
E.K.F. Elbeshehy
Department of Agricultural Botany and Plant Pathology, Faculty of Agriculture, Suez Canal University, Egypt
A.A.A. Sallam
Department of Agricultural Botany and Plant Pathology, Faculty of Agriculture, Suez Canal University, Egypt
International Journal of Virology
Year: 2012 | Volume: 8 | Issue: 1 | Page No.: 90-97







ABSTRACT
Cucumber Mosaic Virus (CMV) was isolated from naturally infected cucumber plants Cucumis sativus L. grown in various garden and greenhouses of Ismailia Governorate, Egypt exhibiting systemic mosaic, blistering, fruit malformation and stunted plant growth and identified by biological, serological and molecular analysis. The isolated virus gave positive reaction with CMV antiserum but not with antibodies of WMV and SqMV using DAS-ELISA. CMV was able to infect different host plant species including squash, pumpkin, pepper, bean, Chenopodium amaranticolor and cowpea, showing foliar symptoms of mosaic, deformations and necrotic and chlorotic ring spots, that resemble those induced by CMV. SDS-PAGE test showed various distinguishable sole novel protein bands in four cucumber cultivars infected with CMV but not in healthy one. RT-PCR, with the primer CMV1 and CMV2 for CMV-cp. gene, yielded 422 base pair DNA fragments. The following sequences were used in the comparison: Brazil (AF418577), China (FJ403473), New Zealand (AY861395) and India (AJ810260). The partial nucleotide sequence alignment, showed (95%) homology between CMV New Zealand isolate and CMV-Egyptian isolate. The sequence alignment also indicated that the CMV isolate is far from similarity with India CMV, showing (92%) according to the phylogenetic homology tree which were 92 to 99% identical to those of CMV-Egypt isolate.
PDF Abstract XML References Citation
Received: June 20, 2011;
Accepted: July 28, 2011;
Published: October 25, 2011
How to cite this article
E.K.F. Elbeshehy and A.A.A. Sallam, 2012. Partial Characterization of an Isolate of Cucumber Mosaic Virus from Ismailia Governorate. International Journal of Virology, 8: 90-97.
DOI: 10.3923/ijv.2012.90.97
URL: https://scialert.net/abstract/?doi=ijv.2012.90.97
DOI: 10.3923/ijv.2012.90.97
URL: https://scialert.net/abstract/?doi=ijv.2012.90.97
INTRODUCTION
About twenty economically important viruses were detected in cucumber (Cucumis sativus L.) crops (Brunt et al., 1996). Cucumber Mosaic Virus (CMV) is reported to infect 1287 plant species. CMV was detected in leguminous, ornamental and tomato plants (Zitikaite and Staniulis, 2006) is one of the most common plant viruses of substantial agricultural importance, infecting more than 1,000 plant species (Van Regenmortel et al., 2000). CMV the type species of the genus Cucumovirus, family Bromoviridae, CMV is a single-stranded positive-sense tripartite genome RNA virus. CMV isolates have been described in several hosts (Maciel-Zambolim et al., 1990, 1994; Dusi et al., 1992; Duarte et al., 1994; Boari et al., 2000; Eiras et al., 2000, 2001; Colariccio et al., 1987, 1996, 2002). CMV are considered to be very important in Africa general and Egypt specially as for their geographical distribution, pathogenic variation and yield loss. The main purpose of this study was isolation and identification of CMV using ELISA, distinguishable sole bands in both healthy and diseased leaves for which it could be used according to it is presence as sign for the virus infection by SDS-PAGE test and RT-PCR technique. The following sequences were used in the comparison: Brazil (AF418577), China (FJ403473), New Zealand (AY861395) and India (AJ810260) (Takeshita et al., 2001; Colariccio et al., 2002; Harakava and Chagas, 2002).
MATERIALS AND METHODS
Serological diagnosis of the virus isolate: Samples of cucumber plants exhibiting systemic mosaic and stunting, reduced fruit yield and deformation were collected from different fields of Ismailia Governorate. DAS-ELISA technique was used for virus detection as described by Clark and Adams (1977) using the antiserum specific for CMV and compared with those of WMV and SqMV. All viruses tested in triplicate using conventional double antibody sandwich enzyme-linked immuno-sorbent assay (DAS-ELISA) according to the manufacture’s instructions (Sanofi-Santi animal, France). Optical density was measured at λ = 405 nm in an ELISA micro well reader (using Dynatech Immunoassay MR 7000). Samples with an absorbency of at least twice that the healthy controls were considered as a positive for the presence of virus.
Virus samples and natural hosts: Plant samples which gave positive reaction in the direct ELISA test with CMV were used as a source of the virus. Inoculum was prepared and used to inoculate the following indicator hosts Chenopodium amaranticolor Coste and Reyn and Vigna unguiculata L. cv. Borma. To obtain virus isolate in a pure form, the single local lesion technique was followed according to Kuhn (1964). Cucumber (Cucumis sativus L.), Squash (Cucurbita pepo L.), Pumpkin (Cucurbita maxima), pepper (Capsicum annuum L.) and Bean (Phaselous vulgaris) known as a systemic hosts for CMV were inoculated with the virus. Inoculated plants were kept in separate cages, as a source of virus infection.
CMV protein marker
Total soluble protein extraction: The leaves of four cucumber cultivars (Dina, Razena, Prince and Riea) were collected from infected and healthy plants and ground to flour in a mortar by using liquid nitrogen. Total soluble proteins were extracted in SDS reducing buffer, (store at room temperature) composed of Deionized water (38 mL), 0.5 M Tris-HCl-pH 6.8 (10 mL), Glycerol (8 mL), 10% (w/v) SDS (16 mL), 2-mercapto-ethanol (4 mL) and 1% (w/v) Bromophenol blue (4 mL) until became total volume 80 mL. The sample was diluted at least 1:4 with sample buffer and the extract was centrifuged at 10.000 rpm for 20 min. Then 10 μL of total soluble protein was taken for electrophoresis.
Protein analyses: Sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) was carried out in 10% acrylamide slab gels following the system of Laemmli (1970).
Separating gels composed of 0.75 M Tris-HCl pH 8.8, 10% SDS, 0.025% of N, N, N, N-Tetramethylenediamine (TEMED) and 30% ammonium persulfate.
Stacking gels contained 0.57 M Tris-HCl pH 6.8, 10% SDS, 0.025% TEMED and 30% ammonium persulfate. Electrode buffer contained 0.025 M Tris, 0.192 M glycine, 0.1% SDS and pH 8.3. Electrophoresis was carried out with a current of 25 mA and 130 volts per gel until the bromophenol blue marker reached the bottom of the gel after 3 h. After electrophoresis, the silver staining method for protein described by Sammons et al. (1981) was used. This method of staining is sensitive and detects as little as 2 mg of protein in single band. Then the gels were photographed. Using SDS-PAGE marker for molecular weight 14.000-29.000-66.000 Cat. Number: WM-SDS-70 L 1 mg mL-1.
Total RNA extraction and RT-PCR: EZ-10 Spin Column Total TNA Minipreps Super Kit (BIO BASIC INC) was used to extract the total RNA from infected and uninfected cucumber leaves according to the kit procedure. A newly designed oligonucleotide specific CMV1 (5´GCCGTAAGCTGGATGGACAA3´). CMV2(5`TATGATAAGAAGCTTGTTTCGCG 3`) primers (Wylie et al., 1993), were used for RT-PCR amplification of CMV coat protein gene. cDNA (One step RT-PCR) was amplified as described by (AB gene, UK). PCR was carried out in the Biometra T Gradient thermocycler with one initial denaturation cycle at 94°C for 2 min, followed by 30 cycles of amplification with temperature profiles of 1 min at 94°C, 1 min at 50°C and 2 min at 72°C. Thirty cycles were followed by one final extension cycle at 72°C for 7 min.
PCR analysis: The amplified DNA was electrophoresed on 1% agarose gel with 1xTAE buffer, stained with ethidium bromide and photographed using (Gel Doc 2000 Bio-RAD). The molecular weight of the PCR products were determined by comparison with DNA markers, 100 bp ladder (AB gene, UK),
Sequencing and phylogenetic analysis: The RT-PCR product amplified with CMV1 and CMV2 primer was used as template using the same primer. The reaction was performed in a final volume of 50 μL consisting of 1x PCR buffer (20 mM Tris-HCl pH 8.4, 50 mM KCl, 2.5 mM MgCl2), 200 μM dNTPs, 1 μL of DNA target from CMV PCR product, 10 μM of each primers., 2.5 units of Taq DNA polymerase (5 Units μL-1, AB gene, UK). The conditions for amplification were a denaturation step at 94°C for 2 min followed by 30 cycles of 1 min at 94°C, 1 min at 50°C and 2 min at 72°C, with a final extension of 10 min at 72°C. Reactions were cycled in a thermocycler Biometra T Gradient. PCR products were purified with GFXTM PCR purification kit (Amersham Pharmacia Biotech Inc. USA). The sequence was carried out in Macrogen Inc, gene link DNA Sequencing service, Seoul, Korea.
The nucleotide sequence of the coat protein was compared and analyzed using DNAMAN Sequence analysis Software (Lynnon BioSoft. Quebe, Canada) comparison with the CMV isolates previously characterized and available in the Gene Bank was achieved. The phylogenetic analysis and the homology rate were also carried out.
RESULTS AND DISCUSSION
Serological diagnosis of the virus isolate: The identity of thevirus isolate was confirmed by DAS-ELISA technique using kits supplied from SANOFI. Positive reaction was obtained with CMV specific antisera.
Virus samples and natural hosts: Infected cucumber plants with CMV showed viral symptoms of systemic mosaic, blistering, fruit malformation and stunted plant growth. All samples gave positive reaction with CMV and were susceptible to cucumber mosaic viral infection with nearly the same degree of sensitivity. CMV was able to infect different host plant species including Squash (Cucurbita pepo L.), Pumpkin (Cucurbita maxima), Pepper (Capsicum annuum L.), Bean (Phaselous vulgarisX), Chenopodium amaranticolor and Cowpea (Vigna unguiculata), showing foliar symptoms of mosaic, deformations and necrotic and chlorotic ring spots after inoculation, that resemble those induced by CMV.
Symptoms started to appear 7-14 days after inoculation. Cucumber plants, showed systemic mosaic after 8 days from inoculation with extracts from the positive reacted samples (Fig. 1). This virus was isolated in previous studies from cowpea and other hosts, by several investigators in different countries (Daniels and Campbell, 1992; Hu et al., 1995; Carrere et al., 1999; Takeshita et al., 2001; Eiras et al., 2000, 2001).
 | |
| Fig. 1: | Symptoms caused by CMV showing a wide range of discoloration and fruit malformation. (a) Systemic mosaic on C. sativus, (b) Necrotic ring spot on Ch. amaranticolor, (c) Chlorotic ring spot on V. unguiculata, (d) Mosaic on C. maxima, (e) Chlorotic blotching on C. pepo, (f, g) Mosaic and Leaf deformation on C. annuum, (h) Mosaic and blisters on Ph. vulgaris and (i, j, k) Mosaic and fruit malformation on pumpkin, squash and cucumber fruits, respectively |
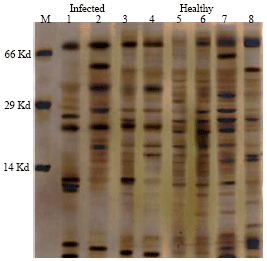 | |
| Fig. 2: | Sodium dodecyl sulphat– polyacrylamide gel electrophoresis (SDS-PAGE) of infected and non infected cucumber leaves cultivars infected with CMV. Lanes 1, 2, 3 and 4 (infected leaves):Lane 1 = cv. Dina, Lane 2 = cv. Razena Lane 3 = cv. Prince, Lane 4 = cv. Riea. Lanes 5, 6, 7 and 8 (non-infected leaves) Lane M = SDS-Marker |
CMV protein marker: Electrophoresed and densitometric analysis of protein bands by SDS-PAGE are illustrated in Fig. 2. The results tabulated in Table 1 showed various distinguishable sole bands in diseased leaves for which it could be used according to it is presence as sign for the CMV infection.
| Table 1: | Hypothesized protein markers for the detection of CMV infection in different cucumber leave cultivar |
 | |
| Bands present in diseased and absent in healthy. M.W: Molecular weight (K.Da), %Amt: Percentage of Amount of protein | |
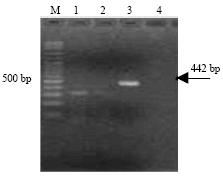 | |
| Fig. 3: | Agarose gel electrophoresis analysis of amplified CMV-cp gene fragment Lanes (1 to 4). RT-PCR products of four CMV samples showing amplified CMV-cp, Gene fragment of the correct size 442 bp (arrow) in lanes (3). Lane (1, 2 and 4) healthy squash. M: 100 bp ladder (AB gene, UK) |
Inference brought about in this table hinted to consideration that it would be acceptable if we assume there is a possible correlation between the virus infection and the novel protein bands presence and showed summary for protein bands markers suggested for detection of CMV infection in different cucumber leaves cultivars (Dina, Razena, Prince and Riea) were presented in disease leaves and absented in healthy. These bands named novel protein bands were distinguished in molecular weight between 70 and 25 Kd in infected four cucumber cultivars. These bands maybe create in infected lanes result effect the virus in plant metabolism and response of plant for CMV infection. Percentage of amount of protein (% Amt.) in Dian and Razena cv., more than % Amt. in Prince and Riea cv., these result illustrated that susceptibility Dian and Razena cv., to CMV infection in excess of Prince and Riea. These results are in agreement with those reported by Sayda (2001). Data in this result confirmed that the protein markers consistently produced in different Cucumber leaves cultivars might suggesting for the detection of CMV infection. In synopsis, during the investigation regarding percentage determined in each band (Sang and Joo, 1992; Sayda, 2001).
Reverse transcription-polymerase chain reaction RT-PCR: The RT-PCR was used for detection of CMV Coat Protein (cp) gene in infected cucumber. PCR fragment of correct size 422 bp was amplified with the primer CMV1 and CMV2 for CMV-cp. gene. Agarose gel electrophoresis analysis of the amplified PCR products is demonstrated in Fig. 3. However, no product was amplified from healthy cumber plants using the same procedure.
Sequence analyses: DNA amplified product from cucumber plants representing Cucumber mosaic virus CMV show 442 bp in Fig. 3. PCR product using CMV1 and CMV2 primer was purified with GFXTM purification kit.
 | |
| Fig. 4: | Sequence alignment of Cucumber mosaic virus CMV coat protein gene sequences. Brazil (AF418577), China (FJ403473), New Zealand (AY861395) and India (AJ810260) |
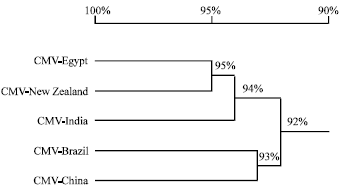 | |
| Fig. 5: | The phylogenetic homology tree based on multiple sequence alignments of the Cucumber mosaic virus CMV coat protein gene with reference to international isolates |
A multiple alignment was done along with sequences previously obtained which have already been used as reference sequences in other studies and from the Gene bank sequence data (Eiras et al., 2004; Deyong et al., 2005). The following sequences were used in the comparison: Brazil (AF418577), China (FJ403473), New Zealand (AY861395) and India (AJ810260). The partial nucleotide sequence alignment in Fig. 4 showed (95%) homology between CMV New Zealand isolate and CMV-Egyptian isolate. The sequence alignment also indicated that the CMV isolate is far from similarity with India CMV, showing (92%) according to the phylogenetic homology tree (Fig. 5).
REFERENCES
- Colariccio, A., C.M. Chagas, J.T. Ferrari, M. Eiras and A.L.R. Chaves, 2002. Molecular characterization and phylogenetic analysis of cucumber mosaic virus in zingiber officinale in Brazil. Proceedings of the 12th International Congress of Virology, (CV, 2002), The World of Microbes, Paris, France,-pp:443.
CrossRef - Daniels, J. and R.N. Campbell, 1992. Characterization of Cucumber mosaic virus isolates from California. Plant Dis., 76: 1245-1250.
CrossRefDirect Link - Deyong, Z., P. Willingmann, C. Heinze, G. Adam, M. Pfunder, B. Frey and J.E. Frey, 2005. Differentiation of cucumber mosaic virus isolates by hybridization to oligonucleotides in a microarray format. J. Virol. Methods, 123: 101-108.
CrossRef - Duarte, L.M.L., E.B. Rivas, M.A.V. Alexandre and J.T. Ferrari, 1994. Detection of CMV isolates from commelinaceae species. Fitopatologia Bras., 19: 248-253.
Direct Link - Hu, J.S., H.P. Li, K. Barry and M. Wang, 1995. Comparison of dot blot, ELISA and RT-PCR assays for detection of two cucumber mosaic virus isolates infecting banana in Hawaii. Plant Dis., 79: 902-906.
Direct Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Sang, G.K. and J.Y. Joo, 1992. Changes in protein patterns resulting from infection of rice leaves with Xanthomonas oryzae pv. orayzae. Mol. Plant-Microbe Interact., 5: 356-360.
Direct Link - Wylie, S., C.R. Wilson, R.A.C. Jones and M.G.K. Jones, 1993. A polymerase chain reaction assay for cucumber mosaic virus in lupin seeds. Aust. J. Agric. Res., 44: 41-51.
Direct Link - Sammons, D.W., L.D. Adams and E.E. Nishizawa, 1981. Ultrasensitive silver-based color staining of polypeptides in polyacrylamide gels. Electrophoresis, 2: 135-141.
CrossRefDirect Link - Clark, M.F. and A.N. Adams, 1977. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol., 34: 475-483.
CrossRefPubMedDirect Link