
A.A.A. Sallam
Department of Agriculture Botany, Faculty of Agriculture, Suez Canal University, Ismailia, Egypt
Azza G. Farag
Department of Virus and Phytoplasma Research, Plant Pathology Research Institute, Agriculture Research Center, Giza, Egypt
Department of Biotechnology, Faculty of Science, Taif University, Kingdom of Saudi Arabia
Essam K. Elbeshehy
Department of Agriculture Botany, Faculty of Agriculture, Suez Canal University, Ismailia, Egypt
Department of Biology, Faculty of Science, Jeddah University, Kingdom of Saudi Arabia
Ahmed K. El Attar
Department of Virus and Phytoplasma Research, Plant Pathology Research Institute, Agriculture Research Center, Giza, Egypt
Sherif A. Sabik
Department of Virus and Phytoplasma Research, Plant Pathology Research Institute, Agriculture Research Center, Giza, Egypt
International Journal of Virology
Year: 2015 | Volume: 11 | Issue: 2 | Page No.: 77-86







ABSTRACT
Four different patterns of chlorotic blotches, mosaic, yellowish mottling and deformation were observed and collected in four fig-growing areas of North Egypt from cv. Sultany farms. Fig. mosaic virus like symptoms were successful purified through transmission an eriophyid mite, Aceria ficus Cotte to healthy potted rooted cuttings grown under glasshouse conditions. Young leaves appearing typical symptoms caused by FMV. Electron microscope observations of thin-sectioned tissues from symptomatic leaves showed Double Membrane Bodies (DMBs) characteristic for FMV particles compared with symptomless leaves. Viral particles belong to family of Bunyaviridae. The RT-PCR performed on total RNA extracted from leaf tissue infected with the studied virus confirmed the specificity of the primers of RNA-dependent RNA polymerase (RdRp) gene for RNA-dependent RNA polymerase. The size of the PCR amplified fragment was 302 bp. The sequence and phylogenetic analysis indicated that the Egyptian FMV isolate was closely related to other FMV isolates, especially the Italian and Arkansas ones with 80%similarity. The genetic relationship between the RdRp gene for the Egyptian isolate of FMV with other FMV sequences on the Genbank was studied.
PDF Abstract XML References Citation
Received: May 22, 2015;
Accepted: July 11, 2015;
Published: August 04, 2015
How to cite this article
A.A.A. Sallam, Azza G. Farag, Essam K. Elbeshehy, Ahmed K. El Attar and Sherif A. Sabik, 2015. Partial Nucleotide Sequence of the Family Bunyaviridae Associated with a Mosaic-Diseased Fig. in North Egypt. International Journal of Virology, 11: 77-86.
DOI: 10.3923/ijv.2015.77.86
URL: https://scialert.net/abstract/?doi=ijv.2015.77.86
DOI: 10.3923/ijv.2015.77.86
URL: https://scialert.net/abstract/?doi=ijv.2015.77.86
INTRODUCTION
Common fig, Ficus carica L. is one of the oldest cultivated Mediterranean fruit (Zohary and Hopf, 2000; Hafez et al., 2011). It is a member of family Moraceae (Watson and Dallwitz, 2004; Alhudaib, 2012). Fig. fruit is well known for its nutritive value and is consumed fresh or dry worldwide. Fig. fruits are also recognized for their mild laxative activity (Baraket et al., 2009) . In the last few years, poor growth of fig trees, scant yield and low quality of fruits have been a common complain of growers in different regions of southern Egypt. Fig. Mosaic Virus (FMV), which was described recently, is the only characterized causal agent of Fig. Mosaic Disease (FMD). Fig. mosaic has a wide distribution and has been reported in United States, Turkey, England, Algeria, Tunisia, Syria, Spain, Italy, Jordan, New Zealand, Puerto Rico, Greece, Israel, China and Australia (Blodgett and Gomec, 1967). Further, it seems likely that fig mosaic is present in all countries, where figs are grown. Condit and Horne (1933) made the first critical study of this disease in 1933 and was found to be transmitted by grafting and in nature by the eriophyid mite Aceria ficus (Flock and Wallace, 1955). The virus is transmitted by vegetative propagation of infected cuttings from diseased trees and by budding or grafting of infected propagate buds or scions to healthy stocks. Earlier observations of thin-sectioned leaf tissues of the Egyptian fig accession, had shown the presence in parenchyma cells of the so-called Double-Membrane Bodies (DMBs) consistently associated with FMD (Appiano et al., 1995). Ultra structural studies have shown FM-affected plants to have a characteristic cytopathology. The cells of the symptomatic plants contain quasi-spherical, Double-Membrane-Bound particles (DMPs) of 100-200 nm in diameter present in the cytoplasm (Ahn et al., 1996). Such bodies were originally observed by Bradfute et al. (1970) in figs and other plants affected by mite transmitted diseases and later described in detail (Martelli and Russo, 1984; Martelli, 1991; Martelli et al., 1993). The consistent association of DMBs with mosaic symptoms and the results of molecular investigations strongly indicated that DMBs are particles of FMV (Elbeaino et al., 2006, 2007, 2011; Elbeshehy and Elbeaino, 2011). Fig. Mosaic Virus (FMV) a member of the newly described genus Emaravirus, has four negative-sense genomics RNAs ranging in size from 7.09-1.47 kb (Walia et al., 2010). The FMV genomic RNAs are not capped or polyadenylated and all four FMV genomic RNAs contain complementary sequences (18-20 nt, depending on the RNA segment) at their 5 and 3 termini. These likely base pair to form a panhandle-like structure which is one of the hallmarks of negative-sense multipartite ssRNA viruses, including tenuiviruses and viruses in the families Bunyaviridae and Arenaviridae (Falk and Tsai, 1998; Walter and Barr, 2011). Each FMV genome segment codes a single Open Reading Frame (ORF) in the virus complementary polarity RNA (vcRNA) (Walia and Falk, 2012; Elair et al., 2014). These ORFs have been annotated as follows. The FMV vcRNA 1 (7093 nt) codes for the 264 kDa RNA dependent RNA polymerase protein (RdRp). The FMV RdRp has 47% amino acid identity with the Emaravirus RdRp and 13% identity with the RdRp coded by some members of Bunyaviridae (Elbeaino et al., 2009b). This investigation was carried out to isolate and identified the Fig. mosaic virus affecting fig trees on the bases of symptomatology, mite transmission, electron microscopy and RT-PCR. The genetic relationship between the RdRp genes for RNA-dependent RNA polymerase for the Egyptian isolate of FMV with other FMV sequences on the GenBank was studied.
MATERIAL AND METHODS
Field survey and plant material: Four types of different symptoms were collected from naturally infected fig trees of cv. Sultany in the four fig-growing province of the northern region of Egypt-Ismailia governorate including College of Agriculture farm-Suez Canal University; Al-Qantarah al-Gharbiyah farm, Al-Tall al-Kabir farm and Abu Sultan farm. Sampling was conducted during May, 2013 for both symptomless and MD-symptomatic trees. Symptoms include, chlorotic blotches, mosaic, yellowish mottling and deformation are recalling virus-like diseases (Fig. 1). Symptomatic leaves gave positive reaction with specific primers for FMV were separated to be used for FMV inoculation by Aceria ficus. The causal virus was transmitted to healthy potted rooted cutting cv. Sultany from MD-symptomatic trees under glass house conditions at 22°C. Mite inoculated plants were observed for symptom expression after two months from inoculation.
Tissue processing and photomicrography: For thin sectioning, tissue pieces from veins and mesophyll tissues of the discolored areas of young leaves were processed according to standard procedures (Martelli and Russo, 1984), i.e., fixation in 4% glutaraldehyde in 0.05 M phosphate buffer for 2 h, post-fixation in 1% osmium tetroxide for 2 h, staining overnight in 2% aqueous uranyl acetate, dehydration in ethanol and embedding in Spurr’s medium.
 | |
| Fig. 1(a-d): | Symptoms of fig-mosaic diseased plants observed in inspected fig orchards and showing a wide range of foliar discoloration and malformation symptoms, (a) College of agriculture farm-SCU, (b) Al-Qantarah al-Gharbiyah farm, (c) Al-Tall al-Kabir farm and (d) Abu Sultan farm |
Thin sections were stained with lead citrate and viewed with a JOEL-JEA100 CX electron microscopy Unit (Cairo University Research Park). Negative controls consisted of leaf tissues from a PCR-negative fig seedling.
Detection of FMV by reverse transcription-polymerase chain reaction: One hundred milligram of fig leaves tissues were ground in liquid nitrogen using a pestle and mortar and transferred to a microfuge tube. Total RNA was extracted using gene jet ™ RNA purification kit (fermentas, USA) following the manufacturer’s instructions. The detection of FMV in RT-PCR was conducted using one set of specific primers, whose nucleotides sequences and use-conditions were previously described (Elbeaino et al., 2009a, 2011; Gattoni et al., 2009; Elbeshehy and Elbeaino, 2011). The one step RT-PCR was performed using Verso TM one step RT-PCR kit (Thermo scientific, ABgene, USA) in a total volume of 25 μL containing 12.5 μL of one step PCR master mix (2x), 3 μL RNA (4 ng μL–1) extract, 1.25 μL RT-Enhancer, 3 μL of each primer (10 μM) and 0.5 μL Verso enzyme mix, made up to the final volume with 4.75 μL of nuclease-free water. The specific primer pair used for the detection of the RdRp gene for RNA-dependent RNA polymerase of FMV was FMV-F: CGGTAGCAAATGGAATGAAA and FMV- R : AACACTGTTTTTGCGATTGG. This primer pair was designed to amplify a portion of the RdRp gene (302 bp).
PCR conditions: The PCR consisted of reverse transcription at 50°C for 15 min, an initial denaturation step at 95°C for 2 min, followed by 35 cycles of 94°C for 30 sec, 56°C for 1 min, 72°C for 30 sec and a final extension at 72°C for 10 min.
Analysis of amplified products: The RT-PCR products were stained with gel star (Lonza, USA) and analyzed on 1% agarose gels in 0.5X TBE buffer then visualized by UV illumination using Gel Documentation System (Gel Doc 2000, Bio-Rad, USA). Fragments were sized using a 100 bp marker.
Gel extraction and DNA sequencing: The fragment of the RT-PCR product was extracted from the agarose gel using QIAquick Gel Extraction Kit from QIAGEN according to the manufacture’s instructions. The fig mosaic virus RdRp gene for RNA-dependent RNA polymerase of FMV (302 bp) was sequenced in one direction using the FMV-r specific primer. The nucleotide sequence was analyzed using DNAMAN Sequence Analysis Software (Lynnon BioSoft. Quebec, Canada) and compared with those of fig mosaic virus homology sequences available in GenBank.
RESULTS AND DISCUSSION
Field survey and observed symptoms: Four different patterns of chlorotic to yellowish mottling and malformation were observed in leaves and collected from naturally infected fig trees of cv. Sultany in the four fig-growing province of the northern region of Egypt-Ismailia governorate (Fig. 1). Chlorotic blotching, vein clearing and banding were appeared in College of Agriculture farm (SCU). While in Al-Qantarah al-Gharbiyah farm mosaic spots were sharply delineated, the light green color of the affected areas contrasted sharply with the normal green color of the foliage. On the other hand, the leave yellowish mottling symptoms were appeared in Al-Tall al-Kabir farm. Deformed leaves showed an infinite variety of shapes and sizes were observed in Abu Sultan farm. The present study was conducted to provide information on the response of field-grown fig plant to FMV infection. Among important viruses that caused devastating losses by reducing either the yield and/or quality of fig fruit is Fig. Mosaic Virus (FMV). For this purpose, the present investigation aimed to identify unidentified isolate of this virus based on different biological and molecular tools, in order to provide a powerful diagnostic tool for early detection of FMV in infected tissues. In the present study, symptomatology identification of the FMV causes symptoms on leaves was performed. Our results were confirmed by other results obtained earlier by several investigators (Kitajima et al., 2003; Elbeaino et al., 2007; Alhudaib, 2012). Most of nursery fig trees were brought from Lebanon, Jordan and Syria. Symptoms were similar to those described by Castellano et al. (2007), chlorotic mottling, blotching, vein banding, vein clearing and malformations. Infected young leaves of potted rooted cuttings from cv. Sultany inoculated by A. ficus began to show different chlorotic ring spot, which increased in size and evolved in to a mosaic pattern. These results agreed with those obtained by Mahmoud et al. (2014).
Ultra structural features: Electron micrograph of sectioned cells revealed the presence of Double Membrane Bodies (DMBs), which are considered to be FMV particles with dimensions of 50-80 nm (Fig. 2). These results were similar to that obtained by Serrano et al. (2004). Comparable large quasi-spherical DMBs of 100~150 nm in diameter were located in the cytoplasm of parenchyma cells.
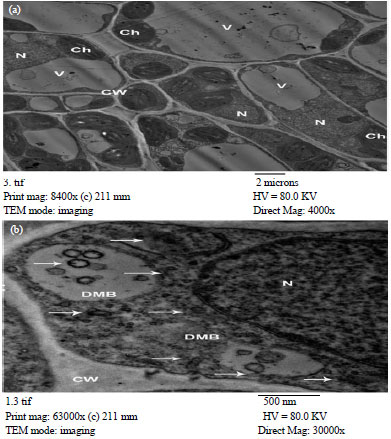 | |
| Fig. 2(a-b): | Electron micrograph of an ultra thin-section of healthy and infected fig leaves showed groups of Double-Membrane Bodies (DMB) cytoplasm of mesophyll cells from an infected fig. Ch: Chloroplast, CW: Cell wall, V: Vesicles and N: Nucleus, Bar: 500 nm, (a) Ultra thin-section of healthy fig leave and (b) Ultra thin-section of infected fig cells with FMV |
Electron dense median core particles were observed as well. Such structures were not found in PCR-negative samples to FMV, thus confirming previous reports on an etiology of DMB in mosaic affected figs (Elbeaino et al., 2009b). These structures are a consistent feature of diseased figs, regardless of the variety and the country of origin (Plavsic and Milicic, 1980; Appiano et al., 1995; Elbeaino et al., 2009c; Mahmoud et al., 2014).
RT-PCR detection: The results shown in Fig. 3 demonstrated that the amplified sequence from total RNA was readily detected in four infected samples. A major PCR product of about 302 bp amplified the RNA-dependent RNA polymerase (RdRp) gene of FMV segment = "RNA1 was present in samples-bearing FMV. Our results are in harmony with those reported for FMV by Dolja et al. (2006), Castellano et al. (2007) and Hafez et al. (2011). Amplification confirms the presence of FMV infection in this plant. Like the present observations, Alhudaib (2012) and Walia et al. (2014) also detected specific sequence of FMV in RNA extracts of infected plants by PCR. No specific product occurred for healthy material and no bands were found when the PCR assay was attempted without the initial RT step.
Sequence analysis: The RT-PCR of fig infected with FMV was purified with agarose gel using QIAquick Gel Extraction Kit from QIAGEN (Fig. 4). The RT-PCR of 302 bp FMV fragment was sequenced.
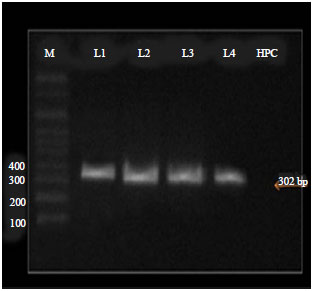 | |
| Fig. 3: | RT-PCR for fig samples tested by FMV-specific primers. M: 100 bp DNA ladder. L1, L2, L3 and L4 are four different types of fig samples infected with FMV. HPC: Healthy plant control |
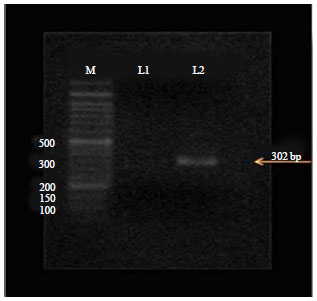 | |
| Fig. 4: | Purification of the RT-PCR product for RdRp gene. M: 50 bp DNA ladder and L2: Purified DNA sample used for sequencing |
Sequence and sequence analysis for the fragment revealed that this partial sequence belongs to the RdRp. The sequence information for the RdRp genes is very important criterion for the taxonomy of members of Bunyaviridae. Contradictory results were presented by Elbeaino et al. (2009a), who proposed that FMV was classified as genus Emaravirus, family Bunyaviridae (based on BLAST analysis of sequence from the four RNA segments). In parallel, Elbeaino et al. (2009b) suggested that FMV is a negative-sense single-stranded RNA virus belonging to the family sequenced. Sequence and sequence analysis for the fragment revealed that this partial sequence belongs to the RdRp. The sequence information for the RdRp genes is very important criterion for the taxonomy of members of Bunyaviridae.
 | |
| Fig. 5: | Sequence alignment graphic for the Egyptian isolate of FMV with other FMV sequences on the Genbank |
Contradictory results were presented by Elbeaino et al. (2009a), who proposed that FMV was classified as genus Emaravirus, family Bunyaviridae (based on BLAST analysis of sequence from the four RNA segments). In parallel, Elbeaino et al. (2009b) suggested that FMV is a negative-sense single-stranded RNA virus belonging to the family Bunyaviridae (based on BLAST analysis of sequence from the two largest RNA segments).One of our research objectives was to sequence a 302 bp fragment of the RNA-dependent RNA polymerase, RdRp gene from FMV. A multiple sequence alignment was done along with sequences previously obtained from the Genbank which have already been used as reference sequences in other studies (Ishikawa et al., 2012) (Fig. 5).
The following sequences were used in our comparisons: FMV JS1 Japan isolate with accession No. (AB697826), Canada isolates (HQ703343), Serbia (AB697827) and Italy (AM941711). The partial nucleotide sequence alignment showed 97% similarity both with FMV- JS1 Japan isolate and FMV-CAN01 Canada isolate (Fig. 6). Therefore, the Phylogenetic homology tree based on multiple sequence alignments of theFMV-Egyptian isolate, revealed that the (RdRp) of our FMV isolate was closely related to FMV-CAN01,FMV-SB1 of Canada and Serbia isolate, respectively. According to Ishikawa et al. (2012) and his co-workers, 2012, RNA1, is complementary sense gene for RdRp of FMV. The RNA2, RNA3 and RNA4 are thought, glycoprotein, NP and movement protein, respectively. The RT-PCR was found to be a reliable method in comparison with ultra structural features. Using RT-PCR the virus could be detected clearly infected in fig plants. Recently, it has been well demonstrated that RT-PCR is a more sensitive method than ELISA test to determine the occurrence of viruses in fig plants of various species.
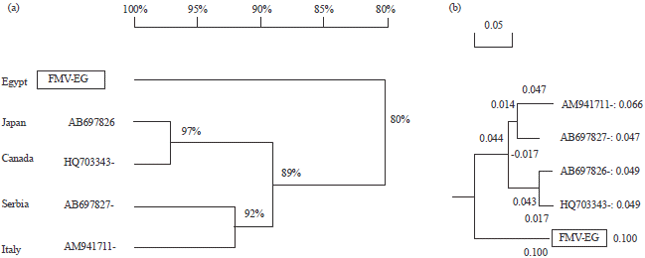 | |
| Fig. 6(a-b): | Phylogenetic analysis based on the nucleotide sequences showing the genetic relationship between the RdRp gene for RNA-dependent RNA polymerase for the Egyptian isolate of FMV-EG with other homology on the GenBank, (a) Percentage of similarity with other FMV isolates and (b) Distance matrix showing branch length and sequence weight between the different isolates |
It has also been reported that RT-PCR RdRp gene may be a useful approach for detection of the presence of FMV for screening fig samples on a large scale. Moreover sequencing of RdRp gene is often (McGavin et al., 2012) used for phylogenetic analyses of many species of the genus Emaravirus.
REFERENCES
- Ahn, K.K., K.S. Kim, R.C. Gergerich, S.G. Jensen and E.J. Anderson, 1996. Comparative ultrastructure of double membrane-bound particles and inclusions associated with eriophyid mite-borne plant diseases of unknown etiology: A potentially new group of plant viruses. J. Submicroscopic Cytol. Pathol., 28: 345-355.
Direct Link - Alhudaib, K., 2012. Incidence of fig leaf mottle-associated virus and fig mosaic virus in Eastern province of Saudi Arabia. Int. J. Virol., 8: 128-132.
Direct Link - Appiano, A., M. Conti and N. Zini, 1995. Cytopathological study of the double-membrane bodies occurring in fig plants affected by fig mosaic disease. Acta Horticult., 386: 585-592.
Direct Link - Baraket, G., K. Chatti, O. Saddoud, M. Mars, M. Marrakchi, M. Trifi and A. Salhi-Hannachi, 2009. Genetic analysis of Tunisian fig (Ficus carica L.) cultivars using Amplified Fragment Length Polymorphism (AFLP) markers. Scientia Horticulturae, 120: 487-492.
CrossRefDirect Link - Castellano, M.A., G. Gattoni, A. Minafra, M. Conti and G.P. Martelli, 2007. Fig mosaic in Mexico and South Africa. J. Plant Pathol., 89: 441-444.
Direct Link - Elair, M., N. Mahfoudhi, C. Bayoudh, I. Selmi, M. Mars and M.H. Dhouibi, 2014. Sanitary selection of virus-tested fig (Ficus carica) cultivars in Tunisia. Tunisian J. Plant Prot., 9: 100-109.
Direct Link - Elbeaino, T., M. Digiaro, A. de Stradis and G.P. Martelli, 2006. Partial characterisation of a closterovirus associated with a chlorotic mottling of fig. J. Plant Patholol., 88: 187-192.
Direct Link - Elbeaino, T., M. Digiaro, A. de Stradis and G.P. Martelli, 2007. Identification of a second member of the family closteroviridae in mosaic-diseased figs. J. Plant Pathol., 89: 119-124.
Direct Link - Elbeaino, T., M. Digiaro, A. Alabdullah, A. de Stradis and A. Minafra et al., 2009. A multipartite single-stranded negative-sense RNA virus is the putative agent of fig mosaic disease. J. Gen. Virol., 90: 1281-1288.
CrossRefPubMedDirect Link - Elbeaino, T., M. Digiaro and G.P. Martelli, 2009. Complete nucleotide sequence of four RNA segments of fig mosaic virus. Arch. Virol., 154: 1719-1727.
CrossRefDirect Link - Elbeaino, T., S. Nahdi, M. Digiaro, A. Alabdullah and G.P. Martelli, 2009. Detection of FLMaV-1 and FLMaV-2 in the Mediterranean region and study on sequence variation of the HSP70 gene. J. Plant Pathol., 91: 425-431.
Direct Link - Elbeaino, T., R.A. Kubaa, M. Digiaro, A. Minafra and G.P. Martelli, 2011. The complete nucleotide sequence and genome organization of fig cryptic virus, a novel bipartite dsRNA virus infecting fig, widely distributed in the Mediterranean basin. Virus Genes, 42: 415-421.
CrossRefDirect Link - Elbeshehy, E.K.F. and T. Elbeaino, 2011. Viruses infecting figs in Egypt. Phytopathol. Mediterr., 50: 327-332.
Direct Link - Falk, B.W. and J.H. Tsai, 1998. Biology and molecular biology of viruses in the genus Tenuivirus. Annu. Rev. Phytopathol., 36: 139-163.
CrossRefDirect Link - Gattoni, G., A. Minafra, M.A. Castellano, A. de Stradis and D. Boscia et al., 2009. Some properties of fig latent virus 1, a new member of the family flexiviridae. J. Plant Pathol., 91: 555-564.
Direct Link - Ishikawa, K., K. Maejima, Y. Komatsu, M. Kitazawa and D. Hashimoto et al., 2012. Identification and characterization of two novel genomic RNA segments of fig mosaic virus, RNA5 and RNA6. J. Gen. Virol., 93: 1612-1619.
CrossRefDirect Link - Kitajima, E.W., C.M. Chagas and J.C.V. Rodrigues, 2003. Brevipalpus-transmitted plant virus and virus-like diseases: Cytopathology and some recent cases. Exp. Applied Acarol., 30: 135-160.
CrossRefDirect Link - Martelli, G.P., M.A. Castellano and R. Lafortezza, 1993. An ultrastructural study of fig mosaic. Phytopathol. Mediterr., 32: 33-43.
Direct Link - McGavin, W.J., C. Mitchell, P.J.A. Cock, K.M. Wright and S.A. MacFarlane, 2012. Raspberry leaf blotch virus, a putative new member of the genus Emaravirus, encodes a novel genomic RNA. J. Gen. Virol., 93: 430-437.
CrossRefDirect Link - Plavsic, B. and D. Milicic, 1980. Intracellular changes in trees infected with fig mosaic. Acta Horticult., 110: 281-286.
Direct Link - Mahmoud, S.Y., S.H. Zeidan, K.A. Fayez and R. Shipat, 2014. Occurrence of fig mosaic virus in Egypt. J. Agric. Technol., 10: 439-447.
Direct Link - Serrano, L., J. Ramon, J. Segarra, V. Medina, M.A. Achon, M. Lopez and M. Juarez, 2004. New approach in the identification of the causal agent of fig mosaic disease. Acta Horticulturae, 657: 559-566.
Direct Link - Dolja, V.V., J.F. Kreuze and J.P.T. Valkonen, 2006. Comparative and functional genomics of closteroviruses. Virus Res., 117: 38-51.
CrossRefDirect Link - Walia, J.J., N.M. Salem and B.W. Falk, 2010. Partial sequence and survey analysis identify a multipartite, negative-sense RNA virus associated with fig mosaic. Plant Dis., 93: 4-10.
CrossRefDirect Link - Walia, J.J. and B.W. Falk, 2012. Fig mosaic virus mRNAs show generation by cap-snatching. Virology, 426: 162-166.
CrossRefDirect Link - Walia, J.J., A. Willemsen, E. Elci, K. Caglayan, B.W. Falk and L. Rubio, 2014. Genetic variation and possible mechanisms driving the evolution of worldwide fig mosaic virus isolates. Phytopathology, 104: 108-114.
CrossRefDirect Link - Walter, C.T. and J.N. Barr, 2011. Recent advances in the molecular and cellular biology of bunyaviruses. J. Gen. Virol., 92: 2467-2484.
CrossRefPubMedDirect Link