
Kh.A. El-Dougdoug
Department of Microbiology, Faculty of Agriculture, Ain Shams University, Egypt
M.F. Ghaly
Department of Botany, Faculty of Science, Zagazig University, Egypt
M.A. Taha
Department of Botany, Faculty of Science, Zagazig University, Egypt
International Journal of Virology
Year: 2012 | Volume: 8 | Issue: 2 | Page No.: 151-164







ABSTRACT
An antiviral producing Streptomyces species were isolated from soil rhizosphere in Zagazig province of Egypt. In order to identify the Streptomyces strains, morphological, physiological, biochemical and antagonism tests were performed. The Egyptian isolates of Streptomyces were found to be a species of calvus, canarius, vinaceusdrappus, nogalater and viridosporus. The Streptomyces spp. were grown in glycerol asparagine broth medium and the culture supernatants obtained were 0.45-μm filter. These isolates were tested in two experiments for their ability to control a Cucumber Mosaic Virus (CMV). In the 1st experiment, one half of leaves of Chenopodium amaranticolor were treated with Culture Filtrate (CF) followed by CMV inoculation on both halves. In the 2nd experiment, the first pair of Cucumis sativus leaves were treated CF with CMV mechanically inoculated onto one leaf, the other non-treated leaf was CMV inoculated after 7 days of treatment. In 1st experiment, CF treatment was able to considerably reduced the number of local lesion and in 2nd experiment, plants treated with CF showed variable visible viral symptoms compared with the broth media treated control 15 days post inoculation and remained symptom less throughout the study period. Such five Streptomyces species identified were able to produce an antiviral component in the culture filtrate, non phytotoxic and effective in local as well as systematically control of CMV infection.
PDF Abstract XML References Citation
Received: September 20, 2011;
Accepted: November 02, 2011;
Published: December 02, 2011
How to cite this article
Kh.A. El-Dougdoug, M.F. Ghaly and M.A. Taha, 2012. Biological Control of Cucumber Mosaic Virus by Certain Local Streptomyces Isolates: Inhibitory Effects of Selected Five Egyptian Isolates. International Journal of Virology, 8: 151-164.
DOI: 10.3923/ijv.2012.151.164
URL: https://scialert.net/abstract/?doi=ijv.2012.151.164
DOI: 10.3923/ijv.2012.151.164
URL: https://scialert.net/abstract/?doi=ijv.2012.151.164
INTRODUCTION
Cucumber Mosaic Virus (CMV), belonging to the genus Cucumovirus, family bromoviridae is one of the economically important viruses which causes enormous losses by infecting more than 1,000 species of plants, shrubs and trees world-wide. It is transmitted non-persistently into healthy plants by aphids which acquire the virus during their brief probes on infected hosts or the symptom less carrier weeds in the field (Zehnder et al., 2000). Various strategies based on the avoidance of sources of infection, control of vectors, modification of cultural practices, use of resistant varieties and transgenic plants have been conventionally employed to minimize the losses caused by CMV. These strategies; however, have not been effective as control measures. Many screening studies have been conducted on antiviral agents from different sources. Most of these come from plants sources with some showing systemic control ability against range of viruses that infect plants (Kubo et al., 1990). Comparatively, antiviral from microbial sources have been little studied. Recently, Raupach et al. (1996) showed the systemic control of CMV in cucumbers and tomatoes employing rhizosphere colorization of some bacteria by an induced systemic infection mechanism. Kim et al. (2004) used culture filtrate from Acinetobacter sp. KTB3 to systematically control some viruses in Korea.
This investigation was primarily concerned with the identification of five Streptomycetal isolates based on cultural growth, morphological, physiological, biochemical and tolerance of salinity, melanin pigment and antibiosis methods. Also, study the effect of heat stable culture filtrate of some Streptomyces spp. isolates as antiviral substance against CMV which produce local lesions in the hypersensitive host and systematically infects many important plants.
MATERIALS AND METHODS
Collection of soil samples: Rhizosphere soil samples (silty clay soil) under cultivated different crops were collected from different locations Zagazig, Hehia, Abu-Kabir and Fakous belong to El-Sharkia Governorate according to the procedures described by Johnson et al. (1960). In this method, soil samples were collected by sterilized hand corer at depth of 15 cm from different regions in clean plastic bags. The collected samples were transfer to the laboratory and kept in refrigerator till used.
Isolation and purification of Streptomyces spp.: The collected soil samples were air-dried, ground in a mortar and then mixed with calcium carbonate (CaCO3) and followed by sieving in 4 mm mesh screen. One gram of prepared soil was stirred in 100 mL sterile distilled water for about 5 min in a 250 mL Erlemeyer flask and the suspension allowed to stand for 30 min. Serial dilution (10-1, 10-5) of each obtained suspension were prepared in sterile saline solution (0.85% NaCl) and 1 mL of each dilution was spread on Petri-dish containing 15 mL of starch nitrate agar medium. The dishes were rotated by hand to insure homogenous distribution of soil suspension dilution and then incubated for 7 days at 28°C till Streptomyces colonies appearance.
The purification was achieved according to Kuester and Williams (1964) by picking up of unique single identical morphological Streptomyces colony based on cultural morphological characters and repeated streaking on starch nitrate agar plates. Each of these was assayed for antiviral activity using the half-leaf method as described by Kubo et al. (1990).
Five selected Streptomycetal isolates out of 30 actinomycetal isolates were used for identification and comparative study of the physiological, morphological, biochemical, tolerance of salinity, melanin pigment and antibiosis analyses.
Morphological and physiological tests: In order to identify the Streptomycetal isolates 48 physiological and biochemical tests were carried out. These tests included growth in different cultural media such as starch nitrate agar, glycerol nitrate agar, inorganic starch agar, Nutrient agar, glucose asparagines agar, yeast malt agar, glucose nitrate agar, glycerol asparagines agar, sucrose nitrate agar and oat meal. Morphological characters, presence or absence of sporangium, length of sporangiophore, spore mass, spore surface, spore chain, spore shape. Physiological characters, melanin pigment, proteolytic activity, lipolytic activity, lethicine activity, cellulolytic activity, gelatin liquefaction, growth in different carbon sources, growth at different temperature, tolerance to NaCl and antibiosis were determined.
Maintenance of virus: CMV was kindly obtained from Virology Lab. Microbiology Dept., Fac. of Agric., Ain Shams University and maintained in N. glutinosa as CMV-propagation host. The inoculum of CMV was prepared from systematically infected N. glutinosa leaves ground in 0.1 M phosphate buffer, pH 7.2.
Antiviral bioassay: The Streptomycetal species was grown in glycerol asparagines broth medium and the culture supernatants obtained were filtered through 0.45 l litter. The antiviral activity of the CF from five selected Streptomyces spp. was estimated in two experiments. In the first experiment, CF was assayed on a hypersensitive host for CMV, i.e., C. amaranticolor using the half leaf method, as previously described by Kubo et al. (1990). The upper right halves of the leaves were treated with CF using paintbrush and the upper left halves were treated with sterilized water as a control treatment. After one hour, the virus was inoculated onto both halves of the leaves. The experiment was performed in duplicate. The plants were kept in a greenhouse, at 12-14 h daylight and a temperature of 30°C. The number of local lesions were counted after seven days post inoculation. The inhibitory effect was calculated according to the formula: I = (1-T/C)x100, where, T the number of local lesions on the treated half of the leaves and C is the number of local lesions on the control half of the leaves. In the second experiment, the 1st pair of cucumber (Cucumis sativus) cv Barakoda leaves were used under each treatment. One leaf was treated with CF, the other non-treated leaf was CMV inoculated after 7 days of treatment. As a control treatment, one leaf was treated with broth media, another untreated leaf was CMV inoculated after 7 days. Each experiment was replicated three times.
RESULTS
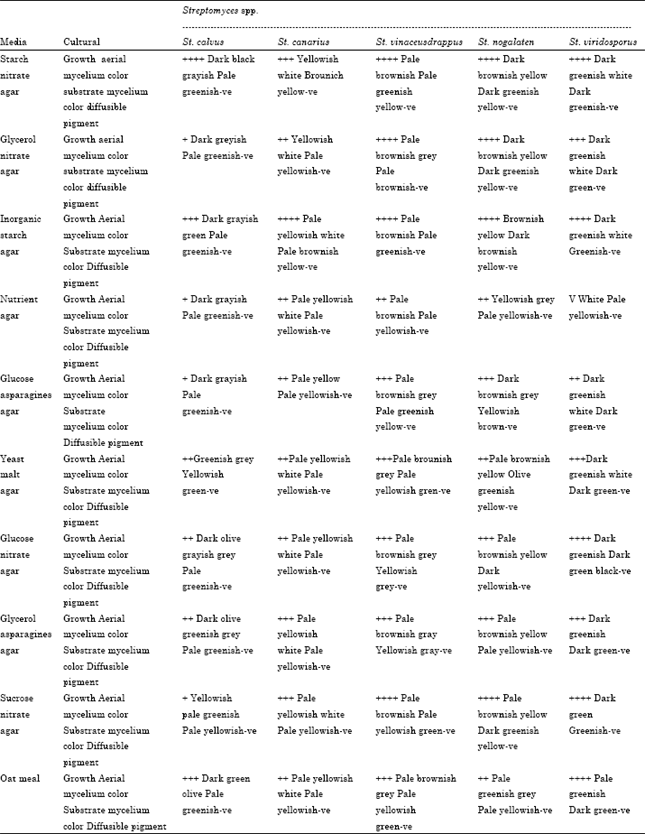
Identification of Streptomycetal isolates: The five Streptomyces spp. were isolated from soil rhizosphere according to variation in growth rate on differential media, morphological and biochemical tests. Five isolates were appeared variation in growth rate on differential media (Table 1). St. viridosporus was revealed variable growth on nutrient agar medium while St. calvus revealed weak rate growth on glycerol nitrate agar, nutrient agar, glucose asparagines agar and sucrose nitrate agar medium compared with other isolates. On the other hand, St. vinaceusdrappus and St. nogalater were revealed strong growth rate on experimental media (Table 1). All five isolates showed diffusible pigment in all experimental media under study. Visual observations by light and electron micrographs (Fig. 2) of the five isolates showed that no sporangium formation and different in sporangiophore where as, 6 μm, 4.5, 7.5, 4 and 3 for St. calvus, St. canarius, St. vinaceus-drappus, St. nogalater and St. viridosporus, respectively (Fig. 1). In related to the spore chain were different in among five isolates, spiral short, spiral long, spiral open long, spiral open long and spiral long with St. calvus, St. canarius, St. vinaceus-drappus, St. nogalater and St. viridosporus, respectively (Fig. 1). In addition, the spore mass also differed between five isolates such as dark grayish, yellowish white, pale brownish, yellow brownish and green for St. calvus, St. canarius, St. vinaceusdrappus, St. nogalater and St. viridosporus, respectively. The spore surface of St. calvus was hairy, St. viridosporus was spiny while St. canarius, St. vinaceusdrappus and St. nogalater were smooth.
Conidiospore morphology were differed among 5 isolates whereas St. calvus and St. viridosporus were revealed oval shape with diameter 12x17 and 10x13 mm, respectively while, St. canarius, St. vinaceusdrappus and St. nogalater were revealed barrel shape with 7x20, 9x11 and 8x10 mm, respectively (Table 2).
| Table 1: | Cultural characteristics of Streptomyces spp. |
 | |
| Growth: ++++: Very strong growth, +++: Strong growth, ++: Moderate growth, +: Weak strong, -ve: negative, +ve: Positive, V: Variable growth | |
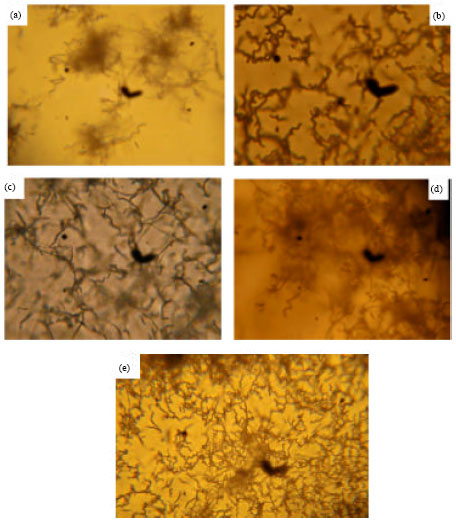 | |
| Fig. 1(a-e): | Light micrograph of spore chains for Streptomyces spp. (X 200); (a) St. calvus, (b) St. canaries, (c) St. vinaceusdrappus, (d) St. nogalater and (e) St. viridosporus |
| Table 2: | Morphological characteristics of Streptomyces spp. |
 | |
| Magnification 10,000 X, -: Absence of sporangium | |
Data represented in Table 3 recorded that St. canarius and St. viridosporus were able to secrete melanin pigment on tyrosine agar medium while St. calvus, St. vinaceus-drappus and St. nogalater not able. On the other hand five isolates were not able to secrete on tryptone broth and peptone yeast iron agar media.
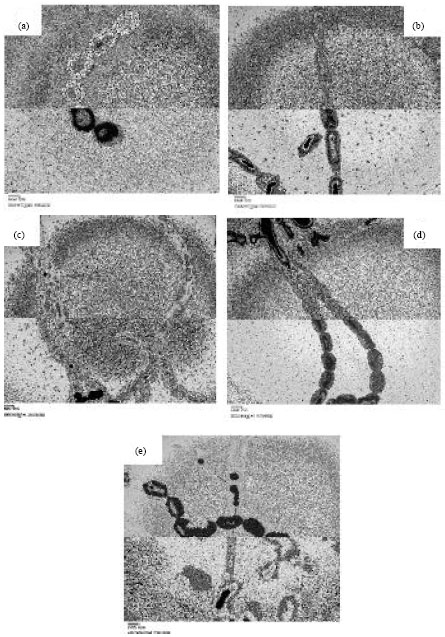 | |
| Fig. 2(a-e): | Electron micrograph of spore surface for Streptomycetal isolates (X 15000); (a) St. calvus, (b) St. canarius, (c) St. vinaceusdrappus, (d) St. nogalater and (e) St. viridosporus |
Data also showed that 5 isolates have proteolytic and amylolytic activity while only St. viridosporus, had cellulytic activity. On other hand, St. calvus, St. vinaceusdrappus and St. nogalater have pectinolytic activity while St. canarius and St. viridosporus not have. The result showed that only St. canarius able to lethicine degradation while the others not able. On the other hand 5 isolates able to make gelatin liquefaction and H2S production as well as St. vinaceus-drappus and St. nogalater can reduce nitrate but other isolates can't reduce (Table 3).
Data showed that St. calvus, St. vinaceusdrappus, St. nogalater and St. viridosporus gave variable growth on medium without carbon source while St. canarius can't grows.
| Table 3: | Physiological characteristics of Streptomyces spp. |
 | |
| ++++ : Very strong growth, +++ : Strong growth, ++ : Moderate growth, + : Weak growth, v : Variable growth, -: No growth | |
On other hand 5 isolates gave different growth rates on media with different carbon sources except St. calvus can't grow in medium with maltose as carbon source.
All 5 isolates showed antimicrobial potentialities against tested organisms except St. vinaceusdrappus, St. nogalater and St. viridosporus not showed against E. coli. On the other hand, Helminthosporium solani appeared the most sensitive one for 5 isolates followed by Fusarium sp. followed by Staph. aureus and finally E. coli. The data showed that fungal isolates were more sensitive to 5 isolates than bacterial isolates. On the other hand, St. canaries showed the higher antimicrobial potentialities against tested organisms due to increasing in the diameter of inhibition zone (Table 4).
| Table 4: | Antimicrobial activity of Streptomyces spp. |
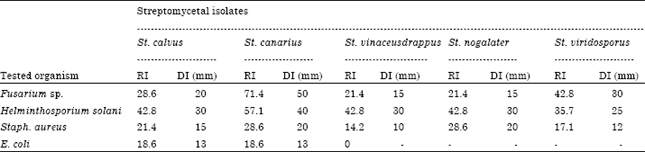 | |
| RI : Relative inhibition, DI: Diameter of inhibition zone | |
| Table 5: | Single local lesion variability of CMV-inoculated plants treated with Streptomyces isolates |
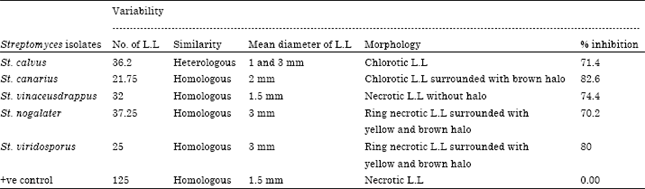 | |
Antiviral effects of streptomycetal isolates: The antiviral culture filtrate from five selected Streptomycetal isolates showed high inhibitory activity against CMV. The CF treated part of the hypersensitive host; C. amaranticolor leaves showed 70.2, 71.4, 74.4, 80 and 82.6% inhibition of the production of local lesions compared to the untreated part of the leaves for Streptomyces calvus, Streptomyces canarius, Strepotomyces vinaceusdrappus, Streptomyces nogalater and Streptomyces viridosporus, respectively. The control plants, treated with sterilized water were unable to show inhibition of CMV induced lesions (Table 5). The average number of local lesions in the case of the CF treated half leaves were much lower 36.2, 21.75, 32, 37,25 and 25, respectively than those of the sterilized water treated half leaves (125).
The five Streptomycetal isolates appeared CMV variability based on the variation of single L.L. St. calvus induced heterologous chlorotic L.L, 1 and 3 mm in diameter with reduction percentage 71.4%, Fig. 3-St. 1. St. canarius induced homologous chlorotic L.L. surrounding with brown halo, 2 mm in diameter with reduction percentage 82.6%, St. 10 (Fig. 3). St. vinaceusdrappus produced homologous necrotic L.L, without halo, 1.5 mm in diameter with reduction percentage 74.4%, Fig. 3-St. 19. St. nogalater induced ring necrotic L.L surrounded with yellow and brown halo, 3 mm in diameter with reduction percentage 70.2%, Fig. 3-St. 28. St. viridosporus produced ring necrotic L.L surrounded with yellow and brown halo, 3 mm in diameter with reduction percentage 80%, Fig. 3-St. 29 and the positive control produced necrotic L.L, 1.5 mm in a diameter. The obtained previous screening Streptomycetal isolates results for induction systemic acquired resistance in cucumber plants against CMV infection revealed that, the most effective individual isolates were five Streptomyces elicitors, Streptomyces calvus, Streptomyces canarius, Streptomyces vinaceusdrappus, Streptomyces nogalater and Streptomyces viridosporus with No. 1, 10, 19, 28 and 29, respectively. These isolates were completely identified and used for next further studies.
| Table 6: | Effect of individual inducers on CMV infectivity in cucumber |
 | |
 | |
| Fig. 3: | Variability of local lesions resulted from CMV infected cucumber plant treated with 5 Streptomycetal isolates using C. amaranticolor as CMV indicator host. *All photo numbers point to numbers of different isolated Streptomyces as used in this work |
Five Streptomyces inducers out of 30 actinomycetal isolates (1) St. calvus (10) St. canarius (19) St. vinaceusdrappus (28) St. nogalater and (29) St. viridosporus out of twenty three Streptomycetal isolates were used for induction Systemic Acquired Resistance (SAR) in cucumber plants against CMV infection. The induced systemic resistance was detected by different methods, biologically (percentage of infection, Disease Severity (DS), virus variability). The five Streptomyces treatments have different percentage of DS, St. nogalater has a low percentage of DS (4.4%), while St. canarius has a high percentage of DS (22.2%).
The other isolates St. calvus, St. vinaceusdrappus and St. viridosporus have DS percentage 10, 20, 10%, respectively. Individual Streptomyces treatments in Table 6 showed that, the selected five Streptomyces isolates reduced the percentage of CMV infection as follow: 71.4, 82.6, 74.4, 70.2 and 80% for (St 1) (St 10) (St 19) (St 28) and (St 29), respectively. The highest percentage of reduction was 82.6% for St 10, while St 28 isolate has the lowest percentage of reduction 70.2%.
The obtained previous results showed that, the filtrate spraying of Streptomyces induced SAR in cucumber plants which played the major role to reduced CMV infection. On other hands showed significant variations in symptoms severity and vegetative growth of CMV infected cucumber plants (Fig. 4, 5) compared with untreated CMV infected Cucumber plants as infected control (Fig. 4, 5).
The CMV symptoms on treated cucumber plants after CMV inoculation were differed such as; vein clearing, mosaic, malformation and blisters (Fig. 4, 5).
 | |
| Fig. 4: | Illustrate the cucumber plant treated with different filtrate of Streptomycetal isolates. St 1: St. calvus, St 10: St. canarius, St 19: St. vinaceusdrappus, St 28: St. nogalater, St29: St. viridosporus, IC: Infected control |
 | |
| Fig. 5: | Development of different CMV symptoms in cucumber plants treated with different filtrate of Streptomycetal isolates. St 1 and St 10: vein clearing. St 19, St 28 and St 29: Mosaic, malformation and blisters. IC: infected control |
DISCUSSION
An antiviral producing streptomycetal species were isolated from soil rhizosphere in Zagazig province of Egypt. In order to identify the Streptomycetal strains, morphological, physiological, biochemical and antagonism testes were performed. The Streptomycetal were found to be a species of Streptomyces calvus, Streptomyces canarius, Strepotomyces vinaceusdrappus, Streptomyces nogalater and Streptomyces viridosporus which were designated as Egyptian isolates. The collected actinomycete isolates were subjected for a process of purification using the specific nutrient growth medium of starch-nitrate agar. The purification of the actinomycete isolates was conducted by means of different purification media which included starch nitrate agar and starch-inorganic media (Abo-Elanin, 2004). Many authors reported that, the isolation of 110 isolates of actinomycete cultures from eight soil samples was carried out using the conventional dilution plate method on Humic acid-vitamin agar, starch casein agar and sorenson's agar (Tan et al., 2001).
The purified actinomycete isolates were subjected to screening against program of antimicrobial activities. Most of isolates exhibited antimicrobial activities against Gram-positive, Gram-negative, acid-fast bacteria, yeasts and filamentous fungi (El-Abyad et al., 1996). Actinomycetal isolates were tested for their antagonistic potentialities according to diffusion (Cork borer) method (Betina, 1983). All 5 isolates showed antimicrobial potentialities against tested organisms except St. vinaceusdrappus, St. nogalater and St. viridosporus not showed against E. coli. On the other hand, Helminthosporium solani appeared the most sensitive one for 5 isolates followed by Fusarium sp. followed by Staph. aureus and finally E. coli. The data showed that fungal isolates were more sensitive to 5 isolates than bacterial isolates. On the other hand, St. canarius showed the higher antimicrobial potentialities against tested organisms due to increasing in the relative inhibition. This percentage is agreed with those described by many authors studying the activity of soil actinomycetes (Saadoun and Al-Momani, 1997; Saadoun et al., 1998; Ndonde and Semu, 2000). The highest antimicrobial streptomycetal isolates selected and inoculated in various liquid nutrient media for investigating its antiviral activity against CMV. Concerning of identification of the most active Streptomycetal isolates that have antiviral activities. The morphological and physiological properties of the actinomycete isolates No. 10 are consistent with assignment of Streptomyces canarius. This reveals that the antiviral activity of CF from Streptomycetal isolates were due to involvement of plant defense mechanism. In both the above experiments, no damage to the host plant was observed due to CF treatment. CF; thus, can be characterized as a non-toxic antiviral agent which would give the necessary efficiency in combating CMV. The activity of the inhibitory agent present in the CF obtain from Streptomycetal isolates was non-toxic and induces protection against CMV in both local as well as systemic hosts. Cucumber mosaic cucumovirus (S. CMV. EG) was obtained from Virology Lab, Agric Microbiological Dept., Fac. of Agric., Ain Shams University. Data indicates that the translocation of the antiviral effect from the CF treated half-leaf to the untreated part of the same leaf. When CF was used to elucidate the systemic control effect of Streptomycetal isolates, it was found that the plants treated with CF showed no visible viral symptoms 25 dpi (days post inoculation) and remained symptom less throughout the study period. The plants treated with broth media showed symptom less. CMV was confirmed by single local lesion assay of C. amaranticolor as reported by many investigators (Polak, 1999; El-Baz, 2004; El-Afifi et al., 2007; Megahed, 2008). CMV was transmitted mechanically to healthy susceptible test plants (Polak, 1999; Paradies et al., 2000; El-Baz, 2004; Awasthi et al., 2005; Hu and Chang, 2006; El-Afifi et al., 2007; Megahed, 2008). Accordingly, anti-infective activity may induce one or more of the following activities:
| • | Direct inactivation virus by the extract without affecting cell receptors or intracellular targets i.e., virucidal effect. This may be achieved by blocking the virus receptor molecules or by virolysis if the inactivant has an enzymatic activity or physically by antagonizing the net electric charge that lead to virus attraction to the host cells or by increasing the size of the virion and preventation of the fitting into the receptor |
| • | Induction of changes in cell membrane or cellular receptor of the virus |
| • | Preventation the virus adsorption and/or uncoating. The last two activities are related to the protective function of a drug |
The protective activity of an extract may include one or more of the following activities:
| • | Induction changes at the cell membrane leading to inhibition of the virus adsorption and/or penetration | |
| • | Induction of changes in the cell lysosomes inhibiting virus uncoating | |
| • | Setting the intracellular biochemical mechanisms in such a way which resists the virus replication as in case of interferon (Dimmock and Primrose, 1994). Finally, the anti-replicative activity may include one or more of the following activities: | |
| • | Inhibition of virus uncoating or Inhibition of cellular and/or viral translation mechanisms | |
| • | Inhibition of cellular and/or viral transcription and replication function | |
| • | Inhibition of viral protein processing and/or Capsid assembly or maturation | |
REFERENCES
- Awasthi, L.P., K. Pardeep and K.M. Nehal, 2005. Management of Cucumber mosaic virus disease in cucumber through root extract of Boerhaavia diffusa. Ann. Plant Prot. Sci., 13: 256-257.
Direct Link - El-Abyad, M.S., M.A. El-Sayed, A.R. El-Shanshoury and N.H. El-Batanony, 1996. Effect of culture conditions on the antimicrobial activities of UV-mutants of Streptomyces corchorusii and S. spiroverticillatus against bean and banana wilt pathogens. Microbiol. Res., 151: 201-211.
CrossRefDirect Link - El-Afifi, S.I., A.M. El-Borollosy and S.Y.M. Mahmoud, 2007. Tobacco callus culture as a propagating medium for cucumber mosaic Cucumovirus. Int. J. Virol., 3: 73-79.
CrossRefDirect Link - Hu, W.C. and Y.C. Chang, 2006. Cucumber mosaic virus in New Guinea impatiens in Taiwan. Acta Hortic., 722: 241-246.
Direct Link - Kim, Y.S., E.I. Hwang, O. Jeong-Hun, K.S. Kim, M.H. Ryu and W.H. Yeo, 2004. Inhibitory effects of Acinetobacter sp. KTB3 on infection of Tobacco mosaic virus in tobacco plants. Plant Pathol. J., 20: 293-296.
Direct Link - Kuester, E. and S.T. Williams, 1964. Production of hydrogen sulfide by streptomycetes and methods for its detection. Applied Microbiol., 12: 46-52.
PubMed - Ndonde, M.J.M. and E. Semu, 2000. Preliminary characterization of some Streptomyces species from four Tanzanian soils and their antimicrobial potential against selected plant and animal pathogenic bacteria. World J. Microbiol. Biotechnol., 16: 595-599.
CrossRef - Raupach, G.S., L. Liu, J.F. Murphy, S. Tuzun and J.W. Kloepper, 1996. Induced systemic resistance in cucumber and tomato against Cucumber mosaic cucmovirus using plant growth-promoting rhizobacteria (PGPR). Plant Dis., 80: 891-894.
Direct Link - Saadoun, I. and F. Al-Momani, 1997. Studies on soil streptomycetes from Jordan. Actinomycetes, 8: 42-48.
Direct Link - Saadoun, I., M.J. Mohammad, F. Al-Momani and M. Meqdam, 1998. Diversity of soil streptomycetes in Northern Jordan. Actinomycetes, 9: 52-60.
Direct Link - Zehnder, G.W., C. Yao, J.F. Murphy, E.R. Sikora and J.W. Kloepper, 2000. Induction of resistance in tomato against Cucumber mosaic cucumovirus by plant growth-promoting rhizobacteria. BioControl, 45: 127-137.
Direct Link