
A.A. Ahmed
Department of Virus and Phytoplasma Research, Plant Pathology Research Institute, Giza, Egypt
M.M. Fath-Allah
Department of Virus and Phytoplasma Research, Plant Pathology Research Institute, Giza, Egypt
International Journal of Virology
Year: 2012 | Volume: 8 | Issue: 1 | Page No.: 50-60







ABSTRACT
Cucumber Mosaic Virus (CMV) and Plum Pox Virus(PPV) (Sharka) were isolated from naturally infected apricot (Prunus armeniaca cv. El-Amar) trees showing leaf mottling, marginal leaf necrosis deformation and abortive flowers grown at Kalubia and el-Fayoum Governorates, Egypt. The identities of the two viruses were confirmed serologically using DAS-ELISA. Graft inoculation of diseased apricot scion onto healthy one showed the same symptoms as those seen in naturally infected trees in the stock shoots. Similar symptoms were caused by slashing inoculation with surgical knife onto the stem of young healthy apricot seedling with mixed inoculum of both viruses of CMV and PPV. CMV was biologically purified from single local lesions which formed on cucumber cotyledon and/or Chenopodium quinoa. Identification of this virus was based on host range, transmissibility and serological tests. CMV was able to infect only 20 plant species and varieties out of 24 tested plants by either grafting or mechanical inoculation and PPV was able to infect only 19 plant species and varieties. CMV and PPV can be transmitted by Aphis gossypii and Myzus persicae from cucumber to cucumber and woody plants. The best time for leaf sampling was to detect the viruses, in diseased apricot tree by sap inoculation or ELISA, in April, followed by May and June. Double infection by PPV and CMV of apricot trees resulted in synergistic effect where symptoms in doubly infected plants were more severe than that in plants singly infected by either viruses. ELISA values revealed that the concentration of CMV in doubly infected apricot plants is significantly higher than that of single infected plant. Increase of CMV in doubly infected apricot was confirmed by local lesion assays and specific infectivity. However, PPV titer did not increase significantly in case of double infection compared to singly infected plant. Reverse transcription- polymerase chain reaction (RT-PCR) amplified 500 bp fragment and 335 bp from infected plant using degenerate oligonucleotide primer specific for potyviruses group and oligonucleotide primer specific for CMV. The results showed also that such multiplex RT-PCR can detect the two viruses either each alone or both together in samples collected from artificially or naturally infected plants.
PDF Abstract XML References Citation
Received: June 20, 2011;
Accepted: July 27, 2011;
Published: October 25, 2011
How to cite this article
A.A. Ahmed and M.M. Fath-Allah, 2012. Double Infections with Cucumber Mosaic Virus and Plum Pox Virus (Sharka) in Apricot Trees. International Journal of Virology, 8: 50-60.
DOI: 10.3923/ijv.2012.50.60
URL: https://scialert.net/abstract/?doi=ijv.2012.50.60
DOI: 10.3923/ijv.2012.50.60
URL: https://scialert.net/abstract/?doi=ijv.2012.50.60
INTRODUCTION
Diseases caused by virus or virus-like agents have induced heavy losses world-wide annually in stone fruit trees (Nemeth, 1986; Roistachecr, 1992). In fruit trees, more than 150 diseases caused by viruses, viroids, phytoplasmas and unknown graft transmissible, agents were reported. To control these diseases, it is important to identify the causal agent and to determine its properties.
This has been possible in certain recalcitrant disease agents with recent advances in serological and molecular techniques (Candresse, 1995). Nevertheless, biological indexing on herbaceous and woody plants remains as an essential tool for identification and characterization of new agents and diseases in certification programmers producing virus-free propagating materials.
Cucumber Mosaic Virus (CMV), the type species of the genus Cucumovirus (family Bromoviridae) (Rybicki, 1995) is a virus of many economically important crops worldwide and found to be able to infect over 1000 plant species from 85 families, including dicotyledons, monocotyledons, herbaceous and woody plants, horticultural crops and wild plants (Palukaitis et al., 1992). Sharka disease was recorded for the first time in Bulgaria in 1918 on plum trees and then in 1932 on apricot trees (Atanasoff, 1932). The virus is transmitted by mechanical, grafting and by aphids in a nonpersistent manner, inducing fast spreading epidemics, mainly in apricot, peach and plum orchards. Plum Pox Virus (PPV) is one of the most detrimental viral diseases of Prunus stone fruit crops in Europe and the Mediterranean Basin Roy and Smith (1994) and it was first found in Egypt in 1987 (Dunez, 1987, 1988, 1989). The virus is transmitted by rafting and by aphids in a non persistent manner, inducing fast spreading epidemics, mainly in apricot, peach and plum orchards (Mazyad et al., 1992).
PPV is a member of the genus Potyvirus (family Potyviridae) presents and has a single stranded genomic RNA (Dallot et al., 2001). The disease causes heavy losses for fruit growers and the international trade of propagation materials and fresh fruits. It is considered one of the most detrimental viral diseases of Prunus stone fruit crops in Europe and the Mediterranean Basin. It was also recently identified in Chile and the United States and has been reported from Cyprus, Syria, Egypt, India, Kazakhstan, Chile, the United States and Canada which causes the economically important sharka disease on stone fruit trees.
The present work is focused on isolation and identification of CMV and PPV from the apricot trees using enzyme-linked immunosorbent assay and detection of CMV and PPV by RT-PCR and detection of CMV and PPV by multiplex RT- PCR.
Also, ELISA is used to monitor the virus titer in singly and doubly infected apricot seedlings grown under green house conditions. The relationship between virus concentration and symptoms severity of doubly infected plants is discussed.
MATERIALS AND METHODS
Isolation and host range of CMV and PPV
Virus source: Inoculum sources were obtained from young apricot leaves showing deformation, curling and mild mosaic were collected from P. armeniaca (cv.El-Amar) in April. Leaves from the original source plant which is obtained from Kalubia governorate. These leaves were ground in a cooled porcelain morter in 5 parts of cold phosphate solution (0.03 M Na2H•PO4 containing 0.2% Na-diethyl dithiocarbamate), pH 7.4 [27-14]. The extracts were inoculated to young, tender leaves of test plants with an absorbent cotton swap by carborundum leaf-inoculation method. After inoculation, plants were kept in partly shaded greenhouse at temperature ±25°C.
Single local lesion technique accompanied with successive dilutions was used to isolate each one in pure form of each virus isolate. For the single lesion isolation, CMV virus from a necrotic local lesion developed on Chenopodium quinoa leaves and PPV from a chlorotic local lesion produced on Chenopodium foetidum leaves (Aboul-Ela et al., 1999). Back inoculation tests of all inoculated plants were carried out by inoculation to C. quinoa, Vigna unguiculata cv. Black eye and other differential hosts and serological tests.
Host range and symptomology: Twenty four plant species belonging to 8 families were used to study the host range of the CMV and PPV. Ten seedlings of each tested plant species or cultivars were mechanically inoculated with each virus and kept under greenhouse conditions. Plants were examined daily, for four weeks for symptoms expression. Inoculated plants that did not show any symptoms were checked for latent infection by back-inoculation to the indicator host Chenopodium foetidum and Chenopodium quinoa.
Viruses inoculation in apricot: Amar apricot seedlings were obtained from the Horticulture Institute, ACR. Plants were tested for eight known viruses Apple Chlorotic Leaf Spot Virus (ACLSV), Arabis Mosaic Virus (AMV), Cucumber Mosaic Virus (CMV), Prune Dwarf Virus (PDV), Peach rosette Mosaic Virus (PRMV), Prunus Necrotic Ring Spot Virus (PNRSV), Plum Pox Virus (PPV) and Tomato Ring Spot Mosaic Virus (TomRSV) by DAS-ELISA. Two different mechanical inoculation procedures, rub inoculation with carorandum and slashing inoculation onto green stem were used. N. tabacum cv. White Burly leaves infected with PPV and Cucumis sativas leaves infected with CMV were homogenized in each 4 volume of extraction buffer. These homogenates were used as inoculum for each test. A surgical knife was dipped into the inoculums and up to 25 slashes per seedlings were made onto about 30 cm length of young green stems. Single inoculation with CMV or PPV-Amar and mixed inoculation with inoculum of both viruses (1:1) were conducted onto stem of 5 seedlings each in May. At the same time, extraction buffer was inoculated to 5 seedlings as negative control. Plants were maintained in a plant green house (±25°C) until September and transferred to outside open-air through January and February.
Insect transmission: Aphid transmissibility of the present viruses was evaluated under controlled conditions on four weeks old seedlings of N. tabacum cv. White Burly and one year old apricot cvs. Local and Hammawy. Colonies of aphid species used in tests were collected in March and April, at the ARC (Horticulture Institute). Myzus persicae were collected from peach orchard from trees and associated herbaceous ground cover in and around the orchard and Aphis gossypii were collected from a bean field at the ARC. Also, in addition virus- free aphids were originally obtained from the stock culture of Aphid Section, Plant Protection Research Institute, ARC; Giza.
Serological reaction: Leaf samples were collected randomly from symptomatic as well as asymptomatic apricot leaves. Samples were stored at 4°C until processed. The standard double-antibody sandwich enzyme- linked immuno-sorbent assay (DAS-ELISA) was used for detection of PPV and CMV. IgG and conjugated IgG for the two different viruses were purchased from Sanofi, Sante Animal, Paris, France. Extraction of infected leaves and virus detection by ELISA were applied as described by Clark and Adams (1977) and optimized in the laboratory. Each sample was replicated in two contiguous wells and healthy samples corresponding to the appropriate Prunus spp. were added at three replicates per plate. PPV-infected leaves of Nicotiana tabacum were used as a positive control. The positive threshold was set at twice the mean of absorbance values (A405) obtained with the healthy samples. In some experiments, virus detecting was also performed by RT-PCR.
Detection of CMV and PPV at various times through the year by sap inoculation and ELISA: As the virus detection from apricot was not always easy, we performed the detection tests in different seasons to determine the best season for sampling. During April 2008 to March 2009, leaves and flower buds samples of the same diseased apricot trees showing the symptoms were collected. from El-Fayoum Governorate several times. The samples were tested by ELISA to detect the viruses as described earlier.
Specific infectivity of CMV and PPV: The effect of age of infection on PPV and CMV titer in singly and doubly infected apricot cv. El- Amar seedling were evaluated by ELISA. Infectivity assay of PPV and CMV from singly or doubly infected apricot seedlings were performed on the local lesion hosts Chenopodium quinoa and Chenopodium amaranticolor for CMV and PPV, respectively.
Reverse transcription polymerase chain reaction (RT- PCR)
Isolation of total nucleic acid: RNA Minipreps super kit type (EaZy Nucleic Acid isolation) (E-Z-N-A). Fifty milligram of healthy and infected apricot tissue were ground under liquid nitrogen to a fine powder using a sterile mortar and then transferred to microfuge tube. A 600 μL QVL lysis buffer were added, vortexed vigorously and incubated at room temp. for 30 min and centrifuged at 10,000 g for 10 min. Supernatant was transferred to a new microfuge tube without disturbing the cell-debris in collection tube, then 350 μL absolute ethanol (96-100%) were added to the cleared lysate and mixed thoroughly by vortexing for 30 sec and centrifuged briefly to collect any liquid droplets from lid. The mixture was placed in HiBind® RNA column with 2.0 mL collection tube and transferred to the column and centrifuged for 1 min at 10,000 g. The flow-through fraction from the HiBind® RNA column was discarded and the column back to the same collection tube. Seven hundred micro liters of wash buffer were added to the column and centrifuged for 1 min at 10,000 g then the flow-through was discarded and transferred into a new collection tube. Five hundred microliters of wash buffer 11 diluted with ethanol were added onto the column, centrifuged for 1 min at 10,000 g and centrifuged once more for 1 min at 10,000 g to remove the residue of wash buffer 11. The column was transferred into clean 1.5 microfuge tube. To elute the RNA, 50 μL of Diethylpyrocarbonate (DEPC)-water (RNase free) were pipetted directly onto the center part of the membrane in the column and incubated at room temp. for 5 min and centrifuged for 1 min at 10,000 g then the RNA kept at-80°C for long term storage. The oligonucleotide primers used for single or multiplex RT-PCR are shown in Table 1. RT-PCR was done using Moloney-Murine Leukemia Virus reverse- transcriptase (M-MLV) Metabion international AG, Lena-Christ- Stasse 44/1, D-82152 Martinsried/Deutschland. RT and PCR are carried out sequentially in the same tube (Bariana et al., 1994). All components required for both reactions are added during set up. First strand cDNA synthesis was done using the following parameters. The 50°C for 30 min at 1 cycle to allow reverse transcription to be preformed. Second strand synthesis of cDNA and PCR amplification were done in the same tube using DNA thermal cycler with the following parameters: 95°C for 15 min at 1 cycle. The amplification parameters: denaturation at 94°C for 1 min, primer annealing at 55°C for 1 min and extension at 72°C for 1 min, for 30 cycles with a final extension at 72°C for 10 min CMV and PPV isolated from apricot plants from El-Fayoum governorate, were used in this test.
| Table 1: | Primer sequences and expected size of PCR product for each primer pair when used to amplify the appropriate viruses (CMV and PPV in infected apricot) template |
 | |
Analysis of RT-PCR: Ten microliter aliquots of RT-PCR were analyzed on 2% agarose gels in TBE buffer (89 mM tris-HCL, 89 mM boric acid, 2.5 mM EDTA, pH 8.5) at 80 volt for 90 min. Mixture of different base pair DNA size markers (ZHBio-Tech) were used to determine the size of RT-PCR products. Gels were stained with ethidium bromide 10 μg mL-1 (agarose gels) and visualized by UV illumination.
Detection of CMV and PPV by Multiplex RT-PCR: Multiplex RT-PCR was used to detect CMV and PPV in artificially infected squash plants as well as in naturally infected squash samples collected from El-Fayoum.
The multiplex RT-PCR assays were optimized with minor modification. Several parameters such as primer concentration (from 0.1 to 1 μM), magnesium chloride concentration (from 1.5 to 4 mM) and elongation time (from 1 to 5 min) were analyzed to establish the best condition for simultaneous amplification targets of two viruses. The optimum cocktail mix prepared in a thin walled 0.2 μL PCR tube, by combining of 5 μL PCR buffer 10 x, 2.5 μL of dNTPs mix, 2 μL of 4 mM MgCl2, 1 μL M-MLV (2000 unit), 0.4 μL. Taq polymerase, 3 μL of 10 μ M of each vira sense primer, 3 μL of 10 μM of each complementary primer and 5 μL of total RNA in a total volume of 50 μL.
The reaction mixture was subjected to amplification as previously mentioned and aliquots were analyzed using mixture of different base pair DNA size markers to determine the size of RT-PCR products.
RESULTS AND DISCUSSION
Host range of the two viruses: Out of twenty four plant species and cultivars belonging to eight families, twenty plant species were susceptible with CMV and many of them were infected systemically showing different symptoms (Table 2). CMV locally infected C. quinoa and Vigna unguiculata plants. In C. foetidum irregular chlorotic spots on the inoculated leaves were observed 8 days to 14 days after the inoculation with PPV.
These results showed that two distinct viruses are associated with mosaic, line pattern, chlorotic rings and curling in apricot. The two viruses were identified as Cucumber Mosaic Virus (CMV) and Plum Pox Virus (PPV). Grafting of naturally infected apricot scion onto healthy apricot plantlets showed the same symptoms in the stock shoots indicating that the disease was graft transmissible. Mixed inoculum of CMV and PPV was able to back to apricot seedling by stem slashing inoculation onto green stem and resulted in similar symptoms, Otsubo et al. (2002).
Results of host range trials showed that CMV and PPV had a wide host range (Table 2, Fig. 1). Twenty of the plant species developed CMV symptoms from twenty two and nineteen of plant species developed PPV symptoms from twenty four. No symptomless infection was detected by DAS-ELISA in leaves. CMV produced necrotic ring spots on the inoculated leaves Chenopodium quinoa and systemic mottle on the top leaves of Chenopodium amaranticolor plants. Also, Zinnia elegans reacted to CMV with chlorosis at the base of the youngest leaf. Chlorotic and necrotic lesions were observed on cotyledons of C. sativus cv. Beita Alpha. Systemic symptoms were consisted of severe mosaic, compact growth and top necrosis. Among Leguminosae, only Vigna unguiculata reacted to CMV by forming red necrotic local lesions on the cotyledons, ten days after inoculation. In Petunia hybrida, systemic chlorotic lesions formed that eventually developed into necrotic lesions and systemic mottling was also observed in plants inoculated with CMV: Vinca rosea and Nicotiana. clevelendii exhibited systemic symptoms ranging from mild mottle to severe leaf distortion.
| Table 2: | Reaction of some hosts inoculated mechanically or grafting by CMV apricot isolate |
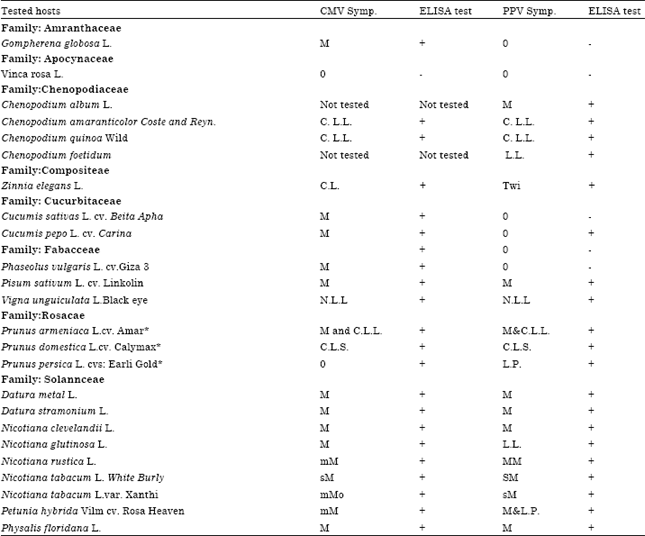 | |
| +: Positive and -: Negative in ELISA test. L.L: Local Lesion.; C.L.L: Chlorotic Local Lesion; *Grafting inoculated, C.L.S: Chlorotic Local Spots; N.L.L: Necrotic LL; Twi: Twisting; Cl: Chlorosis; M: Mosaic; L.P: Line pattern; Mo.: Mottle; mMo: mild mottle; mM: mild Mosaic; sM: Sever mosaic; 0: No symptoms; N: Necrosis and TN: Top necrosis | |
Graft transmission: The different methods of virus transmission may be useful diagnostic criteria. All viruses which are systemic in their hosts can be transmitted by grafting between susceptible and compatible plant. Typical CMV Symptoms induced on graft- inoculated seedlings of different cultivars of apricot, also in P. persica cv Earli Gold, P. domestica cv. Calymax and were generally characteristic of those previously reported for CMV (Nemeth, 1986).
Insect transmission: Data in Table 3 revealed that Aphis gossypii and Muzus persicae were able to transmit CMV and PPV from infected woody seedlings (apricot, peach and plum). M. persicae was more effective than A. gossypii in transmitting PPV and both insects were ineffective on peach and plum. These results agree with Awad et al. (2007).
Serological reaction: All of the CMV and PPV infected samples reacted positively only with antisera specific for the two viruses using DAS-ELISA.
 | |
| Fig. 1(A-H): | Symptoms of PPV and CMV on naturally and artificially infected host lants. (A). Mosaic, vein clearing and chlorotic spots appeared on naturally infected (mixed infection PPV and CMV) appeared on apricot leaves (B). PPV symptoms (Chlorotic rings) appeared on naturally infected apricot leaves (C). Symptoms on infected apricot leaves artificially infected with CMV showing mosaic and curling (D). Mosaic and chlorotic spots PPV symptoms on artificially infected leaves of apricot leaves (E). Infection of apricot leaves showing mosaic symptoms (F). Sever mosaic on celery infected with CMV (G). Necrotic ring spots in C. quinoa CMV infected leaves (H). Necotiana tabacum cv White Burly infected with CMV showing top necrosis and sever mosaic symptoms |
| Table 3: | Results of transmission test CMV and PPV by aphids from apricot to apricot, peach . (+) = Virus was transmitted. ( - ) = Virus was not transmitted |
 | |
| Table 4: | Detection of CMV and PPV at various times through the year by sap inoculation and ELISA |
 | |
| Samples were collected from leaves, except in February as flower buds. +, ++ and +++: in inoculation produced local lesion on Chenopodium quinoa for CMV and systemic Chenopodium quinoa for PPV in mechanical inoculation; in ELISA absorbance values were more two times than control, - : no symptoms on inoculated plants; ELISA absorbance were lower than negative control, 0 : not tested | |
A darker yellow color developed with the positive samples. In all tests, a visual rating of ELISA plates was in agreement with the ELISA reading. Positive reaction obtained using specific antisera against PPV and CMV at a dilution of 1:500 confirmed the identification of the viruses under study. Serological tests, such as ELISA provide rapid and convenient methods for the identification and estimation of plant viruses in leaves or buds (Nemeth, 1986; Candresse, 1995).
Detection of CMV and PPV by ELISA and sap inoculation in apricot infected trees at different growing stages: By using the ELISA assay, CMV and PPV can be detected from leaves concentration in leaves sample April and May then in the following Mach and June and in July, September and November symptoms were disappear and ELISA cannot detect the virus. However, PPV detected by symptoms and ELISA from February to July (Table 4). These results confirm the results obtained from plum (Adams, 1978; Dosba et al., 1986).
Comparative relative concentration of CMV in singly and double infected plants: The PPV relative concentration from singly and doubly infected apricot plants varied during the experiments period (Table 5). The virus relative concentration was highest 5 months post inoculation for either singly or doubly infected plants. The titer in the doubly infected plants had significantly higher CMV titer than did the corresponding singly infected plants (Table 5). The ratio of CMV titer in doubly: singly infected plants ranged from 1.6 to 3.5 (Table 5).
Comparative relative concentration of PPV in singly and double infected plants: No significant differences in PV relative concentration were detected between singly and doubly infected apricot plants (Table 6). The ratio of PPV titer in doubly to singly infected plants ranged from 1.1 to 1.5 (Table 6).
| Table 5: | Comparison of PPV relative concentration determined as reading ELISA values in leaves of apricot Prunus armeniaca cv. EL-Amar singly and doubly infected with CMV and PPV |
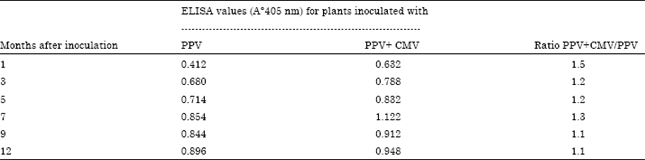 | |
| Leaf extracts of comparable leaves from plants singly infected with CMV served as controls and gave negligible ELISA values (0.042-0.050). Ratio for PPV titer in doubly: singly infected. Values are means±standard error for leaf extracts from five plants with each extract tested in duplicate wells, the plates were read with ELISA reader 30 min following addition of substrate | |
| Table 6: | Comparison of CMV relative concentration determined as reading ELISA values in leaves of apricot Prunus armeniaca cv. EL-Amar singly and doubly infected with CMV and PPV |
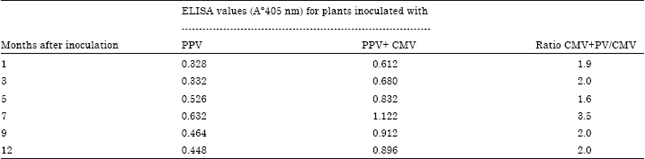 | |
| Leaf extracts of comparable leaves from plants singly infected with PPV served as controls and gave negligible ELISA values (0.038- 0.046). *Ratio for CMV titer in doubly: singly infected. Values are means ± standard error for leaf extracts from five plants with each extract tested in duplicate wells, the plates were read with ELISA reader 30 min following addition of substrate | |
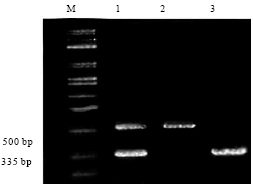 | |
| Fig. 2: | Detection of CMV and PPV by RT-PCR and multiplex-RT-PCR. Lane (M): DNA marker, (1) Mixture of CMV + PPV, (2) Individual MV and (3) Individual PPV |
Infection of Prunus species by viruses such as PPV, or CMV cause considerable damage worldwide in sever epidemic in stone fruit fields either in single or mixed infection (Perry et al., 1994). Double infection by CMV and the Potyvirus PPV resulted in synergistic effect on Prunus host apricot cv. El-Amar such response was obvious in terms of more severe symptoms expression.
A notable increase in host symptoms as a result of double infection were reported in several studies involving different combinations between Potyviruses and viruses belonging to other genera. Examples of such synergistic effect was reported between potato virus Y, Tobacco vein mottling virus or Tobacco etch virus together with Potexvirus potato virus X in Tobacco (Vance et al., 1995), Zucchini yellow mosaic virus (ZYMV)and the Cucumovirus Cucumber mosaic virus (CMV).
Detection of CMV and PPV by RT-PCR: It is very important to recognize a sensitive, simple, reliable, inexpensive method for detection of the virus in the different parts of the infected plant. Previous studies illustrated the importance and usefulness of PCR as a diagnostic tool for plant viruses (Thomson et al., 1995). The obtained results confirmed the specificity of the primers used in this study. The size of the amplification obtained product was approximately 335 bp for potyviruses (PPV) and 500 bp for CMV (Fig. 2). In case of extraction by kits the obtained bands were more distinct. A negative reaction of RT-PCR was obtained with healthy tissue.
In our study RT-PCR was performed on total nucleic acid extracted from leaf tissues infected with the studied CMV and PPV using two specific primers for each virus. Our results confirmed the specificity of the primer sequence used. The PCR product sizes were in agreement with those found by Bariana et al. (1994) for CMV and for PPV.
Detection of CMV and PPV by multiplex RT-PCR: A cocktail of primers to CMV and PPV was used to detect one or both viruses in each sample. The results showed that such assay can detect the two viruses either each alone or both together in samples collected from artificially or naturally infected plants. When samples used for detection were taken from doubly infected plants, simultaneous amplification of a 500 bp fragment of the CMV CP-gene and a 335 bp fragment of the PPV CP-gene were observed (Fig. 2). Multiplex polymerase chain reaction (m-PCR) accommodates several pairs of primers in one reaction, resulting in reduced material costs and time when compared to several individual PCR reactions. Our data showed that simultaneous amplification of a 500 bp fragment of the CMV genome and a 335 bp fragment of the PPV genome were observed. These results were in line with those of Shalaby et al. (2002) and Papayiannis et al. (2005) who used the m-PCR with other viruses. It was the first time to use this procedure to detect PPV and CMV in mixture according to the available literature.
REFERENCES
- Adams, A.N., 1978. The detection of plum pox virus in Prunus species by enzyme-linked immunosorbent assay (ELISA). Ann. Applied Biol., 90: 215-221.
CrossRef - Bariana, H.S., A.L. Shannon, P.W. Chu and P.M. Waterhouse, 1994. Detectionof five seedborne legume viruses in one sensitive Multiplex polymerease chain reaction test. Phytopathology, 84: 1201-1205.
Direct Link - Clark, M.F. and A.N. Adams, 1977. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol., 34: 475-483.
CrossRefPubMedDirect Link - Dallot, S., L. Quiot-Douine, P. Saenz, M.T. Cervera, J.A. Garcia and J.B. Quiot, 2001. Identification of Plum pox virus determinants implicated in specific interactions with different Prunus spp. Phytopathology, 91: 159-164.
Direct Link - Shalaby, A.A., M.K. Nakhla, A.M. Soliman, H.M. Mazyad, A. Hadidi and D.P. Maxwell, 2002. Development of a highlysensitive multiplex reverse transcription-polymerase chain reaction (m-RT-PCR) method for detection of three potato viruses in a single reaction and nested PCR. Arab J. Biotech., 5: 27-28.
- Thomson, K.G., R.G. Dietzgen, A.J. Gibbs, Y.C. Tang, W. Liesack, D.S. Teakl and E. Stackebrandt, 1995. Identification of Zucchini yellow mosaic potyvirus by RT-PCR and and analysis of sequence variability. J. Virol. Meth., 55: 83-96.
PubMed - Vance, V.B., P.H. Berger, J.C. Carrington, A. G.Hunt and X.M. Shi, 1995. 5 proximal potyviral sequences mediate potato virus X/potyviral synergistic disease in transgenic tobacco. Virology, 206: 583-590.
PubMed - Otsubo, T., J. Kurihara, M. Sakakibara and K. Tomaru, 2002. Confirmation of symptom reproduction in mume leaf margin necrosis disease on Prunus mume (cv.Nankou) by sap inoculation with causal two viruses, Cucumber mosaic and prunus necrotic ring spot-like virus. J. Agric. Sci., Tokyo Univ. Agric., 47: 11-16.
- Papayiannis, L.C., N. Ioannou, I.N. Boubourakas, C.I. Dovas, N.I. Katis and B.W. Falk, 2005. Incidence of viruses infecting cucurbits in Cyprus. J. Phytopathol., 153: 530-535.
CrossRefDirect Link - Perry, K.L., L. Zhang, M.H. Shintaku and P. Palukaitis, 1994. Mapping determinants in cucumber mosaic virus for transmission by Aphis gossypii. Virology, 205: 591-595.
CrossRefPubMedDirect Link - Roy, S. and I.M. Smith, 1994. Plum pox situation in Europe. Eur. Plant Prot. Organ. Bull., 24: 515-523.
CrossRefDirect Link