
A.T. Owolabi
Department of Botany, University of Calabar, PMB 1115, Calabar, Nigeria
C.O. Nwachukwu
Department of Botany, University of Calabar, PMB 1115, Calabar, Nigeria
S. Odok
Department of Botany, University of Calabar, PMB 1115, Calabar, Nigeria
International Journal of Virology
Year: 2015 | Volume: 11 | Issue: 2 | Page No.: 66-76







ABSTRACT
Cucumber is a popular fruit vegetable widely cultivated in Odukpani and Akamkpa local government areas of Cross River State, Nigeria, supplying the consumption needs of the nearby metropolitan Calabar City dwellers. However, its cultivation is threatened by infection by Papaya ringspot virus watermelon strain (PRSV-W). This study provides the result of screen house evaluation of USDA Plant Introduction (PI) accessions lines of cucumber for resistance to a Nigerian cucumber strain of PRSV-W. The response of the lines showed a spectrum of reactions to the virus strain from the extremely susceptible ones, that could not withstand the virus infection to the immune that were uninfected by the virus. Out of the 187 accession lines screened, 22 immune lines (PI 391570, PI 419010, PI 419017, PI 432873, PI 432877, PI 432878, PI 432895, PI 489752, PI 137845, PI 211979, PI 211984, PI 605924, PI 606010, PI 227207, PI 390253, PI 223437, PI 118279, PI 422200, PI 525152, PI 406473, PI 390952 and PI 357867) were identified. Besides, 17 others (PI 118807, PI 163217, PI 165509, PI 175121, PI 177364, PI 179263, PI 212985, PI 215589, PI 217644, PI 220791, PI 234517, PI 249562, PI 267746, PI 271326, PI 344384, PI 358813 and PI 358814) showed high tolerant and eight others were moderately tolerant to the virus. The remaining 140 PI lines were very susceptible to the virus. The identification of completely immune and highly tolerant PI accession cucumber lines in this study points the availability at the USDA germ plasm collections of potential materials that could be included in breeding programs to develop high resistant varieties of cucumber to PRSV-W.
PDF Abstract XML References Citation
Received: February 17, 2015;
Accepted: May 25, 2015;
Published: August 04, 2015
How to cite this article
A.T. Owolabi, C.O. Nwachukwu and S. Odok, 2015. Screening Cucumber Plant Introduction Accession Lines for Resistance against Cucumber Strain of Papaya ringspot virus (PRSV). International Journal of Virology, 11: 66-76.
DOI: 10.3923/ijv.2015.66.76
URL: https://scialert.net/abstract/?doi=ijv.2015.66.76
DOI: 10.3923/ijv.2015.66.76
URL: https://scialert.net/abstract/?doi=ijv.2015.66.76
INTRODUCTION
Cucumis sativus L. (Cucurbitaceae), commonly known as cucumber, is believed to have originated in India (Renner et al., 2007), where a great many varieties have been observed and has now been introduced to other parts of the world. It is cultivated for its fruits which are eaten fresh or used in the preparation of sandwiches and salads. It is also reported to be a rich source of phytonutrients such as vitamins B1, C, K and essential minerals (Abiodun and Adeleke, 2010). Cucumber varieties have also been reported to provide valuable antioxidant, anti-inflammatory and anti-cancer substances in the diet (Lee et al., 2010; Kumar et al., 2010; Nema et al., 2011).
Papaya ringspot virus (PRSV), previously described as Watermelon mosaic virus-1 (WMV-1), (Purcifull et al., 1984) is a potyvirus transmitted by a number of aphid species in a style-borne manner (Gonsalves and Ishii, 1980) and considered a major production constraint to papaya and cucurbit production worldwide (Gonsalves et al., 2010; Mohammed et al., 2012). Two biotypes are recognized, the papaya infecting biotype (PRSV-P) which infects papaya and cucurbits and the PRSV-W that infects only cucurbits (Tripathi et al., 2008). PRSV-W which has been reported to as a major limiting factor for watermelon production worldwide (Guner et al., 2002; Strange et al., 2002), has been found to be transmitted in a stylet-borne manner by 24 aphid species in 15 genera with Myzus persicae, Aulacorthum solani, Aphis craccivora and Macrosiphum euphorbiae as natural vectors (Purcifull et al., 1984). Reports of the occurrence of PRSV in cucurbits from Africa abound. The PRSV-P has been reported on Cucurbita pepo in Egypt (Omar et al., 2011), on C. maxima in Sudan (Mohammed et al., 2012), in Carica papaya in Nigeria (Taylor, 2001) and Cote d’Ivoire Diallo et al. (2007) while the PRSV-W biotype has been reported in cucumber in Nigeria (Owolabi et al., 2008).
From the review of literature, resistant accessions have been identified in several crop germ plasm against viruses of economic significance. These include cowpea (Bashir et al., 2002), okra (Rashid et al., 2002), soybean (Arif and Hassan, 2002), watermelon (Strange et al., 2002; Guner, 2004), bottlegourd (Ling and Levi, 2007), urbean (Ashafaq et al., 2007), cotton (Ahmad et al., 2010) and potato (Ahmad et al., 2011; Batool et al., 2011). Others include mungbean (Iqbal et al., 2011; Mondol et al., 2013), tomato (Imran et al., 2012; Osei et al., 2012) and chilli pepper (Ashafaq et al., 2014).
The cultivation of C. sativus has become a thriving business in Odukpani and Akamkpa local government areas of Cross River State, Nigeria, providing jobs for the local folks and meeting the consumption needs of the nearby metropolitan Calabar city dwellers. However, all the local varieties available for cultivation are susceptible to a PRSV-W strain isolated from cucumber in which it elicited mosaic, leaf malformation, rugosity, conspicuous green vein-banding and stunting (Owolabi et al., 2008).
Several approaches have been adopted for the management or control of virus diseases of plants. These include cross protection (Gonsalves, 2004; Zhou and Zhou, 2012), interference with vector activities (Murphy et al., 2009), obtaining virus-free plant materials through meristem culture (Shiragi et al., 2008; El Far and Ashoub, 2009), thermotherapy (Arif et al., 2005; Panattoni et al., 2013), coat protein mediated resistance (Bendahmane et al., 2007; Srivastava and Raj, 2008; Mehta et al., 2013) and the use of transgenic lines (Yu et al., 2011).
Although, some of these approaches may make valuable contributions to virus disease management in specific favourable situations, they sometimes are of little practical benefits. Planting resistant varieties, in many cases, appears to offer a more practical option in controlling plant virus diseases. The aim of the present study was to evaluate 187 accession lines of C. sativus with a view to identifying resistant lines that could provide genetic material for breeding resistance against PRSV-W in Nigeria.
MATERIALS AND METHODS
Source of accession lines: A total of 187 accession lines of C. sativus var. sativus, [except C. sativus var. hardwickii (PI 215589)], were sourced from the United States Department of Agriculture, Agriculture Research Station (USDA-ARS), Iowa State University Regional Plant Introduction Station and certified virus-free by Plant Germ plasm Quarantine Centre, Beltville, Maryland (USA). The accessions lines had their origin from Africa, Asia, Europe and the Americas (33 countries in all). Countries with most accessions included India with 40, China 25, Turkey 17, Japan 16 and Iran 13. Other had less than 10 or fewer.
PRSV virus isolate: The PRSV-W strain used in the study was that described by Owolabi et al. (2008) and maintained on Cucumeropsis manni (Naudin) in the screen house.
Planting and inoculation procedure: Five seeds of each of the accession lines were sown in 20×16 cm polyethylene bags containing heat-sterilized garden soil. The cotyledonary leaves (8-day old seedlings), pre-dusted with 600-mesh carborundum were mechanically inoculated with the PRSV strain in 0.03 mM Na2HSO4, pH 8.00 inoculation buffer. Leaf to buffer ratio was 1:5 (1 g infected leaf to 5 mL of buffer). Three buffer-inoculated seedlings of each accession line served as controls.
The inoculated seedlings were promptly rinsed with water, kept in an insect-proof screen house at 27±°C in the University’s Botanical Garden and left for symptom development for over a period of 21 days. Plants that did not show symptoms of infection were back-indexed on C. manni that had been reported to be readily susceptible to the virus (Owolabi et al., 2008) to check for latent infection.
Disease rating/determination of degree of susceptibility to PRSV: The germ plasm accession lines were evaluated against their response to the virus on a scale of 0-4 according to Bashir et al. (2002) with some modifications on the basis of viral symptoms where, 0 = immune (inoculated plants did not show symptoms and no virus recovered after back-indexing, 1 = highly tolerant (inoculated plant showed mild mottle and no apparent reduction in plant growth), 2 = moderately tolerant (inoculated plant showed mosaic, green vein-banding, leaves not malformed and no apparent reduction in leaf size), 3 = susceptible (stunting of inoculated plants, leaf malformed and accompanied by reduction in leaf size) and 4 = highly susceptible (apical necrosis leading growth cessation).
RESULTS
Reaction of accession lines to PRSV: The reaction of the cucumber accession lines to PRSV inoculation showed a spectrum of responses from complete lack of symptom induction to severe disease reactions (Fig. 1a-i and Table 1) and in some cases, growth cessation occasioned by shoot tip necrosis and eventual death of inoculated seedlings. Some of the symptoms observed included severe leaf malformation and reduction in leaf size (Fig. 1a-e), rugosity (blistering), (Fig. 1f) mosaic and green-vein banding and (Fig. 1h) no symptom at all (Fig. 1i).
Disease rating: Five categories of accessions lines were identified based on the criteria earlier defined. These were the immune lines, the highly tolerant, the moderately tolerant, the susceptible and the highly susceptible lines. The results showed that a significant number of the accession lines, 128 out of the 187, belonged to the susceptible category, representing about 68.45% of the total accession lines screened (Table 2) while, 22 (about 11.76%) were adjudged immune.
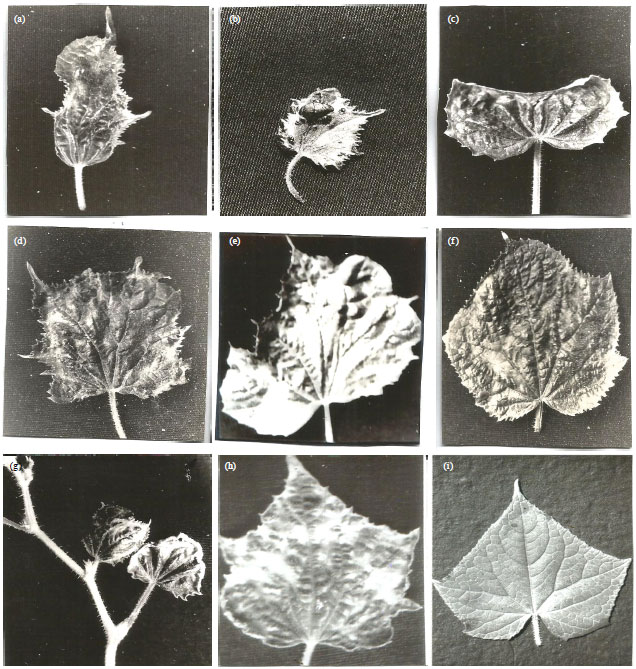 | |
| Fig. 1(a-i): | Symptoms elicited in some accession lines of Cucumis sativus var. sativus to inoculation with Nigerian cucumber isolate of Papaya ringspot virus, (a) (PI 137856, Iran), (b) (PI 285607, Poland), (c) (PI 176523, Turkey), (d) (PI 169395, Turkey), (e) ( PI 271327, India) showing various forms of leaf malformations, (f) (PI 483342, China) showing rugosity while, (g) (PI 220338, Afghanistan), (h) (PI 432891, China) exhibited mosaic and green-vein banding and (i) Shows healthy control |
Out of the 22 immune lines eight (PI 391570, PI 419010, PI 419017, PI 432873, PI 432877, PI 432878, PI 432895 and PI 489752) were of Chinese origin, three (PI 137845, PI 211979 and PI 211984) from Iran, two each from India (PI 605924 and PI 606010) and Japan (PI 227207 and PI 390253) and one each from Afghanistan (PI 223437), Brazil (PI 118279), Czech Republic (PI 422200), Egypt (PI 525152), Netherlands (PI 406473), Russia Federation (PI 390952) and Yugoslavia (Slovania) (PI 357867).
| Table 1: | Accession lines, countries of origin and reactions to Nigerian cucumber strain of Payaya rinspot virus countries of origin |
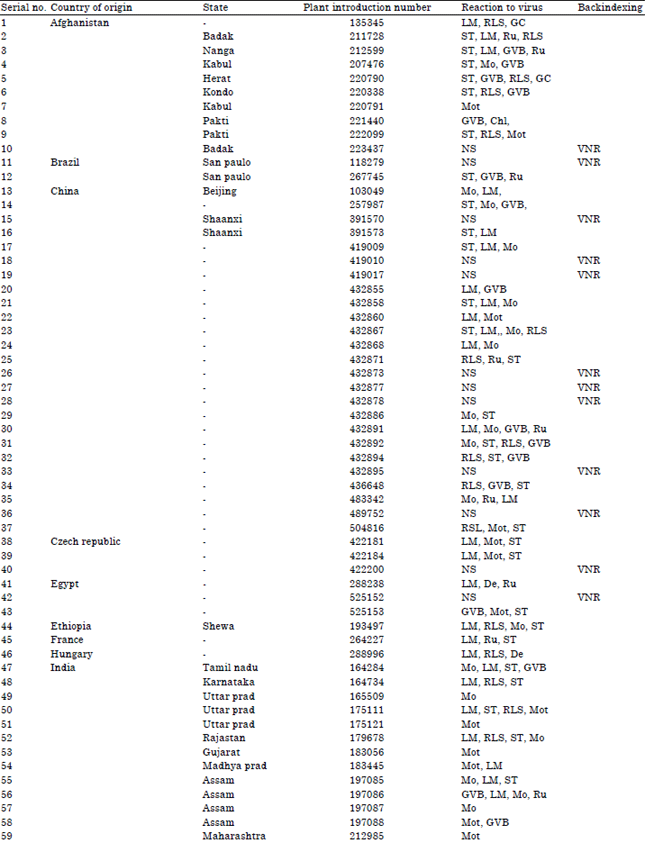 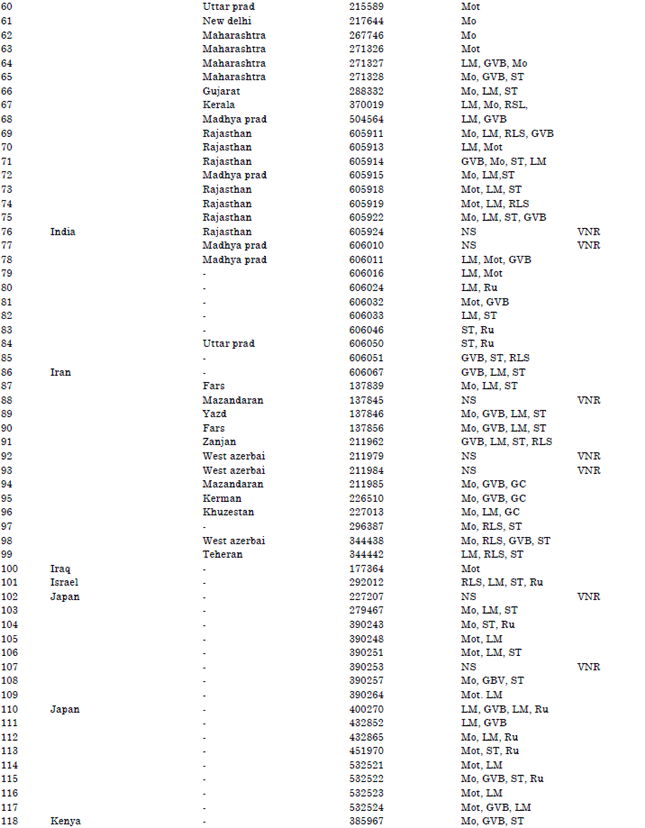 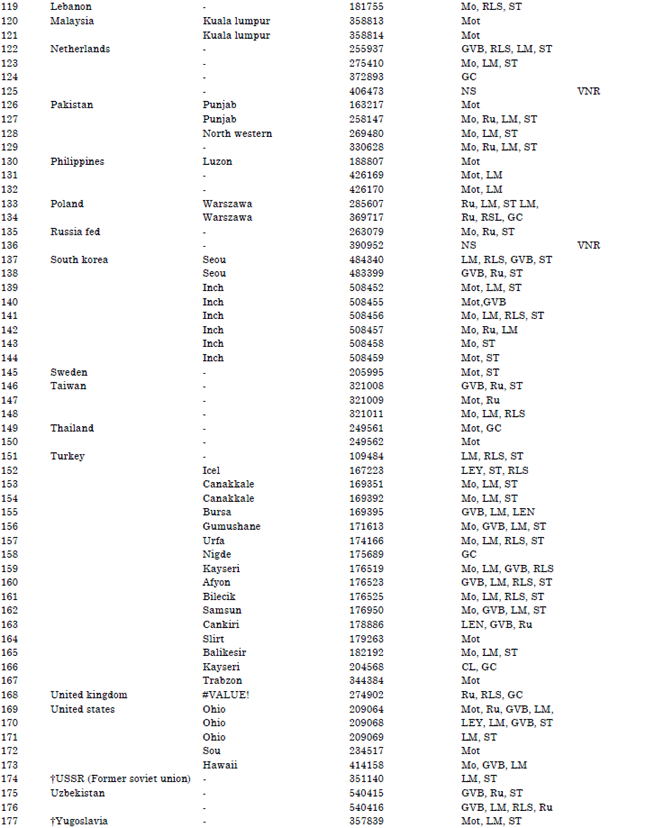 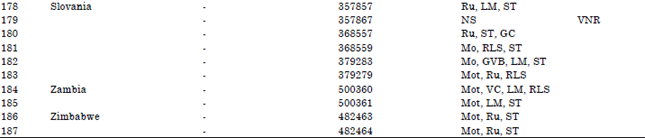 | |
| GC: Growth cessation, LM: Leaf malformation, RLS: Reduced leaf size, Ru: Rugosity, GVB: Green vein-banding, Mo: Mosaic, Mot: Mottle, Chl: Chlorosis, NS: No symptoms, De: Defoliation, VC: Veinal chlorosis, LEY: Leaf edge yellowing, LEN: Leaf edge necrosis and VNR: Virus not recovered, | |
| Table 2: | Grouping of cucumber germ plasm accession lines tested against Nigerian cucumber strain of Papaya ringspot virus |
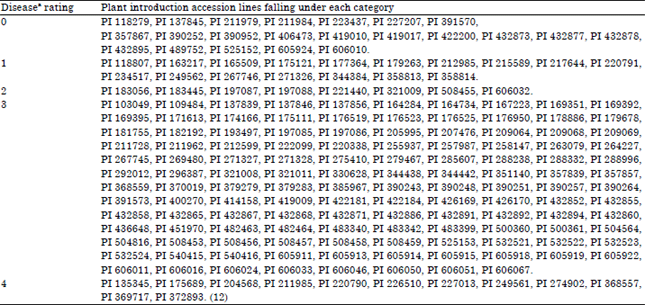 | |
| aDisease rating: 0: Immune, 1: Highly tolerant, 2: Moderately tolerant, 3: Susceptible and 4: Highly susceptible | |
The results also showed that 12 ( PI 135345, PI 175689, PI 204568, PI 211985, PI 220790, PI 226510, PI 227013, PI 249561, PI 274902, PI 368557, PI 369717 and PI 372893) were highly susceptible to the virus as they suffered from growth cessation of the apical bud. The remaining 25 (about 13.37%) were found to be tolerant (both moderate and highly).
DISCUSSION
Cucumis sativus is an important fruit vegetative in Cross River, Nigeria. All available varieties under cultivation are readily susceptible to the PRSV-W strain, with concomitant poor growth. In this study, 187 USAD-ARS Plant Introductions of C. sativus var. sativus were screened against PRSV-W isolate from Nigeria for possible discovery of varieties that could provide materials for resistance breeding programme.
The results of this investigation showed a varying degree of reactions to the cucumber isolate of PRSV-W among Cucumis sativus var. sativus germ plasm collection from the USDA-ARS. A preponderance (about 67.02%) of the accession lines was susceptible to the virus. Similar observations have been made with respect to some cucurbits screened against viruses of economic importance. Strange et al. (2002) found that a greater majority of 1248 accession lines of watermelon were susceptible to PRSV-W. The report of Ling and Levi (2007) also showed that 90 (47.37%) of the 190 screened were susceptible to the Florida strain of ZYMV and Kousik et al. (2009) did not detect resistance to Squash yellowing vein virus (SqVYV) among PIs 218 watermelon accession lines obtained from USDA germplasm collection as all were susceptible to the virus with varying degree of reactions.
Germ plasm collections have become veritable sources of genetic materials for possible breeding programmes against plant viruses. The result of this study demonstrated that no fewer than 22 were completely immune to the PRSV-W strain. These were PI 391570, PI 419010, PI 419017, PI 432873, PI 432877, PI 432878, PI 432895 and PI 489752 of Chinese origin, three (PI 137845, PI 211979 and PI 211984) from Iran, two each from India (PI 605924 and PI 606010), Japan (PI 227207 and PI 390253) and one each from Afghanistan (PI 223437), Brazil (PI 118279), Czech Republic (PI 422200), Egypt (PI 525152), Netherlands (PI 406473), Russia Federation (PI 390952) and Yugoslavia (Slovania) (PI 357867). Completely immune and moderately resistance PI accession lines have been detected among some cucurbit germ plasm sourced from USDA. Strange et al. (2002) reported the existence of PRSV-W resistance in eight PI accessions: three accessions from South Africa (PI 244017, PI 244018 and PI 244019), in three accessions from Zimbabwe (PI 482342, PI 482318 and PI 482379), one accession from Botswana (PI 485583) and one accession from Nigeria (PI 595203) in watermelon germ plasm collection from the USDA. Ling and Levi (2007) also found that of the 190 Lagenaria siceraria PIs screened, 30 were completely immune to the Florida strain of ZYMV while Kousik et al. (2009) reported the existence of moderate resistance in two C. colocynthis (PI 386015 and PI 386024), a Praecitrullus fistulosus (PI 381749) and two C. lanatus var. lanatus PIs (PI 482266 and PI 392291 to Squash vein yellowing virus (SqVYV). On the other hand, Habib et al. (2007) reported that none of 254 lines of mungbean was found to be resistant to MYMV while 247 lines were highly susceptible to the virus.
The USDA watermelon and other cucurbit germ plasm collections have been extensively screened for resistance to PRSV-W and ZYMV. This is the first report of screening cucumber accessions line for resistance against PRSV-W.
CONCLUSION
The result of the present study demonstrates that there is significant genetic resistance to PRSV-W among USDA C. sativus var. sativus germ plasm collections. The twenty two accessions so identified are potential sources of genetic materials for cucumber breeding against the virus.
ACKNOWLEDGMENTS
The authors are grateful to USDA for graciously supplying the accession lines for the research.
REFERENCES
- Abiodun, O.A. and R.O. Adeleke, 2010. Comparative studies on nutritional composition of four melon seeds varieties. Pak. J. Nutr., 9: 905-908.
CrossRefDirect Link - Ahmad, S., N.A. Mahmood, F. Ashraf, K. Hayat and M. Hanif, 2010. Screening of cotton germplasm against Cotton leaf curl virus. Pak. J. Bot., 42: 3327-3342.
Direct Link - Ashafaq, M., S. Iqbal, T. Muktar and H. Shall, 2014. Screening for resistance to Cucumber mosaic virus in chilli pepper. J. Anim Plant Sci., 24: 791-795.
Direct Link - Arif, M. and S. Hassan, 2002. Evaluation of resistance in soybean germplasm to Soybean mosaic potyvirus under field conditions. J. Biol. Sci., 2: 601-604.
Direct Link - Arif, M., M. Ibrahim, A. Ahmad and S. Hassan, 2005. Elimination of citrus tristeza closterovirus from citrus bud-wood through thermotherapy. Pak. J. Bot., 37: 423-430.
Direct Link - Bashir, M., Z. Ahmad and A. Ghafoor, 2002. Cowpea germplasm evaluation for virus resistance under greenhouse conditions. Asian J. Plant Sci., 1: 585-587.
CrossRefDirect Link - Bendahmane, M., I. Chen, S. Asurmendi, A.A. Bazzini, J. Szecsi and R.N. Beachy, 2007. Coat protein-mediated resistance to TMV infection of Nicotiana tabacum involves multiple modes of interference by coat protein. Virology, 366: 107-116.
CrossRefDirect Link - Diallo, H.A., W. Monger, N. Kouassi, D.T. Yoro and P. Jones, 2007. First report of Papaya ringspot virus infecting papaya in Cote d'Ivoire. Plant Pathol., 56: 718-718.
CrossRefDirect Link - Gonsalves, D. and M. Ishii, 1980. Purification and serology of Papaya ring spot virus. Phytopathology, 70: 1028-1032.
Direct Link - Gonsalves, D., S. Tripathi, J.B. Carr and J.Y. Suzuki, 2010. Papaya ring spot virus. The Plant Health Instructor.
Direct Link - Guner, N., E.B. Strange, T.C. Wehner and Z. Pesic-VanEsbroeck, 2002. Methods for screening watermelon for resistance to papaya ringspot virus type-W. Scientia Hortic., 94: 297-307.
CrossRefDirect Link - Habib, S., N. Shad, A. Javaid and U. Iqbal, 2007. Screening of mungbean germplasm for resistance/tolerance against yellow mosaic disease. Mycopath, 5: 89-94.
Direct Link - Kumar, D., S. Kumar, J. Singh, B.D. Vashistha and N. Singh, 2010. Free radical scavenging and analgesic activities of Cucumis sativus L. fruit extract. J. Young Pharm., 2: 365-368.
CrossRefDirect Link - Kousik, C.S., S. Adkins, W.W. Turechek and P.D. Roberts, 2009. Sources of resistance in US plant introductions to watermelon vine decline caused by squash vein yellowing virus. HortScience, 44: 256-262.
Direct Link - Lee, D.H., G.B. Iwanski and N.H. Thoennissen, 2010. Cucurbitacin: Ancient compound shedding new light on cancer treatment. Sci. World J., 10: 413-418.
CrossRef - Ling, K.S. and A. Levi, 2007. Sources of resistance to Zucchini yellow mosaic virus in Lagenaria siceraria germplasm. HortScience, 42: 1124-1126.
Direct Link - El Far, M.M.M. and A. Ashoub, 2009. Utility of thermotherapy and meristem tip for freeing sweetpotato from viral infection. Aust. J. Basic Applied Sci., 3: 153-159.
Direct Link - Mehta, R., T. Radhakrishnan, A. Kumar, R. Yadav and J.R. Dobaria et al., 2013. Coat protein-mediated transgenic resistance of peanut (Arachis hypogaea L.) to peanut stem necrosis disease through Agrobacterium-mediated genetic transformation. Indian J. Virol., 24: 205-213.
CrossRefDirect Link - Mohammed, H., A. Manglli, S. Zicca, A. El-Hussein, M. Mohammed and L. Tomassoli, 2012. First report of Papaya ringspot virus in pumpkin in Sudan. New Dis. Rep., 26: 26-26.
Direct Link - Murphy, J.F., M.D. Eubanks and J. Masiri, 2008. Reflective plastic mulch but not a resistance-inducing treatment reduced Watermelon mosaic virus incidence and yield losses in squash. Int. J. Veg. Sci., 15: 3-12.
Direct Link - Osei, M.K., R. Akromah, J.N.L. Lamptey and M.D. Quain, 2012. Phenotypic and molecular screening of some tomato germplasm for resistance to tomato yellow leaf curl virus disease in Ghana. Afr. J. Agric. Res., 7: 4675-4684.
Direct Link - Omar, A.F., S.A. El-Kewey, S.A. Sidaros and A.K. Shimaa, 2011. Egyptian isolates of Papaya ringspot virus form a molecularly distinct clade. J. Plant Pathol., 93: 569-576.
Direct Link - Panattoni, A., A. Luvisi and E. Triolo, 2013. Review. Elimination of viruses in plants: Twenty years of progress. Spanish J. Agric. Res., 11: 173-188.
CrossRefDirect Link - Rashid, M.H., L. Yasmin, M.G. Kibria, A.K.M.S.R. Mollik and S.M.M. Hossain, 2002. Screening of okra germplasm for resistance to yellow vein mosaic virus under field conditions. Plant Pathol. J., 1: 61-62.
CrossRefDirect Link - Renner, S.S., H. Schaefer and A. Kocyan, 2007. Phylogenetics of cucumis (Cucurbitaceae): cucumber (C. sativus) belongs in an Asian/Australian clade far from melon (C. melo). BMC Evol. Biol., Vol. 7.
CrossRefDirect Link - Shiragi, M.H., M.A. Baque and K.M. Nasiruddin, 2008. Eradication of Banana Bunchy Top Virus (BBTV) and Banana Mosaic Virus (BMV) from infected plant of banana cv. Amritasagar through Meristem culture. South Pac. Stud., 29: 17-41.
Direct Link - Strange, E.B., N. Guner, Z. Pesic-VanEsbroeck and T.C. Wehner, 2002. Screening the watermelon germplasm collection for resistance to Papaya ringspot virus type-W. Crop Sci., 42: 1324-1330.
CrossRefDirect Link - Tripathi, S., J.Y. Suzuki, S.A. Ferreira and D. Gonsalves, 2008. Papaya ringspot virus‐P: characteristics, pathogenicity, sequence variability and control. Mol. Plant Pathol., 9: 269-280.
CrossRefDirect Link - Yu, T.A., C.H. Chiang, H.W. Wu, C.M. Li and C.F. Yang et al., 2011. Generation of transgenic watermelon resistant to Zucchini yellow mosaic virus and Papaya ringspot virus type W. Plant Cell Rep., 30: 359-371.
CrossRefDirect Link