
Adel A. El-Morsi
Department of Botany, Faculty of Science, Mansoura University, Mansoura, Egypt
Samia A. Haroun
Department of Botany, Faculty of Science, Mansoura University, Mansoura, Egypt
Ayat M. Hassan
Department of Botany, Faculty of Science, Mansoura University, Mansoura, Egypt
Dalia G. Aseel
City of Scientific Research and Technological Applications, Department of Plant Protection and Bimolecular Diagnosis, Alexandria, Egypt
Elsayed E. Hafez
City of Scientific Research and Technological Applications, Department of Plant Protection and Bimolecular Diagnosis, Alexandria, Egypt
LiveDNA: 20.3216
International Journal of Virology
Year: 2017 | Volume: 13 | Issue: 1 | Page No.: 53-61







ABSTRACT
Background and Objective: Citrus tristeza virus (CTV) is the most destructive virus of citrus which resulted in huge economic losses to citrus crop in Egypt and all over the world. We aim in this study to determine the presence, distribution and well characterize the virus in Egypt. Materials and Methods: A number of 77 naturally infected leaves and leaf petioles of citrus plants were collected from different cultivated areas in Dakahlia governorate, Egypt. The symptoms in infected plants included tristeza or quick decline, vein clearing and leaf cupping. Moreover, stem pitting of the trunk and branches of infected trees was observed. The leaves were collected and subjected to ELISA test. Results: The results revealed that about 84% of the collected samples with symptoms were positive. The ultra thin sections of CTV-infected leaves showed the presence of viral arrays, fibrous inclusions and accumulated cytoplasmic vesicles. Also, masses of electron-dense bodies were found between the cell wall and cell membrane. Accordingly, lipid accumulations and multi vesicular bodies were abundant in the cytoplasm. More effects were observed such as nuclear membrane invaginations and chloroplast degradation. Electron microscopy examination for the purified preparation (based on PEG precipitation and differential centrifugation) revealed the presence of filamentous virus-like particles with size about 2000×11 nm. Conclusion: The virus characterization has a vital role for rapid detection of CTV and prevents prevalence to other areas. The study proved the presence of tristeza virus in Egypt and the stockholder should take steps to control its distribution all over Egypt. The virus completely destroys the cell and it is highly distributed in all visited areas. Further studies should be carried out to find suitable tools for preventing the virus hassles for citrus production.
PDF Abstract XML References Citation
Received: September 16, 2016;
Accepted: October 19, 2016;
Published: December 15, 2016
How to cite this article
Adel A. El-Morsi, Samia A. Haroun, Ayat M. Hassan, Dalia G. Aseel and Elsayed E. Hafez, 2017. Characterization of Citrus tristeza virus (CTV) Isolated from Dakahlia Governorate, Egypt. International Journal of Virology, 13: 53-61.
DOI: 10.3923/ijv.2017.53.61
URL: https://scialert.net/abstract/?doi=ijv.2017.53.61
DOI: 10.3923/ijv.2017.53.61
URL: https://scialert.net/abstract/?doi=ijv.2017.53.61
INTRODUCTION
Citrus is one of the most valuable crops in Egypt. It covers an area of 166.207 ha (70.8% exist in the Nile Delta, whereas 29.2% are located in the new reclaimed desert areas) with a total production of 4.452.200 t and export about 1081.900 t1.
Egypt is likely to retain its position as the world’s 6th largest orange producer and one of the top orange exporters in the world. In 2015, total area planted with oranges was 133.236 ha producing about 2.750.000 t and export 1.300.000 t with export value exceed 195.000 $ annually2.
Citrus tristeza virus (CTV) a member of the genus Closterovius of the Closteroviridae3 is the most destructive virus in citrus growing regions and cause huge economic losses to the citrus industry4,5 which is responsible for destroying of about 80-100 million trees grafted on sour orange worldwide4,6,7. At the recent times, more than 400 million citrus trees are at risk to CTV decline globally5,8.
The virus is phloem limited and the particles are flexuous, thread like filamentous, about 2000×11 nm in size9. It has the largest known (19.3 kb) positive-sense single stranded RNA genome among the plant viruses and contain 12 open reading frames that encode at least 19 proteins3,10, 11. The virus is semi-persistently transmitted4,12 by various aphid species with Toxoptera citricida being the most efficient vector13, which is not present in the Mediterranean basin.
The use of enzyme linked inmunosorbent assay (ELISA) for CTV provide rapid and easy detection, where it allowed the processing of large numbers of samples in a short period of time in contrast to biological indexing which is expensive and requiring several months for symptoms to develop14-16.
The CTV particles are difficult to purify and concentrate17 because of the limitation to phloem tissues18 and tendency of these particles to aggregate during the precipitation steps19. Several trials were reported for CTV purification, including the procedures depend on polyethylene glycol precipitation followed by differential centrifugation17, density gradient centrifugation in cesium chloride (CsCl) after fixation with formaldehyde20, Cs2SO4 density gradient centrifugation21, a sucrose density gradient22 or short Cs2SO4-sucrose cushion step gradient19.
The CTV produces inclusion bodies in large aggregates that were found in phloem and associated cells23. There are three types of CTV inclusion bodies: Viral arrays, fibrous inclusions and accumulated cytoplasmic vesicles were observed in infected phloem cells. The cytopathology in CTV-infected plant was also observed, including chloroplast degradation, mitochondria vesiculation and nuclear membrane invagination24. The present investigation was conducted to detect tristeza virus, the most destructive virus infecting citrus in Dakahlia governorate for the 1st time, following by identification of it as a start point for designing high sensitive and specific diagnostic tool.
MATERIALS AND METHODS
Samples collection: A number of 77 samples (leaves and leaf petioles) of sweet orange (Citrus sinensis) plants with characteristic symptoms of CTV were collected from the different fields in Dakahlia governorate (Green area) during January and April, 2013-2015 (Fig. 1). These samples were then transferred directly to the laboratory for testing by ELISA.
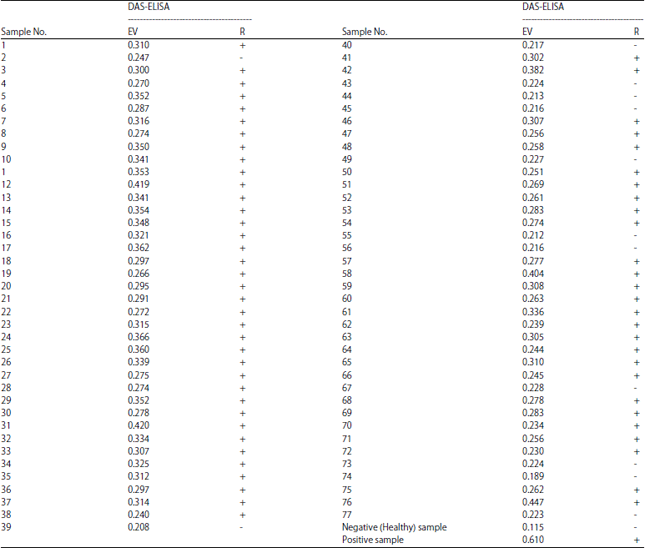
ELISA test: The DAS-ELISA was used for virus detection as described by Clark and Adams25 and Ananthakrishnan et al.26 using the DSMZ kit (DSMZ Inc., Germany). The 96-well plate was coated with coating buffer (pH 9.6) containing IgG. The coated plate was incubated at 37°C for 2 h and washed 3 times with phosphate buffer saline tween (pH 7.4). Leaves and leaf petioles (0.1 g) was ground in 1 mL extraction buffer (pH 7.4) and centrifuged at 3000 rpm for 10 min. The supernatant was collected as the antigen. The antigen was added to the wells and incubated overnight at 4°C. The plate was washed 3 times again, alkaline phosphatase-conjugated Ig G-AP was added and incubated for 2 h 37°C. After washing, the substrate buffer containing 0.1% p-Nitrophenyl phosphate was added to the wells and incubated at room temperature for 30-60 min. The absorbance values were measured at wavelength 405 nm by the BioTek ELISA reader (EL x800, USA). Samples were considered positive if the OD405 values were equal to or more than 2 folds of the healthy control27.
Cytopathological effect: Ultra thin sections of citrus leaves and leaf petioles showing virus-like symptoms were prepared according to Zhou et al.24 and Allam et al.28 Small pieces were fixed in cold 3% glutraldehyde in phosphate buffer, pH 7.2 for 3 h. Then these specimens were post fixed in 1% OsO4 for 1.5 h. After dehydration, they were embedded in spur medium. Thin sections were stained with Uranylacetateand Lead citrate. The ultra thin sections were examined with transmission electron microscope (JEOLJEM-2100, Japan) in the Electron Microscope Unit, Faculty of Agriculture, Mansoura University, Mansoura, Egypt.
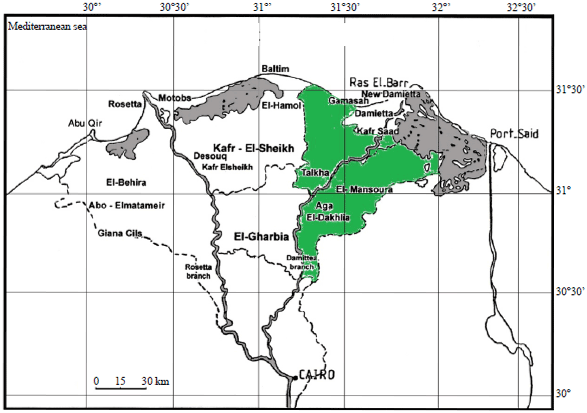 | |
| Fig. 1: | Sketch map showing the study area where the samples were collected |
Virus purification: The CTV was purified according to the method described by Bar-Joseph et al.17 with some modification. Virus-infected leaves and leaf petioles were ground in liquid nitrogen then the powder was extracted in 0.05 M tris [tris (hydroxymethyl) amino methane]-HCl buffer, pH 7.4 (1 g/2.5 mL). After filtration through cheese cloth and centrifugation for 10 min at 4000 rpm, 4 g of polyethylene glycol (PEG 6000) and 4 mL of NaCl (20%) were added to each 100 mL of supernatant followed by slowly stirring overnight at 4°C. The pellet was collected by centrifugation at 10000 rpm for 15 min and resuspended in 0.04 M sodium phosphate buffer, pH 8.2 (25 mL/100 mL original extract). After 10 min centrifugation at 6500 rpm, the supernatant contain virus particles was further purified by centrifugation at 16000 rpm for 8 h. All centrifugation steps were conducted under cooling conditions at 4°C using ultracentrifuge D-37520 Osterode (3-18 K, Germany). The virus pellet was resuspended in a volume of 1 mL 0.04 M sodium phosphate buffer (pH 8.2).
Negative staining: To study the morphological characteristics of purified virus, a drop of purified preparation was placed for 3 min on a copper carbon coated grid (400 mesh) which previously was washed with 0.01 M potassium phosphate buffer. The grid was then washed 3 times with d.H2O and dried with Whitman tissue paper followed by staining with 2% aqueous uranylacetate (w/v) as reported by Morales et al.29. The grid was examined with transmission electron microscope (JEOLJEM-2100, Japan) in the Electron Microscope Unit, Faculty of Agriculture, Mansoura University, Mansoura, Egypt.
RESULTS
Symptoms and ELISA detection for CTV: Figure 2 shows the CTV-like symptoms on sweet orange (Citrus sinensis) plantsin cultivated farms under natural conditions in Dakahlia governorate. These symptoms consisted of yellowing, yellow dots, vein clearing, leaf cupping and stem pitting as a diagnostic symptom for citrus tristeza. All samples were tested for the presence of CTV-infection using DAS-ELISA. Table 1 showed that, out of 77 citrus tested samples, 65 samples gave positive ELISA values ranged from 0.230-0.447, while 12 samples showed negative ELISA value ranged from 0.189-0.228. Thus, the infection percent in Dakahlia governorate was calculated as 84%. Vein clearing and leaf cupping represents the characteristics symptoms of CTV in positive infection judged by DAS-ELISA are showed in Fig. 3.
 | |
| Fig. 2(a-b): | Overview of sweet orange (Citrus sinensis) plants showing virus-like symptoms under open field conditions in Dakahlia governorate, (a) Yellowing and tristeza or quick decline and (b) Stem pitting, a diagnostic symptom |
| Table 1: | DAS-ELISA detection of CTV in Citrus sinensis exhibited virus-like symptoms and collected from different locations at Dakahlia governorate during January and April (2013-2015) |
 | |
 | |
| Fig. 3(a-b): | Characteristic symptoms of CTV on Citrus sinensis samples showed positive infection as judged by DAS-ELISA, (a) Vein clearing with yellow dots and (b) Leaf cupping |
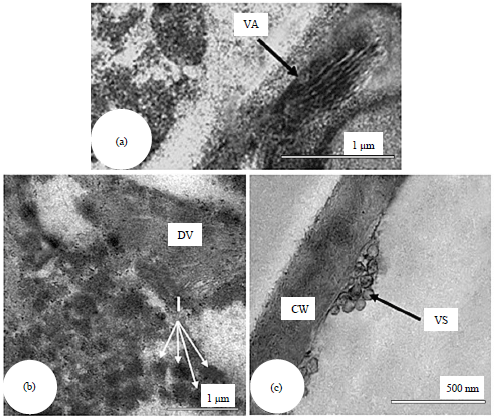 | |
| Fig. 4(a-c): | Electron micrographs of ultrathin sections prepared from leaves of sweet orange (Citrus sinensis) plants infected with CTV showing, (a) Viral arrays (VA), (b) Fibrous inclusions (I) and (c) Accumulated cytoplasmic vesicles (VS), CW: Cell wall and DV: Disorganized virus particles |
Cytopathological effect
CTV-induced inclusions: Data presented in Fig. 4 show the inducing of the characteristic inclusions of CTV that appeared as viral arrays which composed of virions aligned parallel to one another and commonly seem to be spindle shaped, fibrous inclusions that formed by aggregates of fibrous material and finally accumulated vesicles were appeared in the cytoplasm of infected cells resembling Beet Yellow Virus (BYV)-like vesicles.
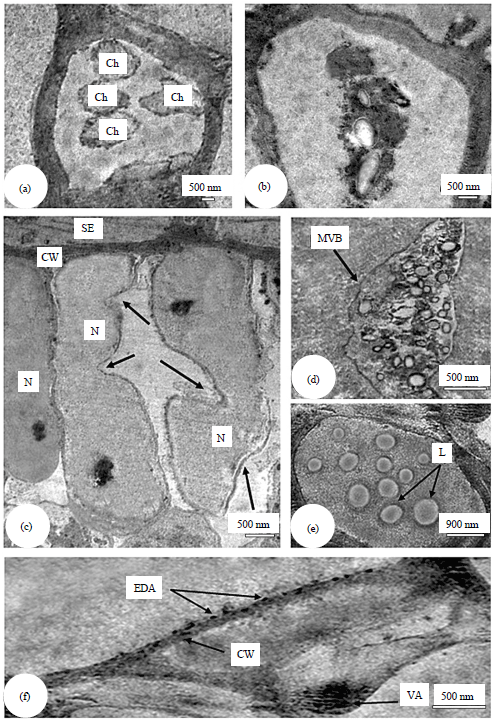 | |
| Fig. 5(a-f): | Transmission electron micrographs of ultrathin sections of sweet orange (Citrus sinensis) leaves infected with CTV showing, (a) Aggregated degrading chloroplasts (Ch), (b) Cell disorganization, (c) Nucleus (N) with multiple invaginations (arrows), (d) Multi vesicular bodies (MVB), (e) Lipid accumulations (L) and (f) Electron dense accumulations (EDA) between the cell membrane and cell wall (CW) |
Cytopathology: Transmission electron micrographs of ultrathin sections of infected citrus leaves and leaf petioles showed aggregation of chloroplasts with abnormal shaped, numerous invaginations of the nuclear membrane, small number of electron dense accumulation was usually present between cell wall and cell membrane, large number of multi vesicular bodies and lipid accumulations were common (Fig. 5).
Purification and morphology of CTV particles: The use of PEG as described by Bar-Joseph et al.17 was successfully used for purification of CTV from virus infected tissues. Where, purified virus preparations had a typical UV absorption spectrum of anucleoprotein with a 260/280 ratio of 1.293 (Fig. 6). Also, electron microscopy of this preparation revealed the presence of flexuous filamentous like particles with dimensions of 2000×11 nm (Fig. 7).
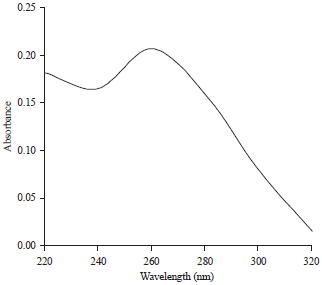 | |
| Fig. 6: | Ultraviolet absorbance profile scans of purified CTV by PEG and differential centrifugation procedures, A260: 0.207 and A280: 0.160 |
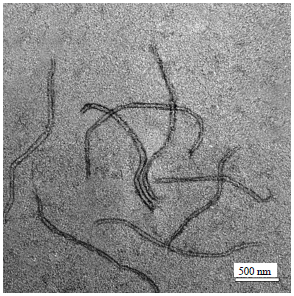 | |
| Fig. 7: | Electron micrograph of purified CTV particles that were negatively stained with 2% uranyl acetate, filamentous like particles were occurred with 2000 nm in length and 11 nm in width |
Yield of purified CTV was estimated to be 123 μg/100 g fresh citrus leaves and leaf petioles (Extension coefficient = 1.2 at 260 nm).
DISCUSSION
Citrus tristeza virus (CTV) is the most economically damaging virus of citrus worldwide5 and represent a major risk to the Egyptian citrus industries30,31. In the present study, the characteristic symptoms of CTV which recorded from naturally infected citrus plants were quick decline, vein clearing, leaf cupping and stem pitting. These observations are similar to those reported by Bar-Joseph et al.9, Moreno et al.4 and Moreno and Garnsey5.
Enzyme linked immunosorbent assay (ELISA) is generally the most accepted and widespread method for the detecting of a large number of plant viruses25,32. These procedures can provide sensitive and rapid detection using crude extracts from citrus for early identification and removing infected trees before natural spread occurs33. The leaves of collected samples were used to determine the presence of CTV infection by DAS-ELISA because it was reported that the midribs and petioles were the best tissues for detection of CTV by ELISA34. The results revealed a high CTV incidence (84%) in Dakahlia governorate where the trees showed sever CTV symptoms, which can be worrying in this regard.
Plant viral inclusions have shown in many hosts to be objective intracellular evidence of viral infection furthermore their role in viral characterization. Transmission electron microscopy of ultrathin sections of sweet orange plants revealed the presence of viral arrays which are similar to the banded inclusions previously observed in infected cells with BYV35, where both of them were composed of stacks of virions. Also, fibrous inclusions were recorded which were formed by masses of fibrous materials as previously reported for BYV24,35,36. Lesemann37 reported that, the vesiculated inclusions accumulated in infected cells are feature shared by all Closteroviridae members. These accumulations have been observed in CTV-infected cells as described by Kitajima and Costa38 and Bar-Joseph et al.39.
In addition, the cytopathology that was observed in CTV infected sweet orange plants, such as lipid accumulations, nuclear membrane invagination and electron dense accumulation along the cell wall was characteristics to those appeared in infected cells by viruses in family Closteroviridae39-42. Ultra structural data showed the occurrence of CTV in infected cells as disorganized viral particles, this observation mirrored that of Zhou et al.24.
There are several difficulties in purification and concentration of CTV particles. The relatively low concentration of virus particles, their tendency to aggregate and fragment and their restriction to phloem cells requires large quantities of tissues rich in phloem22. A successful procedures described by Bar-Joseph et al.17 was approached with some modification depending on gentle grinding, PEG precipitation, minimum of high speed centrifugation and prolonged stirring for suspension partly overcomes these problems as well as enable us to obtain even higher yields of CTV. We considered that, a successful CTV purification schedule could be derived from firstly use of virus source tissue that contained appreciable concentrations of CTV, secondly the effective extraction procedure through powderizing the infected tissue with liquid nitrogen and gentle grinding. Finally, the elimination of additional cycles of differential centrifugation.
The use of electron microscope to investigate the presence of virus particles of CTV in purified virus preparations showed that, filamentous viral particles with diameter 2000×11 nm were found. These results agree with viral dimension which ranged from 1900-2000 nm that found by Bar-Joseph et al.17, Tsuchizaki et al.22, Kitajima et al.43,44 and Price45. The sensitivity of filamentous particles to fragmentation has been observed with other long filamentous viruses46-48. In conclusion, CTV represents a tremendous threat to the citrus industryin Egypt, so it must be overcome by early detection and rapid characterization of causative agent.
CONCLUSION
It can conclude that tristeza virus is presented with high percentage in all visited area (84%). This observation revealed that, the virus is so active and all the conditions are suitable for his propagation and distribution. Further studies are needed to control and prevent the viral transmission to other govern orates in Egypt.
ACKNOWLEDGMENT
We do acknowledgment all the workers in the plant pathology lab and all the farmers for their help and support during this study.
REFERENCES
- Moreno, P., S. Ambros, M.R. Albiach-Marti, J. Guerri and L. Pena, 2008. Citrus tristeza virus: A pathogen that changed the course of the citrus industry. Mol. Plant Pathol., 9: 251-268.
CrossRefDirect Link - Ruiz-Ruiz, S., P. Moreno, J. Guerri and S. Ambros, 2007. A real-time RT-PCR assay for detection and absolute quantitation of Citrus tristeza virus in different plant tissues. J. Virol. Methods, 145: 96-105.
CrossRefDirect Link - Atta, S., C.Y. Zhou, Z.H.O.U. Yan, M.J. Cao and X.F. Wang, 2012. Distribution and research advances of Citrus tristeza virus. J. Integrat. Agric., 11: 346-358.
CrossRefDirect Link - Baruah, B.R., A. Kashyap and P.D. Nath, 2015. Citrus tristeza virus a worldwide threat to citriculture: Advances made in its research and future thrust. Res. J. Agric. For. Sci., 3: 7-14.
Direct Link - Bar-Joseph, M., R. Marcus and R.F. Lee, 1989. The continuous challenge of citrus tristeza virus control. Ann. Rev. Phytopathol., 27: 291-316.
CrossRefDirect Link - Karasev, A.V., V.P. Boyko, S. Gowda, O.V. Nikolaeva and M.E. Hilf et al., 1995. Complete sequence of the Citrus tristeza virus RNA genome. Virology, 208: 511-520.
CrossRefDirect Link - Karasev, A.V., 2000. Genetic diversity and evolution of closteroviruses. Ann. Rev. Phytopathol., 38: 293-324.
CrossRefDirect Link - Garnsey S.M., D. Gonsalves and D.E. Purcifull, 1977. Mechanical transmission of citrus tristeza virus. Phytopathology, 67: 965-968.
CrossRefDirect Link - Rocha-Pena, M.A., R.F. Lee, R. Lastra, C.L. Niblett, F.M. Ochoa-Corona, S.M. Garnsey and R.K. Yokomi, 1995. Citrus tristeza virus and its aphid vector Toxoptera citmicida: Threats to citrus production in the Caribbean and central and North America. Plant Dis., 79: 437-445.
CrossRefDirect Link - Rocha-Pena, M. A. and R.F. Lee, 1991. Serological techniques for detection of citrus tristeza virus. J. Virol. Methods, 34: 311-331.
CrossRefPubMedDirect Link - Bar-Joseph, M., G. Loebenstein and J. Cohen, 1970. Partial purification of viruslike particles associated with the citrus tristeza disease. Phytopathology, 60: 75-78.
Direct Link - Bar-Joseph, M., D.J. Gumpf, J.A. Dodds, A. Rosner and I. Ginzberg, 1985. A simple purification method for citrus tristeza virus and estimation of its genome size. Phytopathology, 75: 195-198.
Direct Link - Bar-Joseph, M., G. Loebenstein and J. Cohen, 1972. Further purification and characterization of threadlike particles associated with the citrus tristeza disease. Virology, 50: 821-828.
CrossRefDirect Link - Gonsalves, D., D.E. Purcifull and S.M. Garnsey, 1978. Purification and serology of citrus tristeza virus. Phytopathology, 68: 553-559.
CrossRefDirect Link - Tsuchizaki, T.A., A. Sasaki and Y. Saito, 1978. Purification of Citrus tristeza virus from diseased citrus fruits and the detection of the virus in citrus tissues by fluorescent antibody techniques. Phytopathology, 68: 139-142.
CrossRefDirect Link - Brlansky, R.H. and R.F. Lee, 1990. Numbers of inclusion bodies produced by mild and severe strains of citrus tristeza virus in seven citrus hosts. Plant Dis., 74: 297-299.
Direct Link - Zhou, C.L.E., E.D. Ammar, H. Sheta, S. Kelley, M. Polek and D.E. Ullman, 2002. Citrus tristeza virus ultrastructure and associated cytopathology in Citrus sinensis and Citrus aurantifolia. Can. J. Bot., 80: 512-525.
CrossRefDirect Link - Clark, M.F. and A.N. Adams, 1977. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol., 34: 475-483.
CrossRefPubMedDirect Link - Ananthakrishnan, G., T. Venkataprasanna, A. Roy and R.H. Brlansky, 2010. Characterization of the mixture of genotypes of a Citrus tristeza virus isolate by reverse transcription-quantitative real-time PCR. J. Virol. Methods, 164: 75-82.
CrossRefDirect Link - Morales, F.J., A.L. Niessen, M. Castano and L. Calvert, 1990. Detection of a strain of soybean mosaic virus affecting tropical forage species of Centrosema. Plant Dis., 74: 648-651.
Direct Link - Amin, H.A., F. Fonseca, C. Santos and G. Nolasco, 2006. Typing of Egyptian Citrus Tristeza Virus (CTV) isolates based on the capsid protein gene. Phytopathol. Mediterranea, 45: 10-14.
Direct Link - Fahmy, H., E.S. Salama and A.M. D'Onghia, 2009. State of the art of Citrus tristeza virus (CTV) in Egypt. Options Mediterraneennes, 65: 111-113.
Direct Link - Salomone, A., M. Mongelli, P. Roggero and D. Boscia, 2004. Reliability of detection of Citrus tristeza virus by an immunochromatographic lateral flow assay in comparison with ELISA. J. Plant Pathol., 86: 43-48.
Direct Link - Bar-Joseph, M., S.M. Garnsey, D. Gonsalves, M. Moscovitz, D.E. Purcifull, M.F. Clark and G. Loebenstein, 1979. The use of enzyme-linked immunosorbent assay for detection of citrus tristeza virus. Phytopathology, 69: 190-194.
Direct Link - Anfoka, G., M.K. Abhary, I. Fattash and M.K. Nakhla, 2005. Occurrence and distribution of Citrus Tristeza Virus (CTV) in the Jordan valley. Phytopathol. Mediterranea, 44: 17-23.
Direct Link - Gowda, S., T. Satyanarayana, C.L. Davis, J. Navas-Castillo and M.R. Albiach-Marti et al., 2000. The p20 gene product of Citrus tristeza virus accumulates in the amorphous inclusion bodies. Virology, 274: 246-254.
CrossRefDirect Link - Bar-Joseph, M., G. Loebenstein and J. Cohen, 1976. Comparison of Particle Characteristics and Cytopathology of CTV with other Morphologically Similar Viruses. In: Proceedings of the 7th Conference of the International Organization of Citrus Virologists (IOCV), Calavan, E.C. (Ed.). University of California Press, Riverside, California, pp: 39-46.
- Pinto, R.L., L.L. Hoefert and G.L. Fail, 1988. Plasmalemma deposits in tissues infected with lettuce infectious yellows virus. J. Ultrastruct. Mol. Struct. Res., 100: 245-254.
CrossRefDirect Link - Faoro, F., R. Tornaghi, S. Cinquanta and G. Belli, 1992. Cytopathology of grapevine leafroll associated virus III (GLRaV-III). Rivista Patol. Vegetale, 2: 67-83.
Direct Link - Faoro, F. and R. Carzaniga, 1995. Cytochemistry and immunocytochemistry of the inclusion bodies induced by grapevine leafrollassociated closteroviruses GLRaV-1 and GLRaV-3. Rivista Patologia Vegetale, 5: 85-94.
Direct Link - Kitajima, E.W., D.M. Silva, A.R. Oliveira, G.W. Muller and A.S. Costa, 1964. Thread-like particles associated with tristeza disease of citrus. Nature, 201: 1011-1012.
CrossRefDirect Link - Price, W.C., 1966. Flexuous rods in phloem cells of lime plants infected with citrus tristeza virus. Virology, 29: 285-294.
CrossRefDirect Link - Bar-Joseph, M. and R. Hull, 1974. Purification and partial characterization of sugar beet yellows virus. Virology, 62: 552-562.
CrossRefDirect Link - Smookler, M. and G. Loebenstein, 1974. Carnation yellow fleck virus. Phytopathology, 64: 979-984.
CrossRefDirect Link