
A.A. Megahed
Department of Plant Pathology, National Research Centre, Dokki, Cairo, Egypt
Kh. A. El- Dougdoug
Department of Agriculture Microbiology, Faculty of Agriculture, Ain Shams University, Shoubra EL-Kheima, Cairo, Egypt
B.A. Othman
Department of Agriculture Microbiology, Faculty of Agriculture, Ain Shams University, Shoubra EL-Kheima, Cairo, Egypt
S.M. Lashin
Department of Plant Pathology, National Research Centre, Dokki, Cairo, Egypt
M.A. Ibrahim
Department of Molecular Biology, National Research Centre, Dokki, Cairo, Egypt
A.R. Sofy
Departmnet of Botany and Microbiology, Faculty of Science, Al-Azhar University,11884 Nasr City, Cairo, Egypt
International Journal of Virology
Year: 2012 | Volume: 8 | Issue: 3 | Page No.: 240-257







ABSTRACT
Cucumber mosaic cucumovirus satellite strain Egyption isolate (st-CMV-EG) was detected using polyclonal antibodies by DAS-ELISA form naturally infected cucumber leaves collected from protected agriculture. The samples which give positive reaction were used to virus isolation. st-CMV-EG isolate has a wide host-range belonging to 6 families. The virus isolate was inactivated at 70°C, it was infected at dilution 10-4 and longevity was 4 days at room temperature. Amorphous and crystalline inclusion bodies were detected in hair cells of epidermal strips of CMV-inoculated cucumber leaves. The U.V absorption ratio, A260/A280 and Amax./Amin. were 1.204 and 1.101, respectively, as well as virus yield was 1.43 mg/100 g fresh leaves in purified virus preparation. Electron micrograph of the purified virus isolate showed spherical particles (28 nm.). The virus isolate was detected serologically by using dot blot immunoassay. CMV-RNA was successfully detected in infected cucumber leaves using specific primer of cp gene by RT-PCR and the expected size was about 582 bp. Sequence analysis of CMV-cp gene of cucumber Egyption isolate was indicated 38% similarity to that of AB024493 and D00542.
PDF Abstract XML References Citation
Received: November 19, 2011;
Accepted: January 12, 2012;
Published: February 13, 2012
How to cite this article
A.A. Megahed, Kh. A. El- Dougdoug, B.A. Othman, S.M. Lashin, M.A. Ibrahim and A.R. Sofy, 2012. A New Egyptian Satellite Strain of Cucumber Mosaic Cucumovirus. International Journal of Virology, 8: 240-257.
DOI: 10.3923/ijv.2012.240.257
URL: https://scialert.net/abstract/?doi=ijv.2012.240.257
DOI: 10.3923/ijv.2012.240.257
URL: https://scialert.net/abstract/?doi=ijv.2012.240.257
INTRODUCTION
Cucumber mosaic virus (CMV), a member of the family Bromoviridae (genus Cucumovirus), has an extremely wide host range, including more than 1200 plant species in 500 genera of 100 families (Tobias et al., 1982). It causes a broad spectrum of symptoms from mild to severe mosaic, stunting, chlorosis, necrosis and filiformism depending on virus strain and the host (Carrere et al., 1999).
The satellite virus had no serological relationship with helper virus or other satellite viruses. These experiments are a good example of the need for care when attempeting to delineate a newly isolated satellite RNA (Valverde and Dodds, 1987). Cucumber mosaic virus supports satellite RNAs consisting of single-strand, linear RNA molecules of 330 to 390 nucleotide. The satellite RNAs of often attenuated the symptoms induced by CMV in certain host plants (Hidaka et al., 1988), but in some cases quite dramatic increases in symptom severity may occur (Kaperi et al., 1988).
Sharma et al. (2005) reported that, CMV was detected and characterized by bioassay, double antibody sandwich enzyme linked immunosorbent assay (DAS-ELISA), electron microscopy, immunosorbent electron microscopy, cytopathology, reverse transcription polymerase chain reaction (RT-PCR) and sequencing.
RNAs 1 and 2 code for proteins that are associated with the replication of viral genome (Hayes and Buck, 1990). RNA 2 also encodes an additional protein that is a suppressor of post transcriptional gene silencing and affects multiple functions including long distance movement and host range (Brigneti et al., 1998). RNA 3 is dicistronic, coding for both the 3a movement protein (Kaplan et al., 1995) and the Capsid Protein (CP), the latter being translated from a subgenomic messenger (RNA 4) (Schwinghamer and Symons, 1977).
This present study aims to isolation and identification of st-CMV isolate biologically, serologically and molecular characters from infected cucumber plants under protected greenhouse conditions in Egypt.
MATERIALS AND METHODS
Source of virus isolate: Ten samples of infected cucumber plants (Cucumis sativus cv. Barakoda) showing distincted viral symptoms were collected from protected greenhouses and used for st-CMV isolation.
The virus was detected using polyclonal antibody specific CMV by DAS-ELISA (kits provided by Ssnofi, Sante, Animal, Paris, Farance) according to Clark and Adams (1977).
Isolation and propagation of virus isolate: The infected cucumber plants which gave +ve reaction against IgG-CMV by DAS-ELISA were used for virus isolation. The inoculum of infectious sap was mechanically inoculated on Chenopodium amaranticolor as indicator host. The single local lesion assay was used for biological purification of isolate and propagated on healthy Nicotiana glutinosa plants as CMV propagative host.
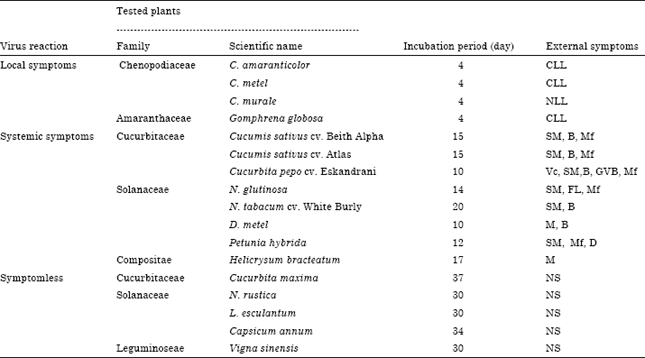
Host range and symptomatology: Eighteen plant species belonging to 6 families (Cucurbitaceae, Solanaceae, Chenopodiaceae, Leguminoseae, Compositae and Amaranthaceae). Table 1 were mechanically inoculated with virus inoculum using five plants of each host. The results were confirmed by DAS-ELISA test against IgG-CMV.
Virus stability: Thermal Inactivation Point (TIP), Dilution End Paint (DEP) and Longevity in vitro (LIV) was performed according to Noordam (1973), using C. amaranticolor as local lesion host to CMV.
Inclusions bodies: Crystalline Inclusion Bodies (CIB) were examined in the epidermal strips from the lower surface leaves of CMV-infected cucumber plant (15 days after inoculation). As well as the amorphous inclusion bodies using bromophenol blue and mercuric chloride. The strips were examined under light microscope, magnification of 400-X (Mazia et al., 1953).
Virus purification: The particles of virus isolate were purified by the method described by Murant (1965). The purified isolate was diluted in extraction buffer 10-1 and measured at range 230-300 nm by spectrophotometer (Shimadzu UV-2401 PC UV-Vis) at Molecular Biology Lab., National Research Centre (NRC). The virus yield determined using an extinction coefficient of 5 for CMV (Noordam, 1973).
Negative staining method: Morphology particles of the virus isolate was examined using negative stain method according to Noordam (1973) by JOEL-TEM-1010 electron microscope unite, electron microscope Department, NRC.
Dot blot immunoassay (DBIA): it was used for identification of virus isolate from infected cucumber as described by Lin et al. (1990) using polyclonal antibodies (CMV-IgG).
Molecular studies
Extraction of total RNA (T-RNA): Total RNA was extracted from infected cucumber leaves by using a method described by Gibbs and Mackenzie (1997). The extracted RNA was stored at -80°C.
cDNA synthesis: Ten microliters of total RNA were added to mixture contained 6 μL of 5X first strand buffer (250 mM Tris-HCl pH 8.3, 375 mM KCl and 15 mM MgCl2), 3 μL of 0.1 M dithiothreitol (DTT), 1 μg of complementary specific primer CP/CMV and sterile H2O to a final volume of 30 μL. The primer annealed at 65°C for 30-45 min. The annealed reaction containing 20 μL of cDNA: 4 μL of 5X first strand buffer, 2 μL of 0.1 M DTT, 1 μL of RNAsin (40 units, Promega Corp., Madison, US), 5 μL of 0.3 M β-mercaptoethanol, 2.5 μL of 10 mM dNTPs and 1 μL of Moloney murine leukemia virus (MMLV) (200 U μL-1) reverse transcriptase (Promega, Corp.). Reaction was mixed briefly and incubated for 1-1.5 h at 42°C (Hsu et al., 1995).
Amplificaion of CMV/CP coding sequence: Amplification was performed in thin-walled PCR tubes contained the reaction mixture as following: 5 μL of cDNA reaction, 45 μL of the amplification mixture containing 5 μL of 10X Taq DNA polymerase buffer (20 mM Tris-HC1, pH 8.2, 10 mM KCl, 6 mM Gelatin, 2 mM MgCl2, 0.1% Triton X-100 and 10 μg mL-1 of nuclease-free BSA), 1 mM each dNTP, 50 ng of each complementary reverse primer 5’-TTGGATCCTCAGACTGGGAG-3’ and forward primer 5’-AACCATGGACAAATCAGAA-3’ (Abdelkader et al., 2006), 2.5 unites of Taq polymerase (5 unites μL-1) (Promega) and completed with sterile distilled water. The amplification was carried out using the UNOII thermocycler system (Biometra, Germany) and using 0.2 mL micro. Amp. PCR tubes. Hard denaturation of the DNA was performed at 94°C for 3 min. followed by 35 cycles of amplification with denaturation at 94°C for 1 min annealing at 45°C for 2 min and extension 72°C for 1.5 min. A single trailing cycle of long extension at 72°C for 7 min ended the run. The PCR products were analyzed by electrophoresis onto 1% agarose gel in 1X TAE buffer (40 mM Tris-acetate, 1 mL acetic acid, 2 mM Na-EDTA and ethidium bromide [10 mg mL-1]).
DNA sequencing of CMV coat protein gene: DNA fragments were purified from agarose gel using the gel slicing and melting method described by Wieslander (1979). The nucleotide sequence of the cp for CMV was carried out at Macrogen 3730XL6-1518-009, Korea by ABI 1.6.0 sequencer. The sequence data was translated and analyzed by DNAMAN program.
RESULTS
Virus isolation: The virus isolate was detected in the naturally infected cucumber plants showing mosaic, net yellow and malformation while gave positive results by DAS-ELISA using specific CMV-IgG. It was biologically isolated by single local lesion from C. amaranticolor gave chlorotic local lesions surrounded by yellowing hallo and propagated in healthy N. glutinosa and cucumber plants as well as confirmed by DAS-ELISA.
| Table 1: | Reactions of host range plants inoculated with CMV* |
 | |
| NS: No symptoms; S: Severe; CLL: Chlorotic local lesions; M: Mosaic; B: Blisters; N: Necrosis; Mf: Malformation; VB: Vein banding; Vc: Vein clearing; FL: Fern leaf and D: Discoloration, NLL: Necrotic local lesions, SM: Severe mosaic, Y: Yellowing. *All CMV infected hosts were detected by DAS-ELISA and local lesion host | |
Virus identification: The virus isolate was identified as CMV on the basis of biological properties (external symptoms, host range, mode of transmission, virus stability, inclusion bodies and virus morphology), serological reactions (DAS-ELISA and DBIA) and molecular characters (RT-PCR and CP-sequence).
Host range: The reaction of eighteen plant species belonged to 6 families to the virus are summarized into three symptom types in Table 1. Local symptoms C. amaranticolor, C. metel and Gomphrena globosa produced chlorotic local lesions while C. murale produced necrotic local lesions on inoculated leaves after 4 days post inoculation. (Fig. 1; 4, 9, 10). Systemic symptoms, cucumber plants showed severe mosaic, blisters and malformation (Fig. 1; 1). While squash plants gave severe mosaic, green vein banding, blisters and malformation (Fig. 1; 2). N. glutinosa appeared severe mosaic, fern leaf and malformation; N. tabacum cv. White Burly produced severe mosaic; Datura metel showing severe mosaic and malformation, Helicrysum bracteatum showed yellowing symptoms and Petunia hybrida gave severe mosaic, malformation and discoloration (Fig. 1; 3, 5, 6, 7, 8). Symptomless, the following species did not react with CMV; Cucurbita maxima, N. rustica, Lycopersicon esculantum, Capsicum annum, Vigna sinensis and Vicia faba (Table 1).
Virus stability in crude sap: The stability of CMV isolate in infectious crude sap extracted from infected Cucumis sativus. It was determined by local lesions on leaves of C. amaranticolor as an indicator host. The virus isolate was inactivated at 70°C for 10 min., DEP was 10-4 and CMV kept its infectivity for 4 days at room temperature 25-28°C.
   | |
| Fig. 1(1-10): | Host plants mechanically inoculated with CMV isolate showing different types of symptoms on leaves: (1) Cucumis sativus, (2) Cucurbita pepo, (3) Helicrysum bracteatum, Healthy (H.) and infected (Inf.), (4) C. amaranticolor, (5-6) Development of symptoms on N. glutinosa, Healthy (H.) and infected (Inf.), (7) Petunia hybrida, Healthy (H.) and infected (Inf.), (8) Infected D. metel, (9) C. murale, Healthy (H.) and infected (Inf.) and (10) Infected G. globosa. |
Crystalline and amorphous inclusions: The crystalline inclusions induced by CMV were observed in epidermal and hair cells, as well as amorphous inclusions stained by bromophenol blue and mercuric chloride in leaves, 15 days post virus inoculation, Fig. 2.
The purified virus: The purification method of CMV isolate gave high yield of virus particles (1.43 mg/100 g fresh weight leaves). The UV spectrum of CMV particles was showed that, the absorption ratio of min., max. nm, Amax. /Amin., A260/A280 and A280/A260 were 248, 261, 1.101, 1.204 and 0.83, respectively.
Electron microscopy of purified CMV isolate was negatively stained with uranyl actate (2%), showed isometric particles with 30 nm in diameter.
Serological reaction: The virus antigen was serologically precipitated reaction against specific polyclonal IgG-CMV by immunoblotting, Fig. 3. A purplish blue color was developed with infected cucumber in the positive reaction, whereas extracts from healthy plants remain green in the negative reactions.
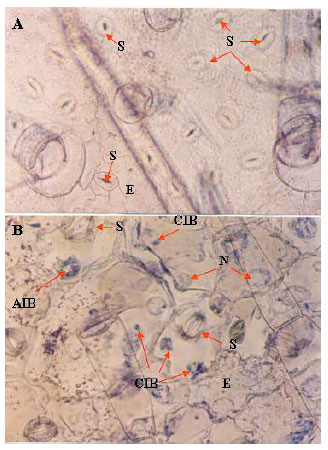 | |
| Fig. 2(A-B): | Light micrograph of epidermal strips and hairs (A) healthy and (B) CMV infected cucumber leaves (15 days post inoculation) showing cytoplasmic inclusion bodies. Magnification (X-400). N: Nucleus, S: Stomatic, E: Epiderms, CIB: Crystalline Inclusion Bodies and AIB: Amorphous Inclusion Bodies |
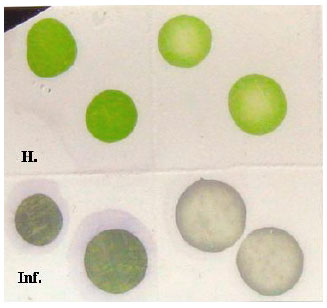 | |
| Fig. 3: | Dot Blot Immunoassay for CMV precipitation against specific IgG-CMV polyclonal. H: Healthy samples, Inf.: Infected samples |
Molecular characters of CMV isolate: The concentration of the RNA was 95 μg/0.2 g of infected tissues and the purity was measured by A260/280 absorbance ratio 1.5.
cDNA of CP-CMV was synthesized by MMLV reverse transcriptase. The resulting complementary DNA (cDNAs) was amplified by PCR after adding one set of primer CMV for cp gene. The efficiency of DNA amplification from CMV infected leaf tissues was detected by analysis PCR product using 1.0% agarose gel electrophoresis. The size of amplified CMV-DNA fragment was as expected 582 bp. (Lane 2) in Fig. 4.
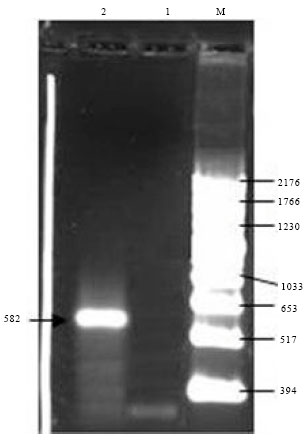 | |
| Fig. 4: | One % agarose gel electrophoresis showing the amplified PCR product of CMV/CP of the correct size (582 bp). Lane 1: Healthy N. glutinosa, Lane 2: amplified CMV/CP isolate, M: DNA marker |
Partial nucleotide sequence of CMV-cp gene: The partial nucleotide sequence (582 nucleotide) of the CMV-EG-CP was aligned with other coat protein sequences of CMV satellite RNA published in GeneBank as shown in Fig. 5 by using DNAMAN program (DNAMAN V 5.2.9 package, Madison, Wisconsin, USA).
A Phylogenetic tree of CMV-EG presented in Fig. 6 revealed the nucleotide sequence of CMV under study has 41% identified to AB024493 and D00542 satellite RNA CMV isolates.
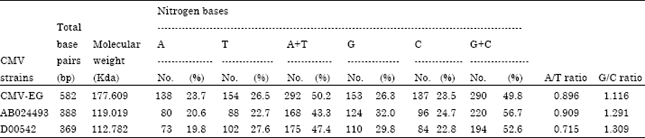
The nucleotide sequence of cp gene for CMV-EG (Table 2) revealed the highest content for Thymine (T) 154 (26.5%) followed by Guanine (G) 153 (26.3%), then Adenine (A) 138 (23.7%) and Cytosine 137 (23.5%).
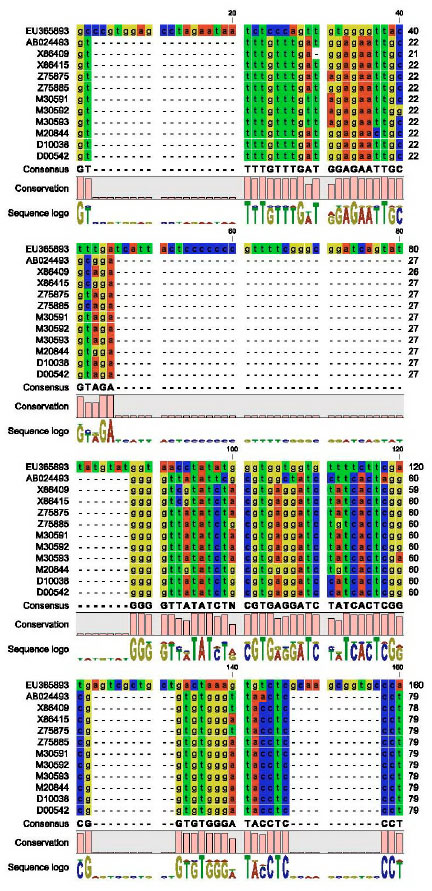 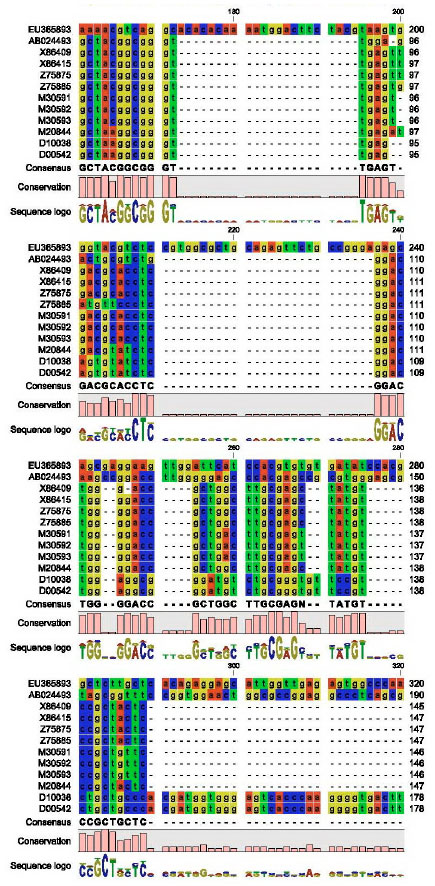  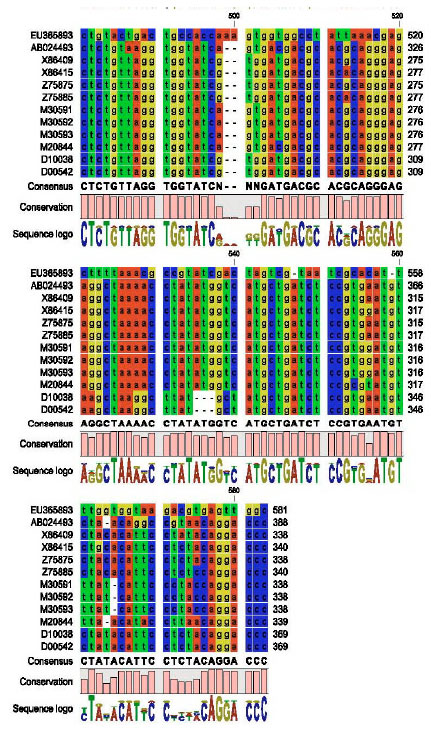 | |
| Fig. 5: | Partial nucleotide sequence alignment of the CP for CMV-EG using DNAMAN program |
Comparison between nitrogen bases composition of partial cp gene sequence for CMV-EG (Accession No. EU365893) and two CMV satellite RNA isolates published in GeneBank, AB024493 and D00542 (Hidaka et al., 1984) was done to determine A/T, G/C ratio as well as A+T and G+C percentage was 0.896, 1.116, 50.2 and 49.8% of CMV-EG, 0.909, 1.291, 43.3 and 56.7% of AB024493 and 0.715, 1.309, 47.4 and 52.6% to D00542, respectively.
Deduced amino acids sequence of CMV/cp gene: The predicted numbers of amino acids produced from translation of partial cp gene nucleotide sequence of CMV-EG were 193 amino acids starting with proline (P) and ended with leucine (L), in Fig. 7.
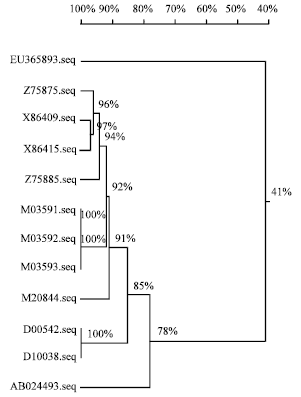 | |
| Fig. 6: | Consensus phylogenic tree constructed from the multiple alignment of the CP/CMV-EG (under study) and eleven satellite CMV isolates |
 | |
| Fig. 7: | Predicted amino acid of the cp gene of the isolated CMV-EG |
| Table 2: | Comparison between bases composition of partial CP sequence CMV and different two CMV satellite isolates published in GeneBank |
 | |
Comparison between the predicted amino acids of partial cp gene sequence of CMV-EG showed that, the type of amino acids consists of 20 amino acids. Leucine and serin has a high frequency and percentage 20 and 10.36%, respectively while tryptophan has a low frequency and percentage 1 and 0.51%.
DISCUSSION
In the present study, the CMV was detected in naturally infected cucumber plants cv. Barakoda which showed distincted viral symptoms in the form of mosaic, mottle, blisters, crinkle, net yellow and malformation. The detected samples gave +ve reaction against specific antiserum by using DAS-ELISA technique (Cardin and Moury, 2007).
CMV was isolated by single local lesion assay on C. amaranticolor as reported by many investigators (Osman et al., 2005; Awasthi et al., 2005; El-Afifi et al., 2007). N. glutinosa was used as a propagative host, where as revealed severe systemic symptoms in the form of severe mosaic, fern leaf and malformation. These symptoms were differed than common CMV which showing mosaic only. These results in agreement with Kim et al. (2011).
The CMV isolate under study was identified based on biological, serological and molecular characters. It infected 12 plant species and cultivars, 8 out of 12 plants showed systemic symptoms while only 4 species showed local lesions, Table 1. These results were in accordance with that of many investigators (Roossinck 2001; Osman et al., 2005; Hu and Chang, 2006).
Duffus (1973) stated that, disease symptoms on plants in the field are almost always inadequate on their own to give a positive identification. This is particularly so when several viruses cause similar symptoms. Matthews (1991) reported that, since, early days of plant virology, searches have been made for suitable species or varieties of host plant that will give clear, characteristic and consistent symptoms for the virus or viruses being studied usually under greenhouse conditions, such as indicator hosts provide one of the most basic tools for routine diagnosis. Many good indicator species have been found in the genera, Nicotiana, Solanum, Chenopodium, Cucumis, Phaseolus, Vicia and Brassica. In this investigation C. amaranticolor and C. murale were used as an indicator plants.
The virus isolate is sap transmissible. Its thermal inactivation point is 70°C, dilution end point 10-4 and the virus completely inactivated after 4 days at room temperature. These results were differed than CMV common which have 65°C, 10-3 and 4 days of TEP, DEP and LIV, respectively. These results were compatible with that found by Kiranmai et al. (1997) and Osman et al. (2005).
Matthews (1991) reported that, stability of the virus (thermal inactivation point, dilution end point and ageing at room temperature) as measured by infectivity (often in crude extracts) was an important criterion in attempting to establish groups of viruses.
The CMV-EG isolate formed cytoplasmic inclusions in infected epidermic cell leaves.
Virus purification was applied using different centrifugation which give 1.43 mg/100 g fresh weight leaves. The absorption ratio A260/A280 was 1.204. These results agreed with those reported by Hancheng et al. (1992).
Electron microscopy of purified CMV showed isometric particles shape with 30 nm in diameter, the same result was mentioned by El-Afifi et al. (2007) and Cardin and Moury (2007).
The results of the dot blot immunoassay technique was found to be sensitive to detect CMV in all infected plants in accordance with that obtained by Awasthi et al. (2005) and Zein et al. (2007). The antigenic determinants of CMV-EG isolate were identified by using polyclonal antibodies specific CMV which give serologically precipitation reacted in DAS-ELISA and in DBIA where the purple color appeared clearly.
RT-PCR of the CMV/CP-Egyptian isolate was used to amplify a fragment of about (582 bp) using primers according to Yu et al. (2005) and Abdelkader et al. (2006). Yu et al. (2005) mention that, the size of DNA fragment was 600 bp for CMV/CP, the results were expected as the same range of CMV/CP with the obtained results while (Wu et al., 2007) the result of the CMV-CP amplified from Capsicum annuum showed the band of the fragment with 740 bp. As well as, Akhtar et al. (2008) found that, amplified PCR product was 850 bp from the CMV infected tomato leaves.
The nucleotide sequence of coat protein gene of CMV was 890 nucleotides which revealed more than 90% similarity with the CMV subgroup I strains (Haq et al., 1996) while CMV-EG isolate under study revealed 582 nucleotides of CP-sequence with 41% identified to AB024493 and D00542 satellite RNA CMV isolates.
The nucleotide sequence of cp gene for CMV-EG revealed that the highest content for Thymine (T) 154 (26.5%) followed by Guanine (G) 153 (26.3%), then Adenine (A) 138 (23.7%) and Cytosine 137 (23.5%).
Comparison between nitrogen bases composition of partial cp gene sequence for CMV-EG (Accession No. EU365893) and two CMV satellite RNA isolates published in GeneBank, AB024493 and D00542 (Hidaka et al., 1984) was done to determine A/T, G/C ratio as well as A+T and G+C percentage were 0.896, 1.116, 50.2 and 49.8% of CMV-EG., 0.909, 1.291, 43.3 and 56.7% of AB024493 and 0.715, 1.309, 47.4 and 52.6% to D00542, respectively.
Comparison between the predicted amino acids of partial cp gene sequence of CMV-EG has showed that, the type of amino acids consists of 20 amino acids. Leucine and serin have a high frequency and percentage 20 and 10.36%, respectively while tryptophan has a low frequency and percentage 1 and 0.51%.
As the previous results, it was found that, the obtained CMV-EG isolate under study was differed from other common CMV isolates on biological characters symptomatology on cucumber, N. glutinosa and C. amaranticolor. On other hands, the obtained nucleotide sequence of CMV-EG had a similarity (41%) with different eleven satellite RNA Cucumber mosaic virus which published in GeneBank, so we suggest that, the obtained isolate is a satellite CMV and its nucleotide sequence was published in GeneBank with Accession No. EU365893 as a new Egyptian satellite strain of Cucumber mosaic cucumovirus.
REFERENCES
- Akhtar, K.P., K.H. Ryu, M.Y. Saleem, M. Asghar, F.F. Jamil, M.A. Haq and I.A. Khan, 2008. Occurrence of Cucumber mosaic virus subgroup IA in tomato in Pakistan. J. Plant Dis. Prot., 115: 2-3.
Direct Link - Awasthi, L.P., K. Pardeep and K.M. Nehal, 2005. Management of Cucumber mosaic virus disease in cucumber through root extract of Boerhaavia diffusa. Ann. Plant Prot. Sci., 13: 256-257.
Direct Link - Brigneti, G., O. Voinnet, W.X. Li, L.H. Ji, S.W. Ding and D.C. Baulcombe, 1998. Viral pathogenicity determinants are suppressors of transgene silencing in Nicotiana benthamiana. EMBO J., 17: 6739-6746.
CrossRefDirect Link - Cardin, L. and B. Moury, 2007. First report of Cucumber mosaic virus in Echium candicans in France. Plant Dis., 91: 1516-1516.
CrossRef - Clark, M.F. and A.N. Adams, 1977. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol., 34: 475-483.
CrossRefPubMedDirect Link - El-Afifi, S.I., A.M. El-Borollosy and S.Y.M. Mahmoud, 2007. Tobacco callus culture as a propagating medium for cucumber mosaic Cucumovirus. Int. J. Virol., 3: 73-79.
CrossRefDirect Link - Gibbs, A. and A. Mackenzie, 1997. A primer pair for amplifying part of the genome of all potyvirids by RT-PCR. J. Virol. Methods, 63: 9-16.
PubMedDirect Link - Haq, Q.M.R., B.P. Singh and K.M. Srivastava, 1996. Biological, serological and molecular characterization of a cucumber mosaic virus isolate from India. Plant Pathol., 45: 823-828.
CrossRef - Hidaka, S., K. Hanada, K. Ishikawa and K. Miura, 1988. Complete nucleotide sequence of two new satellite RNAs associated with cucumber mosaic virus. Virology, 164: 326-333.
PubMed - Hidaka, S., K. Ishikawa, Y. Takanami, S. Kubo and K. Miura, 1984. Complete nucleotide sequence of RNA 5 from cucumber mosaic virus (strain Y). FEBS Lett., 174: 38-42.
CrossRef - Hu, W.C. and Y.C. Chang, 2006. Cucumber mosaic virus in New Guinea impatiens in Taiwan. Acta Hortic., 722: 241-246.
Direct Link - Kaperi, J.M., M.E. Tousignant and M.T. Steen, 1988. Cucumber mosaic virus-associated RNA 5 XI. Comparison of 14 CARNA 5 variants relates ability to induce tomato necrosis to a conserved nucleotide sequence. Virology, 163: 284-292.
CrossRef - Kaplan, I.B., M.H. Shintaku, Q. Li, L. Zhang, L.E. Marsh and P. Palukaitis, 1995. Complementation of virus movement in transgenic tobacco expressing the cucumber mosaic virus 3a gene. Virology, 209: 188-199.
CrossRefPubMedDirect Link - Kim, M.K., H.R. Kwak, S.H. Lee, J.S. Kim, K.H. Kim, B.J. Cha and H.S. Choi, 2011. Characteristics of Cucumber mosaic virus isolated from Zea mays in Korea. Plant Pathol. J., 27: 372-377.
Direct Link - Lin, N.S., Y.H. Hsu and H.T. Hsu, 1990. Immunological detection of plant viruses and a mycoplasmalike organism by direct tissue blotting on nitrocellulose membranes. Phytopathology, 80: 824-828.
CrossRefDirect Link - Mazia, D., P.A. Brewer and M. Alfert, 1953. The cytochemical staining and measurement of protein with mercuric bromphenol blue. Biol. Bull., 104: 57-67.
CrossRefDirect Link - Osman, M., K. El-Dougdoug, E.T. Abd El-Salam, R.M. Taha and R.M. El-Hamid, 2005. Histo-cytopathological effects of cucumber mosaic cucumovirus on squash leaves. Int. J. Virol., 1: 34-34.
CrossRefDirect Link - Roossinck, M.J., 2001. Cucumber mosaic virus, a model for RNA virus evolution. Mol. Plant Pathol., 2: 59-63.
CrossRef - Schwinghamer, M.W. and R.H. Symons, 1977. Translation of the four major RNA species of cucumber mosaic virus in plant and animal cell-free systems and in toad oocytes. Virology, 79: 88-108.
CrossRef - Sharma, A., B.K. Mahinghara, A.K. Singh, S. Kulshrestha and G. Raikhy et al., 2005. Identification, detection and frequency of lily viruses in Northern India. Sci. Horticult., 106: 213-227.
CrossRef - Tobias, I., D.Z. Maat and H. Huttinga, 1982. Two Hungarian isolates of cucumber mosaic virus from sweet pepper (Capsicum annuum) and melon (Cucumis melo): Identification and antiserum preparation. Eur. J. Plant Pathol., 88: 171-183.
CrossRef - Valverde, R.A. and J.A. Dodds, 1987. Some properties of isometric virus particles which contain the satellite RNA of tobacco mosaic virus. J. Gen. Virol., 68: 965-972.
CrossRefDirect Link - Wieslander, L., 1979. A simple method to recover intact high molecular weight RNA and DNA after electrophoretic separation in low gelling temperature agarose gels. Anal. Biochem., 98: 305-309.
CrossRefDirect Link - Zein, H.S., M. Nakazawa, M. Ueda and K. Miyatake, 2007. Development of serological procedures for rapid and reliable detection of Cucumber mosaic virus with dot-immunobinding assay. World J. Agric. Sci., 3: 430-439.
Direct Link - Hancheng, Z., L. Huanting and Y. Dunyu, 1992. Identification of Cucumber mosaic virus from ambary plant. J. Shandong Agric. Univ., 23: 165-169.
Direct Link