
A.A.J. Mofunanya
Department of Botany, Faculty of Science, University of Calabar, Calabar, Nigeria
A. Towolabi
Department of Botany, Faculty of Science, University of Calabar, Calabar, Nigeria
A. Nkang
Department of Botany, Faculty of Science, University of Calabar, Calabar, Nigeria
International Journal of Virology
Year: 2015 | Volume: 11 | Issue: 2 | Page No.: 54-65







ABSTRACT
The effect of Telfairia Mosaic Virus (TeMV) on the growth characteristics of two ecotypes of Telfairia occidentalis was investigated. Seeds of T. occidentalis obtained from Akparabong in Ikom Local Government Area of Cross River State were designated ecotype A and seeds from Eket in Eket Local Government Area of Akwa Ibom State were designated ecotype B. Infection of T. occidentalis by TeMV caused significant reductions (p<0.05) in leaf area, shoot height, leaf number, leaf fresh and dry weight, fresh and dry weight of shoot and root. Percentage difference values for the above parameters were 57.19, 40.10, 46.59, 25.0, 24.5, 31.65, 37.43, 30.78, 19.97, 37.01 and 21.94% for ecotype A and B in infected samples compared to healthy controls. The virus also induced (p<0.05) reductions in Relative Growth Rate (RGR), with percentage difference values for ecotype A and B ranging from 21.1-21.7, 17.5 and 28.2%, with values for Net Assimilation Rate (NAR) for ecotype A and B ranging from 22.1-39.3 and 20.5-34.4%, respectively and percentage difference values for Leaf Area Ratio (LAR) for ecotype A and B ranging from 13.9-26.0 and 15.0-25.3%, respectively. Telfairia mosaic virus infection caused significant (p<0.05) reductions in RGR, NAR and LAR in inoculated plants of ecotypes studied compared to the healthy controls with ecotype A being more susceptible to the effect of TeMV infections than ecotype B. The virus also reduced the quality of the vegetable. Since, the difference in plant growth between infected and healthy plants depend on infection, this explains the difference in growth and plant size. These reductions in plant growth characteristics induced by TeMV infection alter the metabolism and subsequently dry matter yield of T. occidentalis.
PDF Abstract XML References Citation
Received: March 11, 2015;
Accepted: July 10, 2015;
Published: August 04, 2015
How to cite this article
A.A.J. Mofunanya, A. Towolabi and A. Nkang, 2015. Comparative Study of the Effect of Telfairia Mosaic Virus (TEMV) on the Growth Characteristics of Two Ecotypes of Telfairia occidentalis (Hooker Fil). International Journal of Virology, 11: 54-65.
DOI: 10.3923/ijv.2015.54.65
URL: https://scialert.net/abstract/?doi=ijv.2015.54.65
DOI: 10.3923/ijv.2015.54.65
URL: https://scialert.net/abstract/?doi=ijv.2015.54.65
INTRODUCTION
Telfairia occidentalis Hooker fil is a member of Cucurbitaceae family. Telfairia occidentalis play an important role in traditional medical practice and local diets of the South Eastern states of Nigeria, improve households income generation, broaden food base as it supplies the body with minerals, vitamins, hormones precursors and fibre (energy) micronutrients (Akubugwo et al., 2008).
The ecotype is often used in literature to describe a locally adapted population of a widespread species. Such populations show minor changes of morphology and or physiology, which are related to habitat and are genetically induced. Nevertheless, they can still reproduce with other ecotypes of the same species (IPN., 2004). Ecotype is a distinct form of plant species occupying a particular habitat. Ecotype in others biological literatures refers to populations that have evolved in an extreme environment such as soil with unusual mineral concentrations or a typical geographical area (IPN., 2004). The aggregate effect is the evolution of different populations of the same plant species with distinctive characteristics.
Viruses are considered to be of economic importance only when they cause some deviation from normal growth of plant, such as reduction in plant height, size of leaves, flowers and roots and shortening of petioles and internodes (El-Dougdoug et al., 2007). Virus particle multiplication in virus infected plant cells exhibit some physiological changes (Afreen et al., 2011). Significant, insignificant and no reductions due to virus infections have been documented. Stobbs et al. (1991) reported that there was no significant difference in the dry matter content of healthy and infected roots of Rutabaga due to Turnip mosaic virus infection at different stages of development. Takahashi (1972) documented that 100 days after infection, tobacco plants infected with TMV possessed about one leaf less than healthy plants. Careful analysis of leaf initiation following inoculation of very young Chinese cabbage plants with TYMV showed a rapid transient reduction in the rate of leaf initiation. Funayama et al. (1997) observed lower growth rate as a result of virus infection. Reduction in leaf numbers was reported as a very small factor in the stunting of herbaceous plant following virus infection. Garud and Mali (1980), Vyavahare (1981) and Pawar et al. (1990) reported that infection sorghum with SRSV significantly reduced the plant height and leaf area. Funayama et al. (2001) documented that stem growth of Eupatorium makinoi was significantly suppressed by infection and also flowering and survivorship of infected plants decreased with reducing plant height. Booker et al. (2005) in their study recorded maximum reduction in leaf area index, leaf area duration and pod yield in cowpea infected by Cowpea severe mosaic virus. Kazinczi et al. (2006) reported reduction in both fresh and dry weight of S. nigrum due to PepMV and ObPV. El-Dougdoug et al. (2007) studied the influence of Potato Virus Y (PVY) on the growth of Datura metel. They reported reductions in number of leaves, plant height, leaf area, fresh and dry weight of shoots fresh and dry weight of D. metal was negatively influenced by PVY infection.
Rahman and Akanda (2010) documented that the higher rate of PLRV in potato caused significant increase in disease incidence and reduction in plant height, stem number, tuber number, tuber yield as compared to that in control. Ehinmore and Kareem (2010) reported that infection of Amaranthus hybridus by AMV produced significant reduction in plant heights of three Amaranthus cultivars: NHAM/114, NH84/457-IL and commercial by 44, 34 and 37%, respectively. The virus also resulted in significant reduction in the number of leaves in all the three cultivars.
Hooks et al. (2008) reported significant reductions petiole length and distance, pseudo-stem diameter plant height and canopy and leaf area in banana plants infected with banana bunchy top virus.
The growth of stimulated swards of Italian and perennial ryegrass artificially infected with RMV or RMV-free was investigated by Jones et al. (1977). They reported that during the build up of RMV infection, dry matter yield was usually decreased in infected swards.
Kareem et al. (2011) stated greater reduction in fresh and dry weight of leaf and stem of Amaranthus hybridus due to infection by Amaranthus mosaic virus. Reduction in vegetative growth parameters, RWC, chlorophyll a and b, plant fresh and dry weight of different plant parts due to TMV infection have been reported by Pazarlar et al. (2013).
The objective of this study was to compare the effect of Telfairia mosaic virus infection on the growth characteristic of two ecotypes of Telfairia occidentalis (Hooker fil).
MATERIALS AND METHODS
Collection of seeds and planting: Fruits of T. occidentalis from which seeds were obtained were purchased from local farmers in two localities: Ikom (CRS) and Eket (AKS) and grown in Calabar, Nigeria. For the purpose of this work, the seeds obtained from Ikom (CRS) were designated as ecotype A and those from Eket (AKS) as ecotype B. Ikom (CRS) located at latitutde 5°57’ N and longtitude 8°44’ E is a derived savanna zone and fruits were collected from Akparabong town in Ikom Local Government Area (LGA). With total monthly rainfall of 241.65 mm, mean monthly temperature of 31.83°C and mean monthly relative humidity of 81.13% for the year under study.
Akwa Ibom State (AKS) is a rainforest zone and fruits were collected from Eket municipality, Eket LGA located at latitutde 4°37’ N and longitude 7°55’ E. Total monthly rainfall of 326.08 mm, mean monthly temperature of 28.63°C and mean monthly relative humidity of 85.82% (Table 1).
The fruits and seeds of both ecotypes were distinguished by their general morphology. The seeds of ecotype A were generally smaller measuring about 3.0 cm (length/width), fruit pod of 25/16 cm length/width, large and greenish leaves. The seeds of ecotype B measured about 3.5 cm (length/width). The fruit pod of 30/17 cm (length/width), leaf size was larger and leaf colour more greenish than those of ecotype A.
The seeds of these ecotypes were removed and sorted in order to select for uniformity of size. They were sun-dried for two days to enhance germination and thereafter sown in steam-sterilized garden soil in 16 cm diameter polyethylene bags. The seeds were planted in the greenhouse in the Botanical Garden of the University of Calabar. The seeds germinated 14 days after planting and the germinated seeds (seedlings) were staked to promote adequate leaf production and to aid measurement of the shoot height and leaf number count.
Virus source, preparation of inoculum and virus propagation: Telfairia mosaic virus used in this study was obtained from the Federal Biological Research Centre for Agriculture and Forestry, (BBA) Braunschweig, Germany. The TeMV-infected leaf tissues were stored under liquid nitrogen and re-activated by triturating TeMV-infected leaf tissues in pre-sterilized cold pestle and mortar in sodium phosphate (Na2HPO4) buffer 0.03 M, pH 8.0. The inoculum was applied by the conventional leaf-rub method (mechanical or sap inoculation) on Nicotiana benthamiana, with 800- mesh carborundum. The leaves were rinsed with water and observed for symptom expression. Subsequent inoculations using the sap transmission method were carried out on T. occidentalis for propagation and maintenance of the virus under greenhouse condition at 25±3°C.
Experimental design: The plants from the two localities were arranged in two groups each containing 30 plants each set out in three rows of ten plants.
| Table 1: | Rainfall, temperature and relative humidity data in Ikom (CRS) and Eket (AKS) in the year 2007 |
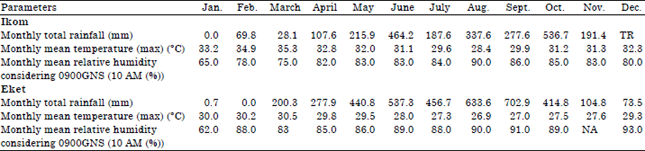 | |
| Source: Nigerian Meteorological Agency, Calabar Airport, TR: Trace, N/A: Not Applicable | |
Before inoculation, plants which were to be inoculated with the virus and those of controls in both groups were kept in a Randomized Block Design (RBD) in the greenhouse. The inoculation of the test plants was as described above. Within each plant groups, fifteen were inoculated with the virus and the remaining fifteen inoculated with buffer to serve as controls. The set up was observed for symptom development (10-14 days post-inoculation) such as mosaic, severe leaf malformation and distortion characteristics of TeMV infection of T. occidentalis.
Effect of TeMV infection on plant growth characteristics: Assessment was carried out on infected and healthy plants of the two ecotypes of T. occidentalis to determine the effect of TeMV on morphology and physiological functions. Parameters determined included leaf morphology, leaf number, shoot height, leaf area, leaf fresh and dry weight and fresh and dry weight of shoot and root. Growth analysis was carried out to determine the Relative Growth Rate (RGR), Net Assimilation Rate (NAR) and the Leaf Area Ratio (LAR).
Effect of TeMV infection on leaf area: Estimate was carried out using leaves of the same age and position in infected and healthy plants of both ecotypes. In this method, the leaf of each ecotype was placed on a 1 mm2 graph paper. The leaf size was traced on the paper and the total area calculated based on the number of squares covered within the traced region. Initial measurement of leaf size was taken at 14 Days Post-Inoculation (DPI), subsequent measurement was carried out at 30 days interval for a period of 120 days in infected and healthy leaves of both ecotypes:
![]()
Effect of TeMV infection on shoot height and number of leaves: The effect of viral inoculation on shoot height was determined by measuring stem length in centimeters from soil level to the tip of the stem for infected and healthy plants of both ecotypes with the aid of a measuring tape. Measurements were taken for five plants each at 14, 30, 60, 90 and 120 DPI. The leaves of the inoculated plants and healthy controls for each ecotype were counted to determine the effect of the virus infection on the number of leaves over the same period of time.
Effect of TeMV infection on leaf fresh and dry weight: The leaves were harvested at 30 DPI from five plants of each ecotype. Leaf fresh weight was taken for each sample. Leaf dry weight was determined by drying leaf samples at temperatures of 70°C for 24 h. Samples were dried and weighed for three times and the mean of three readings were taken as correct value (Miyoshi et al., 1996). The procedure was carried out for infected and healthy.
Effect of TeMV infection on fresh and dry weight of shoot and root: After a period of 120 days, five plants were uprooted. In order to prevent some portion of the roots from breaking off, the plants were placed in a bucket of water and the soil gently washed off. The shoots were cut off from the roots with scissor and the fresh weight of both the shoots and roots were recorded. The shoots and roots were then dried in the hot-air oven (Gallenkamp, England) at 74°C for 24 h. Samples were dried as described above (Miyoshi et al., 1996).
Effect of TeMV infection on Relative Growth Rate (RGR), Net Assimilation Rate (NAR) and Leaf Area Ratio (LAR): The RGR measures the change in weight (dw) of different samples of T. occidentalis from the two localities. The change in weight (dt) was taken at 14, 30, 60, 90 and 120 DPI. Differences were calculated between the healthy and infected leaves for both ecotypes using the equation given below (Atwell et al., 1999):
![]()
Net assimilation rate is a function of the photosynthetic efficiency of leaves of a plant and its leafiness. The difference in the natural logarithm of leaf area over leaf area and leaf dry weight over difference in time was used to show the photosynthetic efficiency between the healthy and infected leaves of both ecotypes:
![]()
The LAR represents the ratio of total leaf area (A) to the whole leaf dry weight (W) over a period of time (30 days interval). The LAR for the healthy and infected plants of both ecotypes was calculated using the equation given by Atwell et al. (1999):
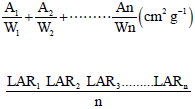
Statistical analysis: Data obtained was analyzed using the independent t-test. Results were also expressed as percentage difference and differences between mean values were determined at 5% probability.
RESULTS
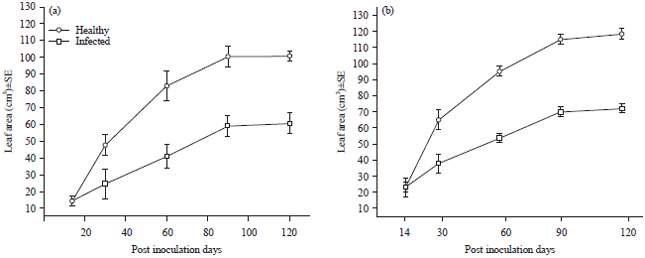
Symptomatology: Evidence of TeMV infection on both ecotypes began to manifest between 10 and 12 DPI. Symptoms induced in ecotype A were green vein-banding, severe mosaic, leaf malformation/leaf distortion and reduced leaf size compared to healthy ones. Ecotype A appeared to be more susceptible comparatively to B. Leaves of the former were smaller than those of the latter. The virus also reduced the quality of leaves. The reduction in leaf size became more pronounced with increasing length of infection (Fig. 1).
Telfairia mosaic virus infection caused significant (p<0.05) decreases in leaf area, shoot height, leaf number, leaf fresh and dry weight, fresh and dry weight of shoot and root of the two ecotypes of T. occidentalis in inoculated plants compared to the healthy. The result also showed that TeMV caused greater decreases in T. occidentalis ecotype A than in ecotype B.
 | |
| Fig. 1(a-d): | (a-b) Healthy and infected leaves of ecotype A at two months and (c-d) Healthy and infected leaves of ecotype B at two months |
| Table 2: | Effect of Telfairia mosaic virus infection on fresh weight and dry weight of shoot and root of Telfairia occidentalis ecotypes |
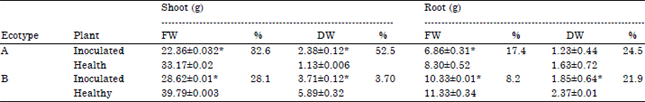 | |
| FW: Fresh weight, DW: Dry weight, Values are Mean±SE, n = 3, p<0.05, *Significant, Percentage difference was obtained by expressing the difference between the value for healthy and inoculated as a percentage of the healthy | |
Changes in mean values in all of these parameters in healthy plants of both ecotypes investigated increased progressively with developmental periods. Results revealed that there was not significant (p<0.05) decrease in the parameters studied at early period of growth (14 DPI) in healthy and inoculated plants of both ecotypes. Reductions became more pronounced with increased length of infection (Fig. 2-6 and Table 2), respectively.
Percentage difference in shoot height for ecotype A and B ranged from 0.07% (14 DPI) to 40.1% (60 DPI) and 0.03% (14 DPI) to 37.4% (60 DPI). Ecotype B had mean values of 30.94±0.00 (14 DPI) to 58.80±0.06 and 30.95±0.00 to 80.57±0.088 (120 DPI) (Fig. 3).
The negative effect of TeMV infection on the number of leaves produced by T. occidentalis was more in ecotype A with percentage difference of 46.6% as against 30.8% for ecotype B. The mean values for number of leaves produced by inoculated plants of ecotype A ranged from 26.03±0.033 to 55.53±0.033 as against values of 26.03 to 66.60±0.058.
 | |
| Fig. 2(a-b): | Effect of Telfairia mosaic virus on leaf area in Telfairia occidentalis ecotypes, (a) Ecotype A and (b) Ecotype B, Mean±SE, n = 5, p<0.05 |
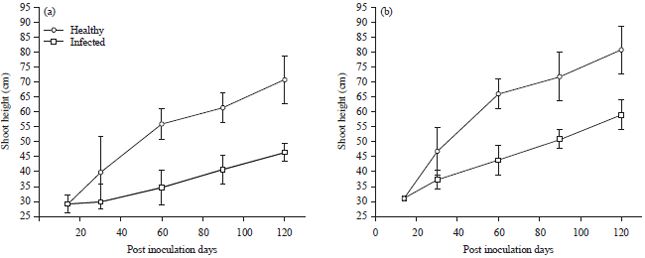 | |
| Fig. 3(a-b): | Effect of Telfairia mosaic virus infection on shoot height in ecotypes of Telfairia occidentalis, (a) Ecotype A and (b) Ecotype B, Mean±SE, n = 5, p<0.05 |
Corresponding mean values for inoculated and healthy plants of ecotype B ranged from 28.17±0.033 to 62.37±0.033 and 28.20±0.00 to 70.70±0.115 (Fig. 4).
Percentage difference values for leaf fresh weight for inoculated and healthy plants of ecotype A and B ranged from 1.43-31.74 and 0.37-23.30%, respectively (Fig. 5). While, corresponding values for leaf dry weight ranged from 10.0% (30 DPI) to 25.0% (90 DPI) and 13.7% (60 DPI) to 20.0% (120 DPI), respectively (Fig. 6).
Percentage difference values for shoot fresh, dry weight and root fresh, dry weight of ecotype A were 32.6, 52.5, 17.4 and 24.5%, respectively. Ecotype B had values of 28.1, 37.0, 8.2 and 21.9%, respectively (Table 2).
The results of RGR and NAR in inoculated and healthy plant samples of T. occidentalis from the two localities revealed a general decrease in values with increasing periods of development with highest and lowest values recorded at 30 and 120 DPI, respectively.
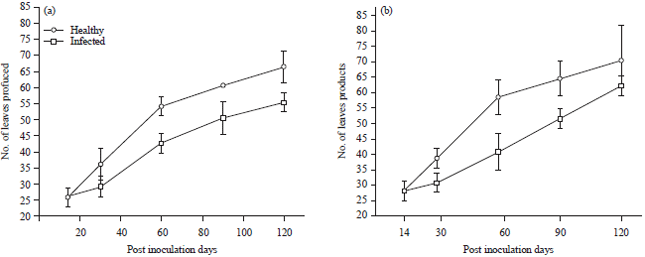 | |
| Fig. 4(a-b): | Effect of Telfairia mosaic virus infection on number of leaves produced by ecotypes of Telfairia occidentalis, (a) Ecotype A and (b) Ecotype B, Mean±SE, n = 5, p<0.05 |
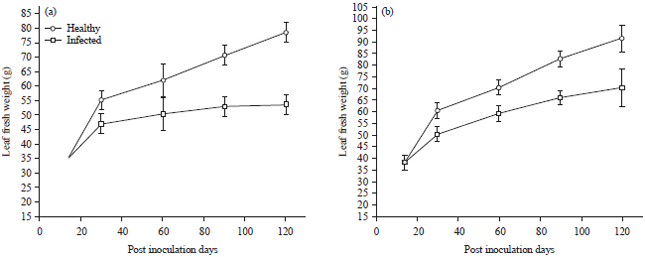 | |
| Fig. 5(a-b): | Effect of Telfairia mosaic virus infection on leaf fresh weight of Telfairia occidentalis ecotypes, (a) Ecotype A and (b) Ecotype B, Mean±SE, n = 5, p<0.05 |
Highest and lowest values percentage difference values for NAR for ecotype A and B were 39.3, 22.1, 34.4 and 20.5%, respectively (Fig. 7). Percentage difference values recorded for RGR for ecotype A and B were 31.7% (30 PDI), 21.1% (120 PDI) and 28.2% (30PDI), 17.5% (120 PDI) (Fig. 8).
Results obtained for LAR for inoculated and healthy plants showed a general trend of increase with progressive periods of growth. Ecotype A had highest and lowest percentage deference values of 26.0 and 13.9% while, ecotype B had values of 25.3 and 15.0% (Fig. 9). Inoculated plants of both ecotypes revealed lower values in RGR, NAR and LAR when compared with healthy plants samples.
Effect of TeMV on RGR, NAR and LAR was more pronounced in T. occidentalis ecotype A than B as revealed by lower values in inoculated plant samples compared with the healthy ones.
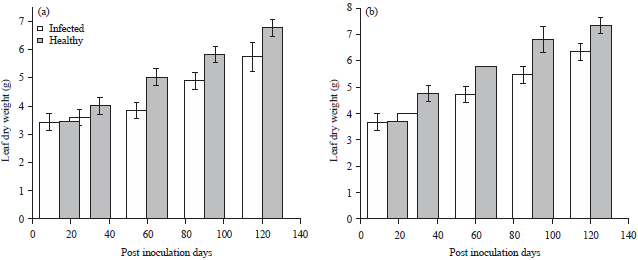 | |
| Fig. 6(a-b): | Effect of Telfairia mosaic virus on leaf dry weight of Telfairia occidentalis ecotypes, (a) Ecotype A and (b) Ecotype B, Mean±SE, n = 5, p<0.05 |
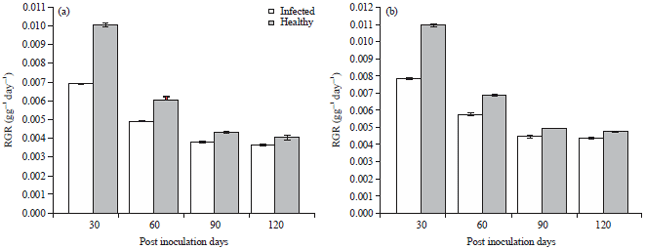 | |
| Fig. 7(a-b): | Effect of Telfairia mosaic virus on the relative growth rate in ecotypes of Telfairia occidentalis, (a) Ecotype A and (b) Ecotype B, Mean±SE, n = 5, p<0.05 |
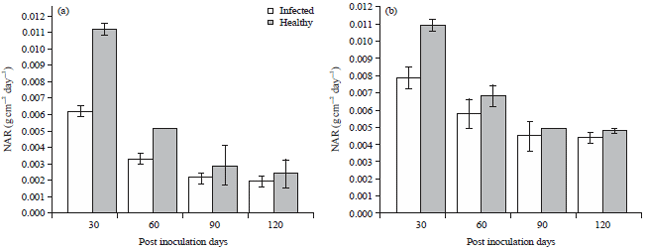 | |
| Fig. 8(a-b): | Effect of Telfairia mosaic virus on net assimilation rate in ecotypes of Telfairia occidentalis, (a) Ecotype A and (b) Ecotype B, Mean±SE, n = 5, p<0.05 |
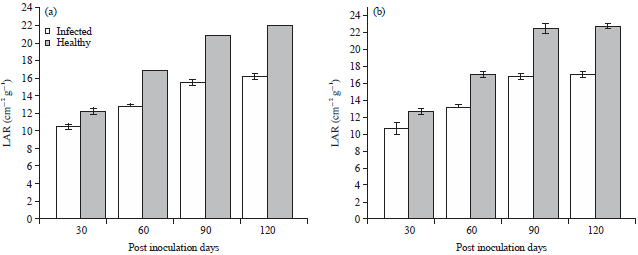 | |
| Fig. 9(a-b): | Effect of Telfairia mosaic virus on leaf area ratio in ecotypes of Telfairia occidentalis, (a) Ecotype A and (b) Ecotype B, Mean±SE, n = 5, p<0.05 |
DISCUSSION
Tecsi et al. (1996) reported that in a compatible interaction between a virus and its host, there must be substantial diversion of metabolites into the accumulation of virus-specific proteins and nucleic acids. This interaction is not accompanied by cell death, in most susceptible plants and that infected cells continue to function despite the presence of massive quantities of virus-specific products. This diversion of resources is evident in the whole plant as changes in plant physiology and development of symptoms occurs (Tecsi et al., 1996). The plant cells are able to accommodate virus synthesis while still sustaining some capacity in the plant for further growth.
Attack by pathogens alters plant primary metabolism which affect growth and development and secondary metabolism which results in the induction of defence programmes (Wood, 1990; Hull, 2002; Heil and Bostock, 2002; Swarbrick et al., 2006). The effect of TeMV on the growth characteristics of two ecotypes of T. occidentalis were studied. The results of infection of TeMV on growth showed significant (p<0.05) reductions in all the growth parameters studied with ecotype A being more susceptible to the effect of the virus than ecotype B. No significant morphological change was observed at early period of infection on all the parameters investigated in spite of severe physiological alterations. This could be due to the fact that it takes between 10-12 days for physical manifestation of TeMV infection on T. occidentalis.
Effect of TeMV on leaf area exhibited no significant change in leaf area at early period of growth but as infection progressed, the leaf area of infected plants became reduced significantly in both ecotypes studied. Similar observations were made by Pawar et al. (1990) and El-Dougdoug et al. (2007), who reported that infection with SRSV on sorghum and PVY on D. metel, respectively significantly reduced the leaf area of infected plants. From the results of this study, infection of TeMV on leaf area was more severe in T. occidentalis ecotype A compared to ecotype B.
Infection of TeMV on shoot height, number of leaves produced, leaf fresh and dry weight, fresh and dry weight of shoot and root exhibited significant (p<0.05) decreases at all developmental stages studied with higher percentage decreases recorded for T. occidentalis ecotype A than B. Similar observations of virus infection on shoot height have been made by Pawar et al. (1990), El-Dougdoug et al. (2007) and Ehinmore and Kareem (2010) on sorghum infected with SRSV, PVY on D. metel and AMV on amaranthus, leaf number (Takahashi, 1972; Fraser and Matthews, 1981; El-Dougdoug et al., 2007; Ehinmore and Kareem, 2010) on tobacco infected with TMV, TYMV, TuMV and CaMV on Chinese cabbage, PVY on D. metel and AMV on amaranthus respectively, leaf fresh and dry weight (El-Dougdoug et al., 2007) on Datura metel infected with PVY.
Because of the fact that total leaf area and number of leaves produced are directly linked with photosynthetic activities which provide the required energy for plant growth and defense against pests and diseases, thus, reduction in these growth parameters caused overall decline in growth resulting in stunted plants, thinner stems and reduction in biomass production when compared with the healthy ones. Hull (2002) submitted that plants become stunted on a dry weight basis if the availability of carbon fixed in photosynthesis is limiting which could be brought about in several ways: by direct effects on the photosynthetic apparatus, starch accumulation in chloroplast, stomatal opening, translocation of fixed carbon and leaf posture.
There were significant (p<0.05) increases in RGR and NAR at early stages of growth and a decrease with increasing period of growth in healthy samples. While, values for LAR decreased at early stages and increased with leaf maturation in healthy samples. A similar trend was recorded for infected samples with significant reductions compared with healthy samples of ecotypes of T. occidentalis investigated. Atwell et al. (1999) observed that variation in LAR frequently has more direct impact on whole-plant growth than variation in NAR. They documented that leaf area is a driving variable for whole plant growth and the proportion of plant biomass invested in leaf area or "leafiness" will have an important bearing on RGR (Atwell et al., 1999) and their reductions caused by the virus will hamper plant growth and development and corresponding yield. Because of the reductions in growth parameters obtained in this study, there is therefore the need to intensify biotechnological research on the production of virus-resistant T. occidentalis.
REFERENCES
- Akubugwo, I.E., N.A. Obasi, G.C. Chinyere and A.E. Ugbogu, 2008. Mineral and phytochemical contents in leaves of Amaranthus hybridus L. and Solanum nigrum L. subjected to different processing methods. Afr. J. Biochem. Res., 2: 40-44.
Direct Link - Afreen, B., M. Gulfishan, G. Baghel, M. Fatma, A.A. Khan and Q.A. Naqvi, 2011. Molecular detection of a virus infecting carrot and its effect on some cytological and physiological parameters. Afr. J. Plant Sci., 5: 407-411.
Direct Link - Booker, H.M., P. Umaharan and C.R. McDavid, 2005. Effect of Cowpea severe mosaic virus on crop growth characteristics and yield of cowpea. Plant Dis., 89: 515-520.
Direct Link - Ehinmore, I. and K.T. Kareem, 2010. Effect of Amaranthus mosaic virus on the growth characters of Amaranthus hybridus. Agric. Biol. J. North Am., 1: 75-79.
Direct Link - El-Dougdoug, K.A., H. Mohamed and A. Abo-Senna, 2007. Effect of PVY viral infection on alkaloid contents of cultivated medicinal plants. J. Applied Sci. Res., 3: 558-563.
Direct Link - Fraser, L. and R.E.F. Matthews, 1981. A rapid transient inhibition of leaf initiation induced by turnip yellow mosaic virus infection. Physiol. Plant Pathol., 19: 325-336.
CrossRefDirect Link - Funayama, S., I. Terashima and T. Yahara, 2001. Effects of virus infection and light environment on population dynamics of Eupatorium makinoi (Asteraceae). Am. J. Bot., 88: 616-622.
Direct Link - Funayama, S., K. Hikosaka and T. Yahara, 1997. Effects of virus infection and growth irradiance on fitness components and photosynthetic properties of Eupatorium makinoi (Compositae). Am. J. Bot., 84: 823-823.
Direct Link - Heil, M. and R.M. Bostock, 2002. Induced systemic resistance (ISR) against pathogens in the context of induced plant defences. Ann. Bot., 89: 503-512.
CrossRefDirect Link - Hooks, C.R.R., M.G. Wright, D.S. Kabasawa, R. Manandhar and R.P.P. Almeida, 2008. Effect of Banana bunchy top virus infection on morphology and growth characteristics of banana. Ann. Applied Biol., 153: 1-9.
CrossRefDirect Link - Jones, M.B., A.J. Heard, J. Woledge, E.L. Leafe and R.T. Plumb, 1977. The effect of ryegrass mosaic virus on carbon assimilation and growth of ryegrasses. Ann. Applied Biol., 87: 393-405.
CrossRefDirect Link - Kazinczi, G., D. Lukacs, A. Takacs, J. Horvath, R. Gaborjanyi, M. Nadasy and E. Nadasy, 2006. Biological decline of Solanum nigrum due to virus infections. J. Plant Dis. Protect., 20: 325-330.
Direct Link - Kareem, K.T., I. Ehinmore, K.E. Oke and O. Arogundade, 2011. The reaction of Amaranthus hybridus to infection by Amaranthus mosaic virus. Int. J. Biol. Chem. Sci., 5: 815-823.
CrossRefDirect Link - Miyoshi, K., T. Sato and N. Takahashi, 1996. Differences in the effects of dehusking during formation of seeds on the germination of seeds of Indica and Japonica rice (Oryza sativa L.). Ann. Bot., 77: 599-604.
CrossRefDirect Link - Rahman, M.S. and A.M. Akanda, 2010. Effect of PLRV infected seed tuber on disease incidence, plant growth and yield parameters of potato. Bangladesh J. Agric. Res., 35: 359-366.
CrossRefDirect Link - Pawar, P.S., T.B. Garud, V.R. Mali and S.D. Choudhari, 1990. Effect of Sorghum Red Stripe Virus (SRSV) on leaf chlorophyll and sugar content of stalk juice in different genotypes of sorghum. Indian Phytopathol., 43: 345-348.
Direct Link - Pazarlar, S., M. Gumus and G.B. Oztekin, 2013. The effects of tobacco mosaic virus infection on growth and physiological parameters in some pepper varieties (Capsicum annuum L.). Not. Bot. Horti Agrobo., 41: 427-433.
Direct Link - Swarbrick, P.J., P. Schulze-Lefert and J.D. Scholes, 2006. Metabolic consequences of susceptibility and resistance (race-specific and broad-spectrum) in barley leaves challenged with powdery mildew. Plant Cell Environ., 29: 1061-1076.
CrossRefDirect Link - Tecsi, L., A.M. Smith, A.J. Maule and C.R. Leegood, 1996. A spatial analysis of physiological changes associated with infection of cotyledons of marrow plants with cucumber mosaic virus. Plant Physiol., 111: 975-985.
PubMedDirect Link