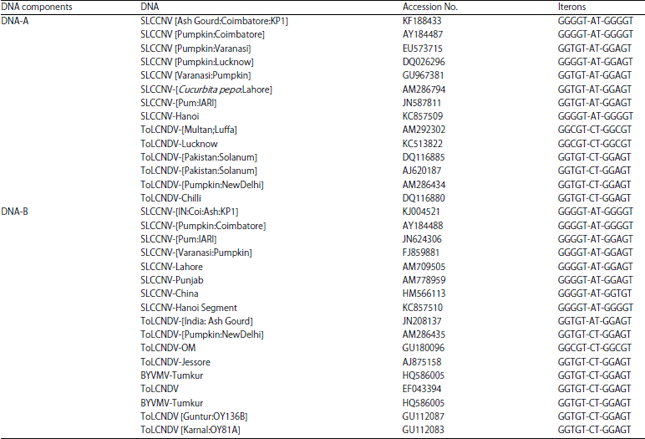 | |
| GGGGT-AT-GGGGT iterons found in DNA-A and DNA-B sequences of SLCCNV-KP1 isolate with DNA-A of SLCCNV sequence KF188433 and ToLCNDV isolates of AM292302 | |
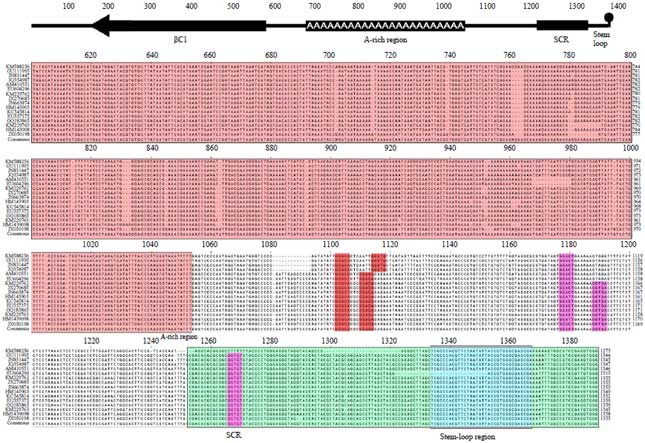 | |
| Fig. 2: | Sequence comparison and analysis of CroYVMB-KP1 isolate with other CroYVMB isolates from NCBI |
In Fig. 3a and b, the DNA-B of SLCCNV-KP1 showed 93% sequence identity with the SLCCNV [IN:Coi:Pum] and 77-79% sequence identity with ToLCNDV species. Both DNA-A and DNA-B of SLCCNV-KP1 showed identity with SLCCNV [IN:Coi:Pum] but this was not with beta satellite DNA. The beta-satellite DNA share 81% sequence identity with the CroYVMB of Bangalore, Lucknow, Pataudi, Pune and 80% with CroYVMB-Bang-Cr1. Phylogenetic analysis showed that SLCCNV-KP1 grouped more closely with SLCCNV [IN:Coi:Pum] while CroYVMB grouped more closely with CroYVMB-[IN:Bang:Croton:CR1] (100% bootstrap support) (Fig. 3c).
Recombination analysis: The occurrence of recombination events in DNA-A/DNA-B components of SLCCNV-KP1 and CroYVMB were analyzed. Among the three DNA components, DNA-A and DNA-B were responded to recombination, while DNA-B isolate KP1 did not response. The p-values of 2.4×10–13 was observed in the recombination of DNA-A at nucleotide coordinates 62-862 with ToLCNDV: DQ116880 as minor parent and SLCCNV: AY184487 as the major parent (Fig. 4). Two recombination events were found in beta-satellite component.
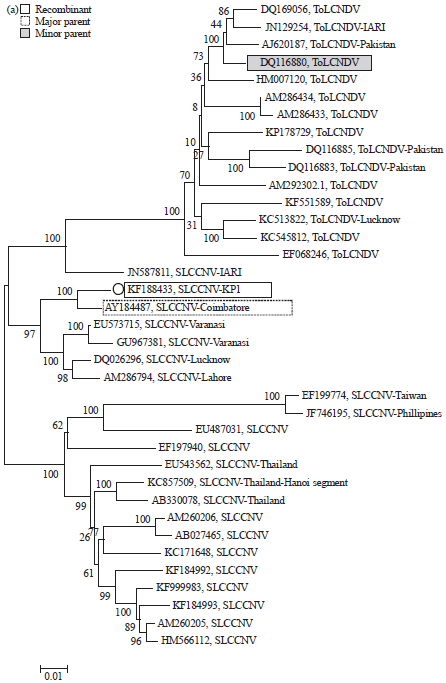 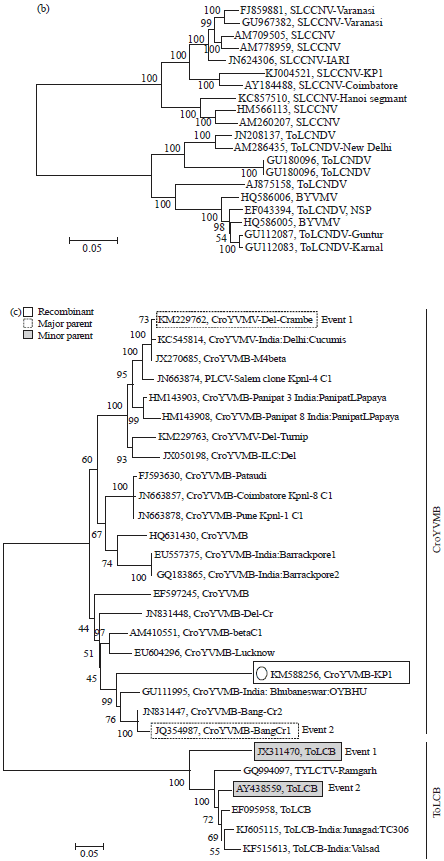 | |
| Fig. 3: | Phylogenetic trees based on neighbor joining method with the complete nucleotide sequences of (a) DNA-A of SLCCNV-KP1 isolate detected in the study compared to selected sequences SLCCNV and ToLCNDV isolates retrieved from NCBI, (b) DNA-B of SLCCNV-KP1 isolate detected in the study compared to selected sequences SLCCNV and ToLCNDV isolates retrieved from NCBI and (c) CroYVMB-KP1isolate detected in the study compared to selected sequences with other CroYVMB and ToLCV isolates retrieved from NCBI |
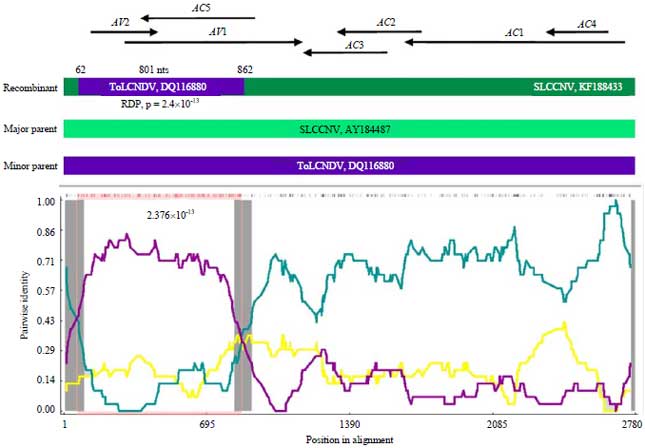 | |
| Fig. 4: | Recombination analysis between DNA-A of SLCCNV-KP1 isolate with other SLCCNV and ToLCNDV isolates |
The p-values of 2.3×10–05 was observed in the first recombination event (1241-1357) of beta-satellites while 2.6×10–05 in the second (96-146) with ToLCV as minor parent and CroYVMB as major parent (Fig. 5).
DISCUSSION
The evolution of new DNA-A species may be mainly affected by virus-host interactions i.e., due to the integration of viral DNA into the host plant nuclear genome3, DNA-A/DNA-B interactions and DNA-A/DNA-β interactions. Therefore, recombination analysis between species using RDP4 program clearly explains that DNA-A of SLCCNV-KP1 has been evolved from DNA-A of SLCCNV-[IN:Coi:Pum] as major parent and the DNA-A of ToLCNDV-[Pak:Khanewal:Chilli] as minor parent that a segment of its DNA-A has been exchanged to evolve a DNA-A of SLCCNV-KP1 species. Padidam et al.24, a new recombinant DNA-A component to evolve, a subsequent DNA-B component is required to be associated through its CR. This indicated that DNA-B of SLCCNV-KP1 is a new component and plays an important role in the evolution of new virus25. Based on the sequence and recombination analysis, separate speciation of DNA-B was understood whereas their iterons and the hairpin loop regions are found to be conserved with the DNA-A of SLCCNV-[IN:Coi:Pum]. The nick site within the origin of replication was known to be the recombination hotspot26. In addition to DNA-A and DNA-B, beta-satellites are characteristic of old world geminiviruses. In the second part of the study, a β-satellite component (~1.3 kb) was amplified by PCR using specific primers27 and the sequence (KM588256) was 88% identical to Croton yellow vein mosaic virus (CYVMV) leaf curl βC1 strain28. Interestingly, the CroYVMB was found in B. hispida it shows that Croton bonplandianum could act as reservoirs of virus populations and has donated its DNA-B to the ash gourd for severe infection of plants, which paralleled the findings, in which the co-existence of CYVMV was reported in tomato plants due to the mixed virus infections14, which could have biological and epidemiological implications under natural conditions. Based on the sequence information, the beta satellites shared 81-86% identity with Croton yellow vein mosaic beta-satellite-Bangalore; CroYVMB-[Bang-Cr1] species. CYVMV has been known to occur with yellow vein mosaic disease in a predominantly growing weed Croton bonplandianum17 since 1963.
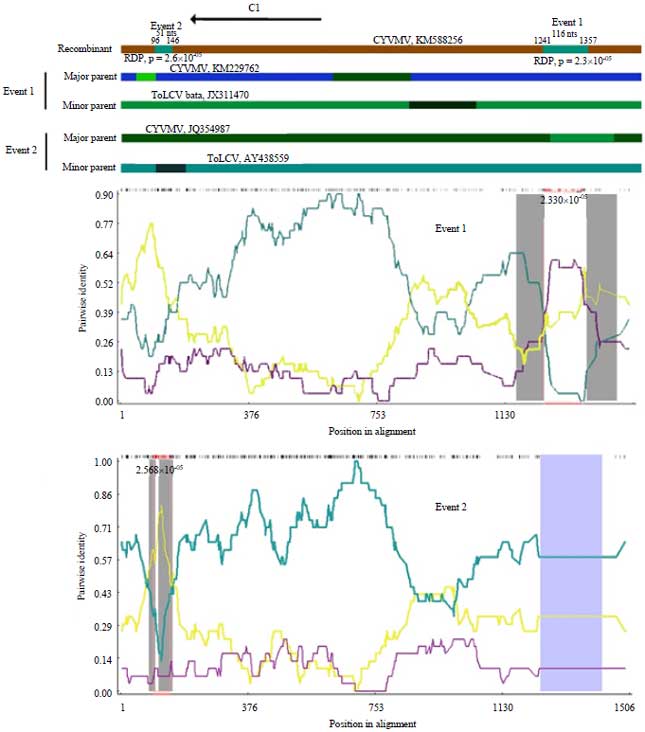 | |
| Fig. 5: | Recombination analysis between CroYVMB-KP1isolate with other CroYVMB and ToLCV |
CONCLUSION
Based on the sequence analysis, it was found that DNA-A/DNA-B of SLCCNV-KP1 isolate and CroYVMB co-exist in the infected ash gourd. The recombination analysis done by using RDP4, it is conferred that DNA-A of SLCCNV-KP1 is a potential recombinant from DNA-A of ToLCNDV and SLCCNV while DNA-B of SLCCNV-KP1 isolate has no recombination and found to be distinct, if it was recombinant the pathogenicity and new host range will be enormous mostly in economically valuable agricultural crops as very quickly.
SIGNIFICANCE STATEMENT
This study discovers the existence of SLCCNV-IN infection in B. hispida in India for the first time, it reports that the Begomovirus causing high losses in B. hispida in India is a variant of SLCCNV-IN [IN:Coi:Pum] and has been designated as Squash leaf curl China virus-India isolate KP1 based on its segment DNA-A, along with CYVMV β-satellite component ~1.3 kb (KM588256) was found to be 88% identical to CYVMV leaf curl βC1 strain. Based on the sequence analysis, it was found that DNA-A/DNA-B of SLCCNV KP1 isolate and CroYVMB co-exist in the infected ash gourd plants.
This study will help the researcher to uncover the critical areas like natural infection of two distinct Begomovirus in cognate with beta-satellite components infecting the economically important vegetable crops like ash gourd, as that many researchers were not able to explore and to understand the frequency of emerging bipartite Begomovirus strains with betasatellite genomes, indicating that the virus species had undergone recombination with diverse virulent strains as a long-term event and thus a new theory on recombination of viral genome pose a serious threat to vegetable crop cultivation.
ACKNOWLEDGMENT
The authors wish to thank University Grant Commission, New Delhi [F. No. 37-440/2009 (SR)] and DST-SERB NPDF [PDF/2016/002741] for financial assistance.
REFERENCES
- Moffat, A.S., 1999. Plant pathology: Geminiviruses emerge as serious crop threat. Science, 286: 1835-1835.
CrossRefDirect Link - Stanley, J., D.M. Bisaro, R.W. Briddon, J.K. Brown and C.M. Fauquet et al., 2005. Virus Taxonomy: Classification and Nomenclature of Viruses. In: Eighth Report of the International Committee on Taxonomy of Viruses, Fauquet, C.M., M.A. Mayo, J. Maniloff, U. Desselberger and L.A. Ball (Eds.)., Elsevier/Academic Press, London, UK., pp: 301-326.
- Bejarano, E.R., A. Khashoggi, M. Witty and C. Lichtenstein, 1996. Integration of multiple repeats of geminiviral DNA into the nuclear genome of tobacco during evolution. Proc. Nat. Acad. Sci., 93: 759-764.
CrossRefDirect Link - Varsani A., J. Navas-Castillo, E. Moriones, C. Hernandez-Zepeda and A. Idriset al., 2014. Establishment of three new genera in the family Geminiviridae: Becurtovirus, Eragrovirus and Turncurtovirus. Arch. Virol., 159: 2193-2203.
CrossRefDirect Link - Nawaz-ul-Rehman, M.S. and C.M. Fauquet, 2009. Evolution of geminiviruses and their satellites. FEBS Lett., 583: 1825-1832.
CrossRefDirect Link - Iqbal, Z., M.N. Sattar and M. Shafiq, 2016. CRISPR/Cas9: A tool to circumscribe cotton leaf curl disease. Front. Plant Sci., Vol. 7.
CrossRefDirect Link - Padidam, M., S. Sawyer and C.M. Fauquet, 1999. Possible emergence of new geminiviruses by frequent recombination. Virology, 265: 218-225.
CrossRefPubMedDirect Link - Holland, J. and E. Domingo, 1998. Origin and evolution of viruses. Virus Genes, 16: 13-21.
CrossRefDirect Link - Hughes, A.L., 1993. The evolutionary biology of viruses. Morse, S.S. (Ed.)., New York: Raven Press. Genet Epidemiol., 11: 353-353.
CrossRefDirect Link - Keese, P. and A. Gibbs, 1993. Plant viruses: Master explorers of evolutionary space. Curr. Opin. Genet. Dev., 3: 873-877.
CrossRefDirect Link - Drake, J.W., 1993. Rates of spontaneous mutation among RNA viruses. Proc. Nat. Acad. Sci., 90: 4171-4175.
CrossRefDirect Link - Medina, C.G.V., E. Teppa, V.A. Bornancini, C.R. Flores, C. Marino-Buslje and P.M.L. Lambertini, 2018. Tomato apical leaf curl virus: A novel, monopartite geminivirus detected in tomatoes in Argentina. Front. Microbiol., Vol. 8.
CrossRefDirect Link - Pramesh, D., B. Mandal, C. Phaneendra and V. Muniyappa, 2013. Host range and genetic diversity of croton yellow vein mosaic virus, a weed-infecting monopartite begomovirus causing leaf curl disease in tomato. Arch. Virol., 158: 531-542.
CrossRefDirect Link - Muniyappa, V. and G.K. Veeresh, 1984. Plant virus diseases transmitted by whiteflies in Karnataka. Proc.: Anim. Sci., 93: 397-406.
CrossRefDirect Link - Hussain, K., M. Hussain, S. Mansoor and R.W. Briddon, 2011. Complete nucleotide sequence of a begomovirus and associated betasatellite infecting croton (Croton bonplandianus) in Pakistan. Arch. Virol., 156: 1101-1105.
CrossRefDirect Link - Dellaporta, S.L., J. Wood and J.B. Hicks, 1983. A plant DNA minipreparation: Version II. Plant Mol. Biol. Rep., 1: 19-21.
CrossRefDirect Link - Larkin, M.A., G. Blackshields, N.P. Brown, R. Chenna and P.A. McGettigan et al., 2007. Clustal W and clustal X version 2.0. Bioinformatics, 23: 2947-2948.
CrossRefPubMedDirect Link - Tamura, K., G. Stecher, D. Peterson, A. Filipski and S. Kumar, 2013. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol., 30: 2725-2729.
CrossRefPubMedDirect Link - Saitou, N. and M. Nei, 1987. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol., 4: 406-425.
CrossRefPubMedDirect Link - Felsenstein, J., 1985. Phylogenies and the comparative method. Am. Natl., 125: 1-15.
CrossRefDirect Link - Martin, D.P., P. Lemey, M. Lott, V. Moulton, D. Posada and P. Lefeuvre, 2010. RDP3: A flexible and fast computer program for analyzing recombination. Bioinformatics, 26: 2462-2463.
CrossRefDirect Link - Padidam, M., R.N. Beachy and C.M. Fauquet, 1995. Tomato leaf curl geminivirus from India has a bipartite genome and coat protein is not essential for infectivity. J. General Virol., 76: 25-35.
CrossRefDirect Link - Rey, M., J. Ndunguru, L. Berrie, M. Paximadis and S. Berry et al., 2012. Diversity of dicotyledenous-infecting geminiviruses and their associated DNA molecules in Southern Africa, including the South-West Indian Ocean islands. Viruses, 4: 1753-1791.
CrossRefDirect Link - Saunders, K., I.D. Bedford, R.W. Briddon, P.G. Markham, S.M. Wong and J. Stanley, 2000. A unique virus complex causes Ageratum yellow vein disease. Proc. Natl. Acad. Sci. (USA)., 97: 6890-6895.
Direct Link - Briddon, R.W., S.E. Bull, I. Amin, A.M. Idris and S. Mansoor et al., 2003. Diversity of DNA β, a satellite molecule associated with some monopartite Begomoviruses. Virology, 312: 106-121.
CrossRefDirect Link - Riyaz, S.M., S. Deepan, M.I. Jesse, G. Dharanivasan and K. Kathiravan, 2015. New record of bipartite Squash leaf curl China virus (SLCCNV) and Croton yellow vein mosaic beta satellite associated with yellow vein disease of ash gourd in India. New Dis. Rep., Vol. 31.
CrossRefDirect Link