
Ahmed M. Soliman
Virus and Phytoplasma Research Section, Plant Pathology Research Institute, Agricultural Research Center (ARC), Giza, Egypt
Sabry Y.M. Mahmoud
Department of Agricultural Botany (Agricultural Microbiology), Faculty of Agriculture, Sohag University, Sohag, Egypt
Rehab A. Dawood
Virus and Phytoplasma Research Section, Plant Pathology Research Institute, Agricultural Research Center (ARC), Giza, Egypt
International Journal of Virology
Year: 2012 | Volume: 8 | Issue: 1 | Page No.: 61-70







ABSTRACT
Garlic samples showing symptoms of Onion Yellow Dwarf Virus (OYDV) were obtained from previous study and tested by indirect-enzyme linked immunosorbent assay (I-ELISA), transmitted to Chenopodium amaranticolor and then confirmed by immunosorbent electron microscopy (ISEM) for the presence of OYDV. PCR primers were used to amplify about 1.1 kb fragment from the viral genome using RT-PCR from infected garlic plants, such fragment were not obtained from healthy- looking plants and/or virus-free seedlings of shoot-tips. The amplified products of OYDV was cloned into pGEM®-T Easy vector and transformed into Escherichia coli strain DH5a. The recombinant plasmids were obtained and sequenced. The nucleotide sequences were compared with corresponding viral nucleotide sequences reported in GenBank. The sequence analysis showed that; nucleotide sequence of OYDV-EG [Egyptian isolate (HM473189)] had 82-96% similarity compared with ten reported OYDV isolates. Thus, a method of identification and detection by RT-PCR of OYDV was established. This study also aimed to obtain OYDV-free plants from infected garlic plants using cloves subjected to electrotherapy, thermotherapy, chemotherapy or meristematic dissection followed by in vitro culture. A combination treatment with electro- and chemotherapy (15 mA/10 min + 20 mg L-1 virazol) was found to more effective on viral elimination and survival of explants. ELISA tests showed that 85% of the plantlets that survived were OYDV-negative.
PDF Abstract XML References Citation
Received: June 20, 2011;
Accepted: July 27, 2011;
Published: October 25, 2011
How to cite this article
Ahmed M. Soliman, Sabry Y.M. Mahmoud and Rehab A. Dawood, 2012. Molecular Characterization of Onion Yellow Dwarf Virus (Garlic Isolate) with Production of Virus-free Plantlets. International Journal of Virology, 8: 61-70.
DOI: 10.3923/ijv.2012.61.70
URL: https://scialert.net/abstract/?doi=ijv.2012.61.70
DOI: 10.3923/ijv.2012.61.70
URL: https://scialert.net/abstract/?doi=ijv.2012.61.70
INTRODUCTION
Many viruses are known to infect garlic (Allium sativum) and a few of them can seriously reduce crop yields and quality. Losses of around 25-50% due to natural infections have been reported (Lot et al., 1998; Dovas et al., 2001a). Whatever their primary means of transmission are based on vegetative propagation of the crop favors their dissemination and accumulation in bulbs (AVRDC, 1997). Onion Yellow Dwarf Virus (OYDV), Leek Yellow Stripe Virus (LYSV) (genus, Potyvirus , family Potyviridae), Garlic common latent virus (GarCLV), Garlic latent virus (GarLV) and Shallot Latent Virus (SLV) (genus, Carlavirus) have been reported to infect garlic, often as mixed infections (Dovas et al., 2001b; Chen et al., 2004; Meenakshi et al., 2006; Shahraeen et al., 2008). OYDV, an aphid-borne potyvirus, is one of the major viral pathogens of onion and garlic. In garlic, OYDV produces symptoms of mild chlorotic stripes to bright yellow stripes depending on virus isolate and cultivars. Reduction in growth and bulb size also occurs (FAO/IPGRI, 1997). OYDV is recognized as a major element of the virus disease complex in garlic (Takaichi et al., 2001).
In previous study, OYDV was isolated and detected in garlic leaves by ELISA and immunocapture/reverse. Transcription-polymerase chain reaction (IC- PCR) (Mahmoud et al., 2007). Since ELISA can not be routinely used for detection of OYDV, a RT- PCR based method was standardized for detection of OYDV in leaves and bulbs of garlic. Group-specific primers used for detection of potyviruses for the amplification of 3' terminal region and part of NIB gene (Pappu et al., 1993; Gibbs and Mackenzie, 1997) did not work, therefore, specific primers from conserved region of RNA-dependent RNA polymerase gene and 3' UTR region of viral RNA of OYDV isolates were designed and synthesized (Meenakshi et al., 2006). The method was validated to detect OYDV in garlic plants . In order to regenerate virus-free garlic, meristem tip culture is a well established method for eliminating viruses from garlic (Sang-II et al., 2002; Hwang and Lee, 2008) and thermotherapy has been found in many cases to aid in garlic virus elimination (Peiwen et al., 1994). Electrotherapy was then recommended in cleaning of Cucumber mosaic virus (CMV), Arabia mosaic virus (ArMV), Grapevine fern leaf virus (GFLV), Chicory yellow mottle (ChYMV) and Tobacco mosaic virus (TMV). Based on these results, Hernandez et al. (1999) built a device to apply electric current in the cleaning of a viral complex in garlic. Other applications of the same technology have been reported in potato (Mahmoud et al., 2007; Dhital et al., 2008), grapevine (Guta et al., 2010) and banana plants (Hernandez et al., 2002) for Potato leaf-roll virus (PLRV) and Potato virus Y (PVY), Grapevine leaf roll associated virus (GLRaV) and Banana Streak Virus (BSV), respectively.
This study aimed to identify the OYDV infecting garlic in Egypt by using viral cDNA cloning, sequencing and phylogenetic analysis of OYDV and also to find appropriate methods of virus elimination using tissue culture techniques together with other treatments.
MATERIALS AND METHODS
Virus isolate and plant materials: OYDV-garlic (Allium sativum L. cv. Balady) cloves used in this study was obtained from Virology laboratory, Faculty of Agricultutre, Ain Shams University. (Mahmoud et al., 2007). Garlic cloves were cultivated under greenhouse (Fig. 1a) to obtain materials for further studies.
The virus isolate was re-identified depending on symptomatology and differential hosts by inoculation using 0.05 M borate buffer, pH 8.1 containing activated charcoal (1:100, w/v) and sodium diethyldithiocarbamate (1:1000, w/v). Also, serologically by I- ELISA using OYDV-specific polyclonal antibodies prepared by Mahmoud et al. (2007). Leaves and bulbs infected materials resulted in greenhouse were used in molecular detection and virus elimination, respectively.
Molecular study
Extraction of total RNA from plant tissues: Total RNA was isolated from the infected garlic plants using Simply P total RNA Extraction Kit obtained from BioFlux according to manufacture ’s instructions.
PCR primers: The PCR primers obtained from Meenakshi et al. (2006) and the designed for two conserved regions among the OYDV genome were used. One primer present at the 3`-end of RNA-dependent RNA polymerase gene (forward primer) and the other one present in the 3`-UTR region (reverse primer). The sequence of the reverse primer designed from the 3`-UTR region was OYDVVKBR-5`-GTCTCYGTAATTCACGC-3` with degeneracy at one point. The sequence of forward primer designed from RNA-dependent RNA polymerase gene was OYDVVKBF-5`- ATAGCAGAAACAGCTCTTA-3` (Meenakshi et al., 2006).
Reverse transcription-polymerase chain reaction (RT-PCR): Total RNA extracted from infected garlic plants was used as template for RT-PCR amplification reaction using QIAGEN One Step RT-PCR Kit. Reverse transcription reaction started with incubation at 5°C for 30 min, followed by denaturation at 95°C for 15 min. PCR amplification was performed by 30 cycles in a thermal cycler starting with denaturation at 95°C for 1 min, primer annealing at 48°C for 45 sec and extension at 72°C for 1 min with final extension at 72°C for 10 min. Five microliters aliquots of RT-PCR products were analyzed on 1% agarose gels in 0.5X TBE buffer. One killobite DNA ladder (Fermentas) was used to determine the size of RT-PCR products. Gels were stained with ethidium bromide and visualized by UV illumination using Gel Documentation System (Gel Doc 2000, Bio-Rad, USA). The expected size of the PCR product was 1.1 kb.
Cloning and sequencing of RT-PCR product: Amplified fragment covering the RNA dependent RNA polymerase gene, CP gene and 3`UTR from Egyptian isolate were extracted using Gel Extraction kit (QIAGEN). The PCR product was ligated into pGEM®-T Easy vector (Promega, USA) and the recombinant plasmids were introduced into E. coli strain DH5a as described by manufacture’s instructions. The nucleotide sequence of clones having ~ 1.1 kb insert were selected for dideoxy sequencing. The nucleotide sequence was compared and analyzed using DNAMAN Sequence Analysis Software (Lynnon BioSoft. Quebec, Canada) with those of OYDV isolates available in GenBank.
Virus-free garlic production
Plant materials and tissue culture: Garlic plants cultivated under greenhouse conditions were harvested. Bulbs were air-dried and stored for two months before treatments, except on electrotherapy treatment, whereas the harvested bulbs were treated directly then air-dried and stored. The infection of harvested bulbs with OYDV was 100% as estimated by I-ELISA. Bulbs were split into individual cloves and the outer dry papery scales removed. Cloves were surface sterilized for 2 min in 70% ethanol and 15 min in 15% sodium hypochlorite with 2-3 drops of Tween 80 and then washed three times (2 min for each one) in sterile water. Isolated basal plates were used as explants and cultured on MS basal medium containing 0.2 mg L-1 BA. After 4 weeks of growth on initiation media, explants were transferred to shoot multiplication media containing 0.5 mg L-1 BA for 6 weeks (2 subcultures).
Therapy treatments
Meristem tip culture: Meristem culture was conducted from basal plates explants cultured on MS basal medium containing 0.2 mg L-1 BA as previously reported. The shoots were excised from basal plates and were cut to about 2 mm. Meristem tips were excised in sterilized conditions under a binocular, measured along their base (meristem size) and plated on MS basal medium without plant growth regulators. After 4 weeks of growth on initiation media, explants were transferred to multiplication media containing 0.4 mg L-1 BA.
Thermotherapy: Two temperature regimes 37±1 and 38±1°C were used to analyze the effect of therapeutic treatments on viral elimination. The micropropagated shoots were cultivated in the shooting media then subjected to the previous temperature and incubated for 3 weeks at 16 h light and 8 h dark photoperiod.
Electrotherapy: The whole bulb was electric shocked by connected it to the electrodes for electric current intensity-time combinations: 5, 10 or 15 milliamper (mA) for 5 or 10 min according to Mahmoud et al. (2007). Electricity was supplied by electrophoresis power supply (LABCONCO power supply 433- 3240). Shoot apices were excised from cloves and cultured as previously reported.
Chemotherapy: Virazol was used to analyze the effect of antiviral compound on viral elimination; the micro propagated shoots were cultivated in the shooting media supplemented with 10, 20, 30 and 40 mg L-1 of virazol for 3 weeks.
Combined therapy: For study the combination effects between chemo and electrotherapy the 15 mA/10 min, electric-treated cloves were removed from bulbs and basal plates were isolated as mentioned above. Basal plates were cultured in shooting media supplemented with 20 mg L-1 of virazol and the others were cultured on basal MS media and then subjected to thermotherapy as mentioned above (Fig. 4). After the end of each treatment, samples were tested by I-ELISA then shoot clumps were transferred to control media and then dissected into single shoots and placed on MS medium supplemented with 0.2 mg L-1 NAA for root induction (Fig. 5).
Virus detection: After 3 weeks survive plantlets were indexed for OYDV by I-ELISA. Polyclonal antibody was prepared on previous work (Mahmoud et al., 2007). Then the percentages of survival and virus elimination were calculated. Therapy efficiency % was calculated by survival plantlets percentage multiply by virus-free plantlets percentage according to Mahmoud et al. (2007).
RESULTS
Isolate verification: OYDV-G isolated from garlic infected plants showing irregular yellow striping to complete yellowing and downward curling, crinkling and stunting (Fig. 1a). Samples gave positive reactions with indirect-ELISA (I-ELISA) were mechanically inoculated on Ch. amaranticolor leaves and gave chlorotic local lesions (Fig. 1b). Lesions were extracted and inoculated on Narcissus sp. for propagation (Fig. 1c). By ISEM in the same sample extracts, it was observed filamentous virus particles (Fig. 1d).
 | |
| Fig. 1(a-d): | Symptoms induced by OYDV. (a) Garlic plant showing yellow strips, curling and stunting; (b) Ch. amaranticolor showed chlorotic local lesions; (c) Narcissus sp. showed yellow stripes and (d) OYDV occurs as filamentous particles in sap |
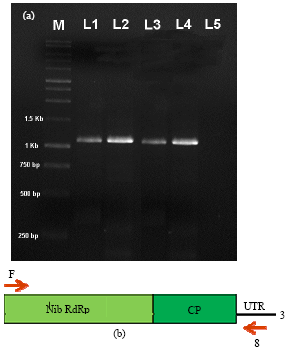 | |
| Fig. 2(a-b): | (a) Schematic diagram showing the PCR prime’s positions. (b) Agarose gel electrophoresis analysis of RT-PCR amplified products. M: 1 kb DNA ladder (Fermentas); L1 to L4: Garlic samples infected with Egyptian isolate of OYDV, L5: Healthy garlic sample (negative control) |
RT-PCR: The used PCR primers amplified one fragment covering three different regions among the viral genom. The amplified PCR fragment included the viral RNA dependent RNA polymerase gene, CP gene and 3`UTR as shown in Fig. 2a. The PCR amplification was carried out using the total RNA isolated from infected garlic plants. Electrophoresis analysis of RT-PCR product showed a single amplified fragment of ~ 1.1 kb and no fragments were amplified from the RNA extracted from symptomless or healthy plants (Fig. 2b).
Cloning of RT-PCR fragment into pGEM®-T Easy vector: The pGEM®-T Easy Vector System is convenient system for the cloning of PCR products. This vector is characterized by adding a 3` terminal thymidine to both ends. These single 3`-T overhangs at the insertion site greatly improve the efficiency of ligation of a PCR product into the plasmids by preventing recircularization of the vector and providing a compatible overhang for PCR products generated by certain thermostable polymerases. This allows PCR inserts to ligate efficiently with the vector (Mezei and Storts, 1994; Robles and Doers, 1994).
Isolation of recombinant plasmids: Several white colonies resistant to ampicillin were selected to test for recombinant plasmids containing the OYDV-PCR product. Restriction enzyme digestion with Eco RI released the cloned gene at the expected size.
Sequence analysis: Nucleotide sequencing of the RT-PCR amplified fragment in the recombinant plasmid was completed to determine if this RT- PCR fragment was OYDV or not and to compare the sequence from this isolate with those of other OYDV isolates available in GenBank.
 | |
| Fig. 3: | A phylogenetic tree showing relationships among reported isolates of OYDV and the Egyptian isolate based on the nucleotide sequences. Horizontal distances indicate degree of relatedness |
The nucleotide sequence of the Egyptian isolate of OYDV was submitted in the GenBank under Accession No. HM473189. Multiple sequence alignment of the nucleotide sequence of OYDV Egyptian isolate (HM473189)] with the corresponding sequence of ten different OYDV isolates available in GenBank [Japan (AB000840 and AB000841); China (AJ292224 and FJ765739); UK (AJ293278 and AJ409310); India (EU045556 and EU045558); Australia (DQ925454) and Argentina (X89402)] were analyzed using DNAMAN software (Fig. 3). Sequence comparisons showed the percentage of similarity ranged from 82-96% of the ten reported isolates of OYDV with the Egyptian isolate of OYDV. The similarity of the nucleotide sequences suggested that the architecture of the potyviruses is highly conserved.
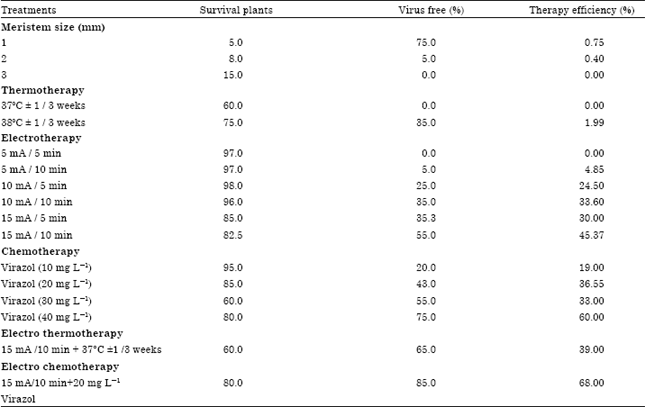
Plant regeneration and virus eradication: To investigate the effect of meristem size, thermal, chemical and electrical shock treatments on the generation rate of plantlets during the tissue culture, the regeneration and growing pattern of each regenerated plantlets were observed. As indicated in Table 1, the rate of regeneration was retarded by using small sizes of meristems and thermal treatments. The rate of regeneration and growth of shoots were not greatly retarded by treatment of virazol supplemented in the culture media. Electrical shock treatment enhancement the regeneration rate until using 10 mA/10 min, then decreased after treatment with 15/5 and 15/10 (mA/min). On the other hand, electrical shocked plant materials and then cultured on virazol (20 mg L-1) resulted 80% as are generation rate and good survival especially when 15 mA current was used (Table 1; Fig. 4). Where as, the combination of electrotherapy (15 mA/10 min) with thermotherapy (37°C for 3 weeks), the survival plants percentage reached to 60 % only.
| Table 1: | Final assessment of efficiency of virus elimination treatments for OYDV eradication from garlic plants |
 | |
 | |
| Fig. 4: | Effect of Electrotherapy and virazole antivirals on in vitro propagated garlic plantlets |
Multiplication and rooting of the treated plantlets: The treated shoot clumps were transferred to control media (Fig. 5a) and then dissected into single shoots and placed on MS medium supplemented with 0.2 mg/LNAA for root induction (Fig. 5b).
Therapy efficiency (TE): The greatest TE, 68%, was obtained from garlic cloves which exposed to 15 mA for 10 min, then shoot apices were excised and cultured on media supplemented with 20 mg L-1 of virazol. This is a result of 80% regenerated plantlets of which, 85% were (virus-free Table 1).
 | |
| Fig. 5(a-b): | Micropropagation stages of treated garlic plantlets. (a) Multiplication stage and (b) Rooting stage |
DISCUSSION
Garlic is an economically important crop for several Egyptian agricultural regions. Egyptian growers traditionally produce their own garlic propagative material. This fact accounts for the observed heavy viral infection and implies a potentially high reduction in yield and quality of this crop. If the farmers use their own material as it happens in Egypt, 100% infection is most likely to occur. To face this problem, a strategy for production of virus-free garlic propagative material was needed. The occurrence of OYDV infecting garlic plants in Egypt has been reported previously (El-Kewey et al., 2004; Mahmoud et al., 2007). Our study extends the information provided in earlier reports to viral occurrence in onion and garlic plants in Egypt. In contrast to other virus genera, serology is not a very good parameter for virus differentiation among viruses of the genus Potyvirus, as serological cross reactions often cause misinterpretation of results (Conci et al., 1999). Although, serology can be used for Potyvirus detection, it is not suitable for potyvirus taxonomy (Shukla and Ward, 1988). These observations support the application of molecular techniques for characterization of the garlic viruses, as demonstrated by others (Lot et al., 1998; Tsuneyoshi et al., 1998). The sequence of the coat protein gene has been used as an efficient tool in defining the Potyvirus species (Shukla and Ward, 1988). The specific primers from RNA-dependent RNA polymerase region and 3'-UTR successfully detected OYDV in infected garlic plants. This result confirmed our previous findings in 2007 (Mahmoud et al., 2007) when used specific primer for amplification of common central region of OYDV cp gene which produced an amplicon of 601 bp in all the samples indicating the presence of OYDV in the tested samples. Peiwen et al. (1994) reported that yields of virus-free garlic increased 25 to 80 and 35 to 89%, respectively, compared with infected garlic. However, re-infection in plants in the field is the major factor that discourages use of virus-free clones. Traditional techniques applied for cleaning of viral diseases in plants, i.e., meristems culture, thermotherapy and chemotherapy fail to produce enough quantities of clean material in most species. Alternative procedures using electric current treatments have become in an efficient tool to overcome this problem. Black et al. (1971) demonstrated a relation between growths stimulation in tomato plants that were treated with low current densities (3-15 μA/plants) during 4, 5, 12 and 24 h and the ion concentrations detected. Quacquerelli et al. (1980) applied electric current to Cactanucia tree stakes showing intense mosaic symptoms caused by virus, proving that treatments of 500 V/5-10 min lead up to 90% of cleaned plants. Those results settled the basis for the electrotherapy concept. It's known that molecular structures, protein or nucleoprotein, could be redefined as molecular machines. The molecular machines inside the cells can be treated, or they behave, as harmonic oscillators in thermal bathroom. If the viral particles behave as oscillators, then we should hope the population's of viral particles half energy is proportional to the magnitude RT, where R is the gases constant and T is the absolute temperature, because RT is the media energy, for particle mole, for a oscillator in balance in a thermal bath (Schneider, 1991). In particular, we assume that the viral particles behave as quantum oscillators, that is to say, they can take alone discreet securities of energy. Starting from this reasoning a theoretical model is developed to find the dependence between the absorbance and the electrical power in each explant. In conclusion, the study suggests that RT–PCR-based detection using specific primers from conserved regions of OYDV is a more sensitive technique than ELISA, based on our previous findings.
Also meristem culture, thermotherapy and chemotherapy are not efficient techniques for OYDV eradication from garlic, whereas electrotherapy seems to be more attractive and alternative method. It avoids the time- consuming meristem excision especially when combined with chemotherapy. Results might lead to wider application for the eradication of other viruses from other hosts.
REFERENCES
- Black, J.D., F.R. Forsyth, D.S. Fensom and R.B. Ross, 1971. Electrical stimulation and its effect on growth and ion accumulation in tomato plants. Can. J. Bot., 49: 1809-1815.
Direct Link - Chen, J., H.Y. Zheng, J.F. Antoniw, M.J. Adams, J.P. Chen and L. Lin, 2004. Detection and classification of allexiviruses from garlic in China. Arch. Virol., 149: 435-445.
CrossRef - Dhital, S.P., H.T. Lim and B.P. Sharma, 2008. Electrotherapy and chemotherapy for eliminating double-infected potato virus (PLRV and PVY) from in vitro plantlets of potato (Solanun tuberosum L.). Hort. Environ. Biotechnol., 49: 52-57.
Direct Link - Dovas, C. I., E. Hatziloukas, R. Salomon, E. Barg, Y. Shiboleth and N.I. Katis, 2001. Incidence of viruses infecting Alliums sp. in Greece. Eur. J. Plant Pathol., 107: 677-684.
Direct Link - Gibbs, A. and A. Mackenzie, 1997. A primer pair for amplifying part of the genome of all potyvirids by RT-PCR. J. Virol. Methods, 63: 9-16.
PubMedDirect Link - Guta, I.C., E. Buciumeanu, R.N. Cheorghe and A. Teodorescu, 2010. Solutions to eliminate grapevine leaf roll associated virus serotype 1+3 from V. vinifera L. cv. Ranai Magaraci. Romanian Biotechnol. Lett., 15: 72-78.
Direct Link - Lot, H., V. Chovelon, S. Souche and B. Delecolle, 1998. Effect of Onion yellow dwarf and leek yellow stripe viruses on symptomatology and yield loss of three French garlic cultivars. Plant Dis., 82: 1381-1385.
Direct Link - Meenakshi, A., V.K. Baranwal, Y.S. Ahlawat and L. Singh, 2006. RT-PCR detection and molecular characterization of onion yellow dwarf virus associated with garlic and onion. Curr. Sci., 91: 1230-1234.
Direct Link - Pappu, S.S., R. Brand, H.R. Pappu, E.P. Rybicki, K.H. Gough, M.J. Frenkel and C.L. Niblett, 1993. A polymerase chain reaction method adapted for selective amplification and cloning of 3' sequences of potyviral genomes: Application to dasheen mosaic virus. J. Virol. Methods, 41: 9-20.
CrossRefDirect Link - Schneider, T.D., 1991. Theory of molecular machines. I. Channel capacity of molecular machines. J. Theor. Biol., 148: 83-123.
PubMed - Shahraeen, N., D.E. Lesemann and T. Ghtbi, 2008. Survey for viruses infecting onion, garlic and leek crops in Iran. OEPP/EPPO Bull., 38: 131-135.
CrossRef - Shukla, D.D. and C.W. Ward, 1988. Amino acid sequence homology of coat proteins as a basis for identification and classification of the Potyvirus group. J. Gen. Virol., 69: 2703-2710.
CrossRef - Takaichi, M., T. Nagakubo and K. Oeda, 2001. Mixed virus infections of garlic determined by a multivalent polyclonal antiserum and virus effects on disease symptoms. Plant Dis., 85: 71-75.
Direct Link - Tsuneyoshi, T., T. Matsumi, K.T. Natsuaki and S. Sumi, 1998. Nucleotide sequence analysis of virus isolates indicates the presence of three Potyvirus species in Allium plants. Arch. Virol., 143: 97-113.
PubMed