
Walid H. Kilany
National Laboratory for Veterinary Quality Control on Poultry Production (NLQP), Animal Health Research Institute, P.O. Box 264, Giza, Egypt
Mohamed A. Soliman
National Laboratory for Veterinary Quality Control on Poultry Production (NLQP), Animal Health Research Institute, P.O. Box 264, Giza, Egypt
Marwa A. Safwat
National Laboratory for Veterinary Quality Control on Poultry Production (NLQP), Animal Health Research Institute, P.O. Box 264, Giza, Egypt
Osama Mehana
National Laboratory for Veterinary Quality Control on Poultry Production (NLQP), Animal Health Research Institute, P.O. Box 264, Giza, Egypt
Marwa A. El- Magid
National Laboratory for Veterinary Quality Control on Poultry Production (NLQP), Animal Health Research Institute, P.O. Box 264, Giza, Egypt
Mohamed K. Hassan
National Laboratory for Veterinary Quality Control on Poultry Production (NLQP), Animal Health Research Institute, P.O. Box 264, Giza, Egypt
Soad A. Nasif
National Laboratory for Veterinary Quality Control on Poultry Production (NLQP), Animal Health Research Institute, P.O. Box 264, Giza, Egypt
International Journal of Virology
Year: 2015 | Volume: 11 | Issue: 3 | Page No.: 139-145







ABSTRACT
Avian Leukosis Virus subgroup J (ALV-J) is widely described in meat-type chickens and layers type but rarely observed in ducks. In this study, two flocks of Peking duck breeder bred in Egypt showed 25-30% mortality, 20-30% drop in egg production and 60-65% drop in hatchability. Gross picture showed severe enlargement of liver, spleen, white raised nodules in heart and ovarian atrophy in all examined birds. The liver and spleen had diffuse, multifocal white raised foci on the surface as well as on the cut-surface. Histopathological examination revealed numerous myelocytes with bigger volume, large peripheral nucleus and packed reddish cytoplasmic granules infiltrated in heart, liver, kidney and ovary. Some of myelocytic cells had mitotic figures. Results were positive for detection of ALV antigen- p27 by antigen capture ELISA in cloacal samples. The PCR results confirmed that the flocks were positive for ALV-J with specific fragment of 545 bp, but negative for ALV-A, Marek’s Disease Virus (MDV) and Reticuloendotheliosis virus (REV). The study provided some information on ALV-J induced myelocytomatosis for ducks. It concluded that ALV-J virus is broadening host range including the ducks. Also, myeloid leukosis is an enduring problem facing the poultry industry.
PDF Abstract XML References Citation
Received: June 27, 2015;
Accepted: August 31, 2015;
Published: October 14, 2015
How to cite this article
Walid H. Kilany, Mohamed A. Soliman, Marwa A. Safwat, Osama Mehana, Marwa A. El- Magid, Mohamed K. Hassan and Soad A. Nasif, 2015. Detection of Avian Leukosis Virus Subgroup J from Commercial Peking Duck Breeder Farm in Egypt. International Journal of Virology, 11: 139-145.
DOI: 10.3923/ijv.2015.139.145
URL: https://scialert.net/abstract/?doi=ijv.2015.139.145
DOI: 10.3923/ijv.2015.139.145
URL: https://scialert.net/abstract/?doi=ijv.2015.139.145
INTRODUCTION
Avian leukosis virus subgroup J (ALV-J) is one of ten subgroups, six subgroups from A-J was isolated from chickens and turkey and four uncommon subgroups (F, G, H and I) of ALVs were isolated from wild fowl (e.g., pheasant and quail) according to host range, viral envelope and cross-neutralization patterns (Payne, 1992).
The ALV-J (HPRS-103 strain) was first isolated in 1988 as new subgroup of ALV with new env gene designated subgroup-J from white meat-type chickens in Great Britain (Payne et al., 1991, 1992a, b; Bai et al., 1995). For two decades was assumed that ALV-J was restricted in meat type chickens. Although egg-type chickens have been experimentally infected with ALV-J to induce tumors (Payne et al., 1992a) the first field case of ALV-J infection in egg type chickens was reported in China 2004 (Gingerich et al., 2002; Xu et al., 2004). The ALV-J virus was isolated in meat type chickens in Egypt in 2000 (Aly, 2000). During 2000 till 2004 several cases of infected ALV-J in meat type breeder chickens in Egypt (Soliman, 2005; Arafa et al., 2007). The clinical infected hosts of ALV-J are characterized as high mortality, delayed growth and development of a variety of tumors including myelocytomas, hemangiomas, nephromas and erythroblastosis (Arshad et al., 1997). An increase in incidence of myelocytomatosis was reported in meat type chickens with the strain HPRS-103 of ALV-J (Payne et al., 1992b; Fadly and Smith, 1999).
As the spread of the disease is associated with diverse pathotypes and could result in enormous economic losses, ALV-J-induced disease in layer-type chickens has become one of the most important problems facing the global poultry industry (Xu et al., 2004).
Since 2004, the host range of ALV-J has obviously increased, starting from infecting meat-type chickens, layer flocks and then duck flocks as recorded in China (Zeng et al., 2014).
As far we know, this study represent the first report of detection ALV-J in ducks in Egypt depend on histopathological examination, p27 group specific antigen detection of ALV by antigen capture ELISA and molecular detection the env gene of ALV-J in samples from two flocks of commercial Peking ducks.
MATERIALS AND METHODS
Clinical samples: Forty ducks from two breeder flocks (20 per flock) were collected for necropsy and diagnosis. Age of ducks was 35-45 weeks. Cloacal swabs were collected for detection of ALV group specific antigen p27 using Antigen Capture (AC) ELISA. Organs from examined birds that showed gross tumour lesions were used for PCR and histopathological examination.
Histopathological examination: Samples of liver, spleen, ovary and heart were collected and fixed in 10% buffered neutral formalin. The fixed tissues were embedded in paraffin, sectioned at 4 μm thick and stained with haematoxylin and eosin. The slides were examined under light microscopy.
ALV-p27 antigen capture ELISA: Group specific antigen (gsA) for Avian leukosis virus p27 antigen was tested as described by Fadly and Witter (1998). Cloacal swabs of 20 ducks/flock collected in tubes containing sterile PBS (pH 7.4), with antibiotics. Solid materials were removed from samples by centrifugation at 2000 rpm for 10 min. Supernatants were taken and drawn off in sterile tube, tested for detection of ALV-p27 antigen by using an Antigen Capture Enzyme-Linked Immunosorbent Assay (AC-ELISA) Commercial ELISA kits (IDEXX laboratories, Inc, Main).
Polymerase Chain Reaction (PCR): Organs obtained from birds of positive ALV-p27 antigen were selected for further confirmation using PCR. Proviral DNA was extracted from organs using Bio-robot EZ1automated extractor and EZ1 virus mini kit cat. No. 955134 (Qiagen GmbH, Germany).
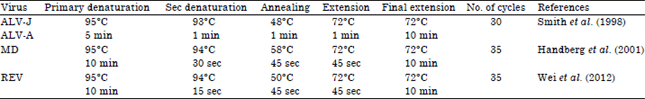
Primers and PCR amplification: Primers were supplied from Metabion (Germany) and were listed in Table 1. The ALV-J and ALV-A (Smith et al., 1998), MDV (Handberg et al., 2001) and REV (Wei et al., 2012), were performed, respectively. Primers were utilized in a 25 μL reaction containing 12.5 μL of Emerald Amp Max PCR Master Mix (Takara, Japan), 1 μL of each primer of 20 pmol concentrations, 4.5 μL of water and 6 μL of DNA template. The reaction was performed in a T3 Biometra thermal cycler. Thermal programs were listed in Table 2.
| Table 1: | List of forward and reverse primers for ALV-J, ALV-A, MDV and REV |
 | |
| Table 2: | Thermal conditions for ALV-J, ALV-A, MDV and REV using applied thermal cycler |
 | |
Analysis of PCR: The products of PCR were separated by electrophoresis on 1.5% agarose gel (Applichem, Germany, GmbH) in 1X TBE buffer at room temperature using gradients of 5 V cm–1. For gel analysis, 15 μL of the products was loaded in each gel slot. A 100 bp DNA Ladder (Qiagen, Germany, GmbH) was used to determine the fragment sizes. The gel was photographed by a gel documentation system (Alpha Innotech, Biometra) and the data was analyzed through computer software.
Primer H5 was designed against the 3' region of the pol gene and was conserved across several ALV subgroups. Primer H7 was designed from a well conserved region of the gp85 sequence of the variant viruses and is expected to give a 545 bp product in amplifications carried out with primer H5 (Smith et al., 1998).
RESULTS
Gross lesions: Examined birds showed depression, recumbence and lethargy. Grossly, birds were characterized with enlarged visceral organs. Liver was friable in consistency and showed diffuse multifocal, white foci (Fig. 1a). Raised multifocal white foci were observed on surface of the heart (Fig. 1b). Ovarian atrophy was prominently detected in all examined birds.
Histopathology: Microscopic appearance was characterized by aggregates of well differentiated myelocytes. Their nuclei were large and peripherally located. The cytoplasm was closely crowded with esinophilic granules, which are usually spherical. Areas of less differentiated myelocytes can be seen (Fig. 2d). In the liver, these groups of myelocytes were seen surrounding blood vessels and made pressure atrophy on hepatocytes. As a result areas of necrosis and few round mononuclear cells can be seen in liver. The Disse’s spaces of liver were dilated and engorged with blood (Fig. 2a). In the ovary, myelocytes accumulated in the stroma layer underneath the granulosa cells (Fig. 2b). In the heart, multifocal aggregates of myelocytes were observed in the myocardium and covered by the pericardium (Fig. 2c).
AC-ELISA: The presence of ALV group specific antigen (gsA) p27 in the samples was demonstrated positive results by AC-ELISA on anti-p27 antibody-coated plates.
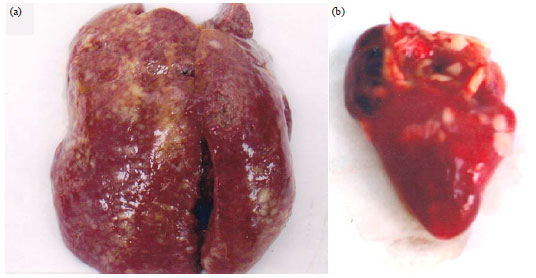 | |
| Fig. 1(a-b): | Gross lesion of examined birds, (a) Enlarged liver, with diffuse multifocal white flattened foci throughout enlarged lobes and (b) Multifocal white raised foci over the heart surface |
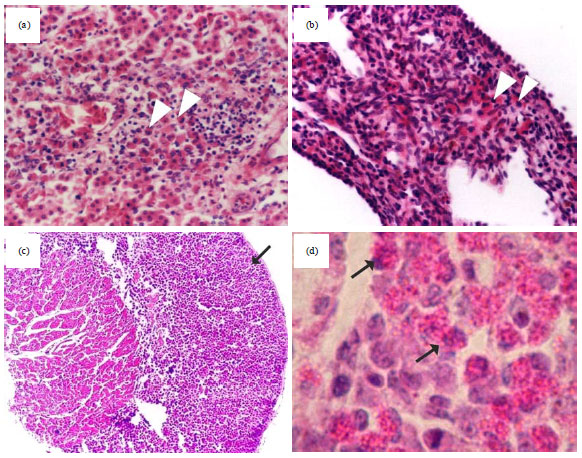 | |
| Fig. 2(a-d): | Histopathology, (a) Myelocytes were accumulated in dilated Disse’s space of liver, X100, (b) Myelocytes were accumulated in stroma of granulosa cells in ovary, X100, (c) Focal aggregates of myelocytes was raised on myocardium of the heart, X40 and (d) High magnification of myelocytic cell with bright eosinophilic granules and peripheral nucleus, X630 H and E |
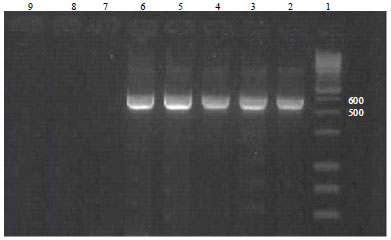 | |
| Fig. 3: | Result of PCR for ALV-J detection, Lane1: 1 kbp marker, Lane2-6: Five samples from duck samples showed positive amplification at 545 bp molecular weights, Lane 7: ALV-A primers were used, Lane 8: MDV primes were used and Lane 9: REV primers were used |
PCR assay: Organs obtained from birds of positive ALV-p27 antigen were selected for further confirmation using PCR. Provirus DNA was extracted from liver and ovary of the ducks from the two flocks. Pooled extracts (n = 29) were assayed using ALV-J, ALV-A, REV and MDV specific primers. The results showed that all the pooled samples tested were negative for ALV-A, REV and MDV. Samples were positive for ALV-J with a PCR product fragment of 545 bp as expected (Fig. 3).
DISCUSSION
The present study demonstrated the occurrence of ALV-J in commercial Peking duck breeder. Myelocytomatosis induced by ALV-J was characterized by numerous myelocytes aggregated in the liver, heart and ovary of the infected birds. The ALV-J mainly induce myelocytomatosis because of their tropism to the cells of the myeloid rather than the lymphoid lineage (Payne, 1998; Payne and Fadly, 2003).
Compression of proliferative myelocytes in the liver and the ovary triggered hepatic necrosis and ovarian atrophy. The observed visceral myelocytomatosis in the examined flocks was the prominent cause for the mortalities and the decrease in egg production. The ALV-J since reported, cause a major economic problem in chickens and other domestic fowls worldwide associated with the high oncogenicity and broad tumor spectrum (Venugopal, 1999). Other neoplastic viral diseases, MDV and REV were eliminated via histopathological examination due to absence of malignant lymphocytes.
Commercial antigen capture ELISA kit for demonstration of ALV gsA p27 in the cloacal swabs was used to detect the shedder birds which transmit ALV horizontally (Koch et al., 2000). Meconium for chicks and fecal matter contain high concentrations of ALV and are an important source of virus for horizontal transmission to the housemates. The horizontal transmission that occurs through close contact with infected birds is a major challenge for the eradication programs since the spread of infection can rise quickly during the first few weeks of infection (Venugopal, 1999). Positive results of AC-ELISA for detection of ALV gsA p27 in cloacal swabs indicate presence of positive shedder ducks. The p27 positive results were further supported using the reference primers for molecular detection of ALV-J. The PCR reactions for the positive (gsA) p27 ducks were to detect the specific env gene for ALV-J (Zeng et al., 2014).
In the present study, the env gene of extracted DNA from pooled organs were detected as the env genes in prototype strain HPRS-103 as in broiler ALV-J isolates and they were similarly detected with some layer chicken ALV-J isolates (Xu et al., 2004).
The ALV-J have changed rapidly, which results in a change in host range, first infecting meat-type chickens and layer flocks, then detected in wild ducks in china. Molecular characterization of ALV-J in wild ducks in China identified amino acids substitutions when compared with the prototype strain HPRS-103. These substitutions might be related with changes in the host range and pathogenicity of ALV-J (Zeng et al., 2014).
This study demonstrated that ALV-J was detected in domestic ducks. Further studies of molecular epidemiology of ALV-J in ducks required to answer several questions whether the strain found in ducks come from other chicken flocks or from wild ducks.
ACKNOWLEDGMENT
The present study was financially supported by National Lab for Veterinary Quality Control on Poultry Production.
REFERENCES
- Arshad, S.S., K. Howes, G.S. Barron, L.M. Smith, P.H. Russell and L.N. Payne, 1997. Tissue tropism of the HPRS-103 strain of J subgroup avian leukosis virus and of a derivative acutely transforming virus. Vet. Pathol. Online, 34: 127-137.
CrossRefDirect Link - Bai, J., K. Howes, L.N. Payne and M.A. Skinner, 1995. Sequence of host-range determinants in the env gene of a full-length, infectious proviral clone of exogenous avian leukosis virus HPRS-103 confirms that it represents a new subgroup (designated J). J. Gen. Virol., 76: 181-187.
CrossRefDirect Link - Fadly, A.M. and R.L. Witter, 1998. Oncornaviruses: Leukosis/Sarcoma and Reticuloendotheliosis. In: A Laboratory Manual for the Isolation and Identification of Avian Pathogens, Swayne, D.E. (Ed.). 4th Edn., American Association of Avian Pathologists, Kennet Square, PA., ISBN-13: 9780915538072, pp: 185-196.
- Fadly, A.M. and E.J. Smith, 1999. Isolation and some characteristics of a subgroup J-like avian leukosis virus associated with myeloid leukosis in meat-type chickens in the United States. Avian Dis., 43: 391-400.
CrossRefDirect Link - Gingerich, E., R.E. Porter, B. Lupiani and A.M. Fadly, 2002. Diagnosis of myeloid leukosis induced by a recombinant avian leukosis virus in commercial white leghorn egg laying flocks. Avian Dis., 46: 745-748.
Direct Link - Handberg, K.J., O.L. Nielsen and P.H. Jorgensen, 2001. The use of serotype 1- and serotype 3-specific polymerase chain reaction for the detection of Marek's disease virus in chickens. J. Virol., 30: 243-249.
CrossRefDirect Link - Koch, G., J. van der Velde, L. Hartog, A.L.J. Gielkens and W.J.M. Landman, 2000. Horizontal and vertical transmission of ALV-J and ALV-A virus in broiler breeders chickens. Proceedings of the International Symposium on ALV-J and other Avian Retroviruses, Jun 5-8, 2000, Rauischholzhausen, Germany, pp: 141-151.
- Payne, L.N., S.R. Brown, N. Bumstead, K. Howes, J.A. Frazier and M.E. Thouless, 1991. A novel subgroup of exogenous avian leukosis virus in chickens. J. Gen. Virol., 72: 801-807.
PubMedDirect Link - Payne, L.N., K. Howes, A.M. Gillespie and L.M. Smith, 1992. Host range of Rous Sarcoma Virus pseudotype RSV(HPRS-103) in 12 avian species: Support for a new avian retrovirus envelope subgroup, designated J. J. Gen. Virol., 73: 2995-2997.
ASCIDirect Link - Payne, L.N., A.M. Gillespie and K. Howes, 1992. Myeloid leukaemogenicity and transmission of the HPRS-103 strain of avian leukosis virus. Leukemia, 6: 1167-1176.
PubMedDirect Link - Payne, L.N., 1998. HPRS‐103: A retro virus strikes back. The emergence of subgroup J avian leukosis virus. Avian Pathol., 27: S36-S45.
CrossRefDirect Link - Smith, L.M., S.R. Brown, K. Howes, S. McLeod and S.S. Arshad et al., 1998. Development and application of Polymerase Chain Reaction (PCR) tests for the detection of subgroup J avian leukosis virus. Virus Res., 54: 87-98.
CrossRefPubMedDirect Link - Venugopal, K., 1999. Avian leukosis virus subgroup J: A rapidly evolving group of oncogenic retroviruses. Res. Vet. Sci., 67: 113-119.
CrossRefDirect Link - Wei, K., Z. Sun, S. Zhu, W. Guo and P. Sheng et al., 2012. Probable congenital transmission of reticuloendotheliosis virus caused by vaccination with contaminated vaccines. Plos One, Vol. 7.
CrossRefDirect Link - Xu, B., W. Dong, C. Yu, Z. He and Y. Lv et al., 2004. Occurrence of avian leukosis virus subgroup J in commercial layer flocks in China. Avian. Pathol., 33: 13-17.
Direct Link - Zeng, X., L. Liu, R. Hao and C. Han, 2014. Detection and molecular characterization of J subgroup avian leukosis virus in wild ducks in China. Plos One, Vol. 9.
CrossRefDirect Link