
A.R. Sofy
Department of Botany and Microbiology, Faculty of Science, Al-Azhar University, 11884 Nasr City, Cairo, Egypt
A.M. Soliman
Plant Virus and Phytoplasma Research Section, Plant Pathology Institute, Agriculture Research Center, Giza, Egypt
A.A. Mousa
Department of Botany and Microbiology, Faculty of Science, Al-Azhar University, 11884 Nasr City, Cairo, Egypt
Kh.A. El-Dougdoug
Department of Microbiology, Faculty of Agriculture, Virology Laboratory Agricultural, Ain Shams University, 11241 Cairo, Egypt
International Journal of Virology
Year: 2012 | Volume: 8 | Issue: 2 | Page No.: 133-150







ABSTRACT
This study reports the characterization of Hop stunt viroid which has been isolated previously in Egypt from sweet orange infected with gummy bark disease namely citrus viroid II. The native structure of mature, circular forms of the gummy bark agent was detected by gel electrophoresis. The expected size of amplified cDNA by RT-PCR was approximately 300 bp. A phylogenetic tree of the Egyptian citrus gummy bark agent (Accession no. FJ984562) revealed 100, 99.7 and 97.8% a moderate degree of similarity to CVd-IIb (USA), CCaVd (Spain) and CCaVd (Egypt), respectively. The minimum free energy of a secondary structure for HSVd-EG-RNA was determined using its primary sequence at 37°C. The sequence appeared to fold into a rod-like structure at -122.1 kcal moL-1 while CCaVd-EG at -120.5 kcal moL-1. The five domains of the rod-like structure were determined. The sequence variations between Egyptian citrus gummy bark isolate and Egyptian citrus cachexia isolate in the pathogenic domain (P) tend to influence the pathogenicity of the HSVd-EG. Finally, the genetic diversity and evaluation of entropy power for the Egyptian citrus gummy bark agent and HSVd-citrus populations registered in GenBank, were viewed against the phylogenetic background of known CVd-II variants including the non-cachexia (CVd-IIa) and the causal agents of severe (CVd-IIb, CVd-IIc), more moderate (Ca903) and mild (Ca909).
PDF Abstract XML References Citation
Received: July 21, 2011;
Accepted: September 25, 2011;
Published: December 02, 2011
How to cite this article
A.R. Sofy, A.M. Soliman, A.A. Mousa and Kh.A. El-Dougdoug, 2012. Molecular Characterization and Bioinformatics Analysis of Viroid Isolate Associated with Citrus Gummy Bark Disease in Egypt. International Journal of Virology, 8: 133-150.
DOI: 10.3923/ijv.2012.133.150
URL: https://scialert.net/abstract/?doi=ijv.2012.133.150
DOI: 10.3923/ijv.2012.133.150
URL: https://scialert.net/abstract/?doi=ijv.2012.133.150
INTRODUCTION
The disorder of trees of sweet orange grafted on sour orange rootstock in Egypt, was described by Nour-Eldin (1956, 1959) as phloem discoloration of sweet orange. Further study by Nour-Eldin (1968) showed gummy deposits are largely in the outer bark tissues, then the name was changed to gummy bark of sweet orange. When the bark of affected trees is scraped, a line of reddish-brown, gum-impregnated tissue can be seen around the circumference and especially near the bud union. The discoloration and gumming may extend 60 cm or more above the bud union in the bark of the trunk and main branches of the sweet orange. The causal agent of citrus gummy bark disease has been transmitted experimentally by bud inoculation from infected trees to sweet orange; however, symptoms are visible only after 5-10 years incubation period (Nour-Eldin, 1968; Roistacher, 1991; Onelge et al., 1996). Etiology of gummy bark disease is unknown but viroids have been suggested as the causal agent. sPAGE and RT-PCR results indicated that, CVd-II associated with gummy bark disease as reported by Onelge et al. (2004) and Sofy et al. (2010).
Hop stunt viroid (HSVd; synonym: citrus viroid [CVd]-II) is the causal agents of citrus cachexia disease which specific to mandarins and tangelos as reported by Childs (1952). Citrus Viroid (CVd) group II is comprised of Hop stunt viroid (HSVd)-related variants of 295 to 302 nucleotides. This group include; cachexia-inducing agents Citrus cachexia viroid (or CVd-IIb), CVd-IIc, Ca-903 and Ca-909 as well as the non-cachexia-inducing variant CVd-IIa (Reanwarakorn and Semancik, 1999; Palacio-Bielsa et al., 2004). Hop Stunt viroid-citrus was reported previously in Egypt associated with cachexia disease by El-Dougdoug et al. (1997) and Amin (1999). Symptoms of citrus gummy bark, a disease specific to sweet orange (Nour-Eldin, 1956, 1959) bear similarities to cachexia, a disease specific to mandarins and tangelos (Childs, 1952) caused by variants of the Hop stunt viroid (HSVd)-related CVd-II (Semancik et al., 1988). It could be suggested that a variant of this viroid may also be involved in citrus gummy bark disease expression (Onelge et al., 2004) quence analyses of the Egyptian gummy bark agent and CVd-IIb (Ca902) strain did not reveal nucleotide change or unusual mutations that might suggest a relationship to CVd-II with citrus gummy bark disease (Sofy et al., 2010).
This study aims at characterization of the viroid isolate associated with gummy bark disease which reported by Sofy et al. (2010) in Egypt using RT-PCR, sequence analysis and bioinformatics.
MATERIALS AND METHODS
Plant material: The principal gummy bark isolate namely citrus viroid II used in this study was derived from an old line navel orange tree in 2008 from Kalyobiya governorate, Egypt (Sofy et al., 2010). This isolate has been maintained in sour orange as rootstock. The viroid isolate was transmitted by blind bud inoculation to Arizona 861-S1 ‘Etrog’ citrons and kept in a greenhouse under controlled temperatures (28-32°C) for at least 5 months.
Dot-blot hybridization: The fresh leaf tissue (1 g) was ground in 4 mL of grinding buffer (200 mM glycine, 100 mM Na2HPO4, 600 mM NaCl, 1% SDS, pH 9.5, autoclave for 20 min at 120°C and then added 0.1% DIECA, 0.1 mM DTT) using sterilized pestle and mortar. The extract was clarified by centrifugation for 10 min at 10000 rpm. Aliquots (10 μL) of the partially purified nucleic acids were denatured with formaldehyde denaturation buffer {5X SSC, 25 mM Na2HPO4, 5X Denhart (0.1% each of BSA, Ficoll and PVP 360), 50% deionized formamide, 200 μg mL-1 of denatured calf thymus DNA}. The mixture was incubated for 60 min at 60°C and loaded under vacuum on positively charged nylon membranes with a Bio-Dot-SF apparatus (Biorad Laboratories, Hercules, CA, USA) and immobilized by 5 min of UV treatment. Dot-blot analyses were performed using digoxigenin (Dig)-labeled cDNA probes specific for HSVd and CEVd (kindly supplied by Dr. El-Dougdoug Virology Lab. Fac. of Agric. Ain Shams Univ.). Pre-hybridization and hybridization were performed in 50% formamide and 5X SSC. The membranes were pre-hybridized at 42°C for 2 h and hybridized overnight at 50°C. After hybridization, they washed three times for 5 min each in 2X SSC, 0.1% SDS at room temperature, followed by two times for 15 min each in 0.2X SSC, 0.1% SDS at 50°C. The Dig-labeled hybrids were detected with an anti-Dig-alkaline phosphatase conjugate and visualized with the chemiluminescent substrate CSPD (Roche Diagnostic, Indianapolis, USA) (Candresse et al., 1990).
Isolation of low molecular weight RNAs: Five grams of leaf tissues of each citrus tree was triturated in 5 mL of extraction buffer (0.4 M Tris-HCl, pH 8.9; 1% SDS; 5 mM EDTA, pH 7.0; 4% β-mercaptoethanol) and 15 mL of water-saturated phenol neutralized at pH 7.0. The nucleic acids were partitioned in 4 M LiCl and the soluble fraction was concentrated by ethanol precipitation and resuspended in 300 μL TKM buffer (10 mM Tris, 10 mM KCl, 0.1 mM MgCl2, pH 7.4) (Semancik et al., 1975).
Polyacrylamide gel electrophoresis: Aliquots (20 μL) of partially purified RNAs were analyzed by 5% sPAGE. The first gel was polymerized in TAE buffer (40 mM Tris, 20 mM sodium acetate, 1 mM EDTA, pH 7.2) and subjected to a constant current of 54 mA at 4°C for 3 h. A segment of the gel defined by the xylene-cyanol dye and 1 cm below was excised and placed on the top of a second gel containing 8 M urea and polymerized with TBE buffer (22.25 mM Tris, 22.25 mM boric acid, 0.5 mM EDTA, pH 8.3). The second gel was subjected to a constant current of 15 mA at room temperature for 4 h and stained with silver nitrate (Iglio, 1983; Rivera-Bustamante et al., 1986).
RT-PCR: Total RNA was extracted using RNA Minipreps Super Kit (BIO BASIC, Cat. # BS584). The resulting RNA preparations were subjected to RT-PCR amplification. Samples were heated to 70°C for 5 min in the presence of the HSVd-specific oligonucleotide 26-mer HSVd(-) (5`-dAGGGGCTCAAGAGAGGATCCGCGGCA-3`) and immediately cooled on ice. The mixture was then subjected to Reverse Transcription (RT) with moloney murine leukemia virus reverse transcriptase (Promega Corp., Madison, WI, USA) at 37°C for 60 min in 25 μL volume. One-tenth of the RT product was directly subjected to PCR amplification in a 25 μL volume reaction in the presence of the oligonucleotide HSVd(-) and the 24-mer HSVd(+) (5`-dCTGGGGAATTCTCGAGTTGCCGCA-3`) (El-Dougdoug et al., 2010). PCR cycling parameters were: denaturation at 94°C for 2 min, followed by 35 cycles of 94°C for 1 min, 65°C for 1 min and 72°C for 2 min, to finish with extension at 72°C for 10 min. PCR-amplified DNA fragments were separated by agarose gel electrophoresis in 1.5% agarose (Seakem LE, FMC, Bio products, Cat. # 50004) minigels in 0.5X TBE buffer (Tris-borate-EDTA, 90 mM Tris acetate, 90 mM boric acid, 2 mM EDTA, pH 8.0) using 100 bp DNA ladder (Promega, Cat # G2101). The gels were visualized with UV light after staining for 10 min with 10 μg mL-1 of ethidium bromide [2,7-Diamino-10-ethyl-9-phenylphenanthridinium bromide; homidium bromide. (Sigma, Cat. # E7637)] (Sambrook et al., 1989) and visualized with UV illumination using Gel Documentation System (Gel Doc 2000 BIO RAD 1000/115 V ~ 50/60 Hz-150 VA).
Cloning and sequence analysis: The amplified RT-PCR products were purified from the agarose gel using the QIAEX II Agarose Gel Extraction Protocol (Qiagene, Cat. # 20021). The resulting fragments were cloned into the pGEM®-T vector (Promega) and transformed into E. coli DH5α. The cDNA clones from all isolates were identified by PCR and restriction analysis. Selected clones were sequenced using an automated DNA sequencer (ABI Prism 3730XL DNA Analyzer) and analyzed by FinchTVTM version 1.4.0 software. The sequence was reported to GenBank and are archived under the accession number : FJ984562 (Sofy et al., 2010). Sequences were aligned with the Egyptian CVd-II sequence that was archived in GenBank database (accession number: FJ984562) using ClustalW (Ver.1.74) program (Thompson et al., 1994). The nucleotide distances were estimated considering alignment gaps by using Jukes and Cantor (1969) method for correction of superimposed substitutions using the Molecular Evolutionary Genetics Analysis (MEGA) software (Ver. 4.0) (Tamura et al., 2007). Neighbour Joining (NJ) implemented through MEGA 4.0 software and bootstrap analysis (1000 replicates) was performed to assess the reliability of the constructed phylogenetic tree.
Prediction of the secondary structure of viroid RNA: Secondary structures were obtained with the MFOLD program (circular version) of the GCG package (Zucker, 1989) at 37°C. Circular structures were obtained with CLC RNA Workbench software, Denmark.
RESULTS
Primary characterization of gummy bark-causing agent: The principal gummy bark disease agent was collected from an old line navel orange tree in 2008 in Kalyobiya governorate exhibited vein chlorosis leaf, gummy bark symptoms and tree stunted. When the bark was scraped, a line of reddish-brown, gum-impregnated tissue can be seen around the circumference and especially near the bud union. The discoloration and gumming may extend 60 cm or more above the bud union in the bark of the trunk. Also, dark brown streaks of gum-impregnated tissue were seen in both the circumference and in longitudinal sections (Fig. 1).
Dot blot hybridization was used to confirm the source of gummy bark infected with viroids. The samples gave clear positive with HSVd DIG-labeled probe compared with positive and negative control. Also, the source of gummy bark not infected with Citrus Exocortics Viroid (CEVd) using CEVd Dig-labelled probe compared with positive and negative control (Fig. 2). HSVd-EG isolate (cv. Navel) was indexed by graft inoculation on Etrog citron plants. It gave petiole wrinkle and midvein browning symptoms appeared after 12-16 weeks.
 | |
| Fig. 1: | Infected citrus navel orange tree in the field showing different types of gummy bark symptoms as discoloration of phloem tissues and vein chlorosis in leaves |
 | |
| Fig. 2(a, b): | Nitrocellulose membrane was spotted with extracted leaves from citrus cv. Navel tree appeared gummy bark and grafted Etrog citron using (a) HSVd Dig-labeled cDNA probe and (b) CEVd Dig-labeled cDNA probe by dot-blot hybridization |
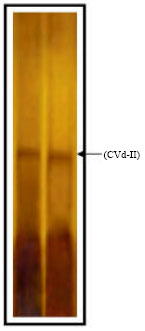 | |
| Fig. 3: | Five Percent S-PAGE showing RNA bands extracted from citrus cv. Navel tree appeared gummy bark symptoms and grafted Etrog citron stained with silver nitrate |
S-PAGE for gummy bark-causing agent: The RNA of gummy bark-causing agent that was extracted from citrus cv. Navel tree appeared gummy bark and grafted Etrog citron was determined by sPAGE. It was found an identical relative mobility on sPAGE with the electrophoretic mobility of HSVd (Fig. 3).
Molecular characterization of gummy bark-causing agent:
| • | Total RNA extraction: Total RNA was extracted, from citrus cv. Navel tree appeared gummy bark and grafted Etrog citron, using RNA Minipreps Super Kit (BIO BASIC, Cat. # BS584). The integrity and quantity of the purified RNA were confirmed by gel electrophoresis and UV spectrophotometer. The concentration of RNA was 45 μg 0.5 g-1 of infected tissues and the purity of the total RNAs obtained measured by an A260/280 absorbance ratio (1.8) for indicating high yield and purity of the extracted RNAs |
| • | Amplification of the gummy bark-causing agent: Gummy bark-causing agent RNA that was prepared previously, was reversed by M-MLV reverse transcriptase. The resulting cDNA was amplified by PCR using primers specific for HSVd. The RT-PCR products were analyzed using 1.5% agarose gel by electrophoresis. The expected size of amplified cDNA was approximately 300 bp. While the healthy leaves have no reaction (Fig. 4a) |
| • | Cloning of gummy bark-causing agent into pGEM®-T Easy Vector: pGEM®-T Easy Vector System I (Promega, Cat # A1360) provides a highly efficient one step cloning strategy for the direct insertion of Taq polymerase-amplified PCR products into a plasmid vector for regulated expression in E. coli DH5α |
| • | Isolation of recombinant plasmids and Enzyme digestion: Several white colonies resistant to ampicillin were selected to test for recombinant plasmids containing the gummy bark agent (~300 bp). PCR was done to confirm if the transformation of pGEM®-T Easy Vector with gummy bark agent into strain DH5α bacterial strain was carried out or not (Fig. 4b). Restriction enzyme digestion with EcoRI representing insert in the right direction at ~320 bp (Fig. 4c) |
Nucleotide sequence analysis: Nucleotide sequencing of the RT-PCR amplified fragment in the recombinant plasmid for the gummy bark-causing agent was completed to confirm PCR fragment is belonging to pospiviroidae group and to compare the sequence with other sequences of citrus viroids available in GenBank (Fig. 5).
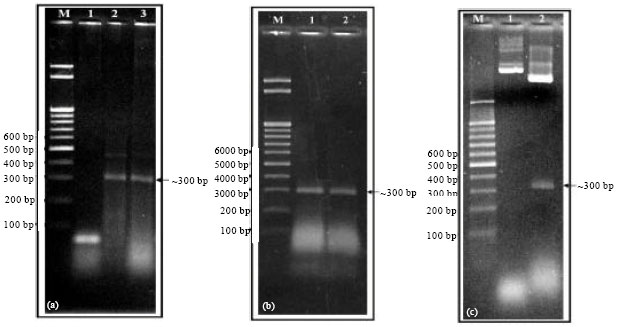 | |
| Fig. 4(a-c): | 1.5% agarose gel electrophoresis showing: (a) RT-PCR products amplified from total RNA extracted from citrus cv. Navel tree appeared gummy bark and grafted Etrog citron infected with Egyptian isolate of HSVd using specific primer Lanes (Lanes 2 and 3) as well as healthy Lane (1). (b) PCR products of two transformed colonies from the transformation plate (Lanes 1 and 2). and (c) Enzyme digestion of the recombinant plasmids from the cloning of the RT-PCR amplified product where Lane (1): DNA minipreps before digestion and Lane (2): DNA minipreps digested with EcoRI. M: DNA molecular weight marker (100 bp ladder) is indicated on the left. Arrow indicates the location of the products |
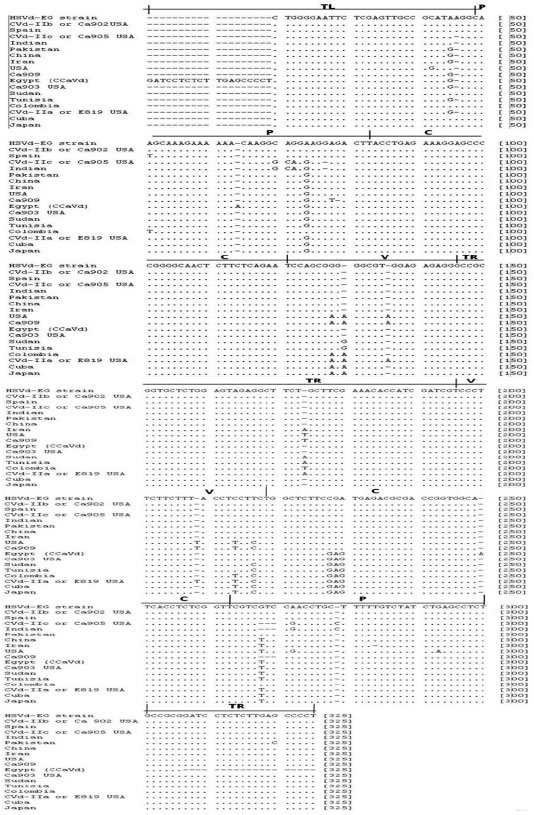 | |
| Fig. 5: | Alignment of HSVd-EG isolate (accession No. FJ984562) and 17 HSVd-citrus sequences using ClustalW program resulting in 325 positions including the gaps. Domains: TL (terminal left), P (pathogenic), C (central), V (variable) and TR (terminal right) (Keese and Symons, 1985) |
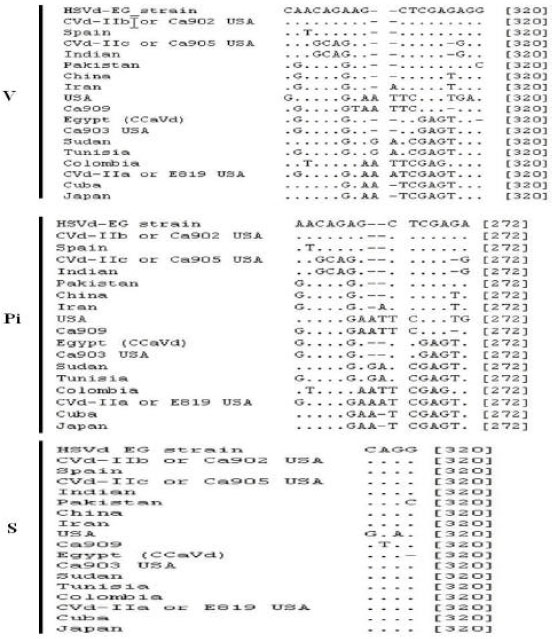 | |
| Fig. 6: | Nucleotide diversity of HSVd-EG strain with 17 HSVd-citrus isolates showing the characteristic changes variable (V), parsimoniously informative nucleotide (Pi) and singleton (S) sites |
Bioinformatics analysis for characterization of gummy bark-causing agent
Statistical analysis of sequence and alignment: For sequence heterogeneity within HSVd-EG isolate, the sequence was aligned using CVd-IIa, CVd-IIb, CVd-IIc, Ca903 and Ca909 as the original reference sequences for HSVd-citrus. In addition to that, genetic diversity of the HSVd-EG isolate characterized in this work was carried out together with the 17 reported HSVd-citrus sequences from Egypt, Spain, USA, Pakistan, Cuba, Iran, Colombia, Japan, Indian, Tunisia, China and Sudan. All of these sequences were multiple-aligned using the ClustalW program with minor manual adjustments, resulting in 325 positions including the gaps (Fig. 5). Total of 20 variable sites were found in HSVd and 16 were parsimoniously informative nucleotide sites as well as 4 were singleton sites (Fig. 6).
| Table 1: | Comparison between bases composition of complete genome sequence for HSVd-EG isolate (Accession No. FJ984562) and 17 HSVd-citrus isolates published in GenBank |
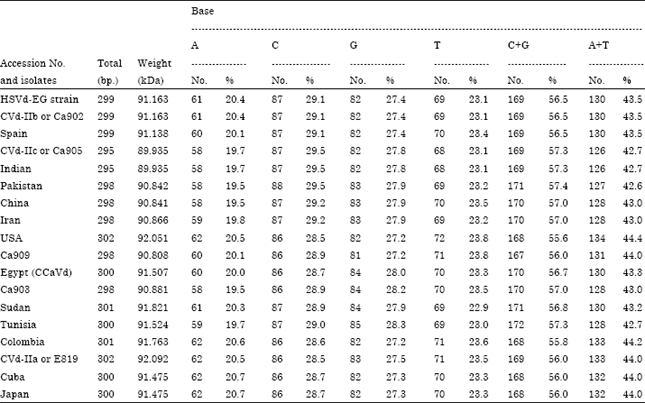 | |
The nucleotide sequence of complete genome for HSVd-EG strain (Accession No. FJ984562) revealed highest content for Cytosine (C) 87 (29.1%) followed by Guanine (G) 82 (27.4%), then Thymine (T) 69 (23.1%) and Adenine (A) 61 (20.4%). Data also showed that, the ratio between C+G to A+T was 1.3 (Table 1). Moreover, bases composition data for complete genome sequence of HSVd-EG (gummy bark agent) and 17 isolates in GenBank was tabulated to determine C+G and A+T ratio (Table 1).
The nucleotide distances between isolates ranged from 0.000 to 0.042 (Table 2). The lower values were recorded for isolate pairs HSVd-EG with CVd-IIb (1-2), CVd-IIc with Indian (4-5), China with Iran (7-8), CCaVd Egyptian isolate with Ca903 (11-12) and Cuba with Japan (17-18) confirming that these isolates had the same nucleotide sequences (Table 2). The higher nucleotide distance values were recorded for isolate pairs CVd-IIc with Colombia (4-15) and Indian with Colombia (5-15) (Table 2). Genetic diversity value between HSVd populations, representing the average number of nucleotide substitutions per site between pairs of sequences, was 0.021±0.005 (Table 2).
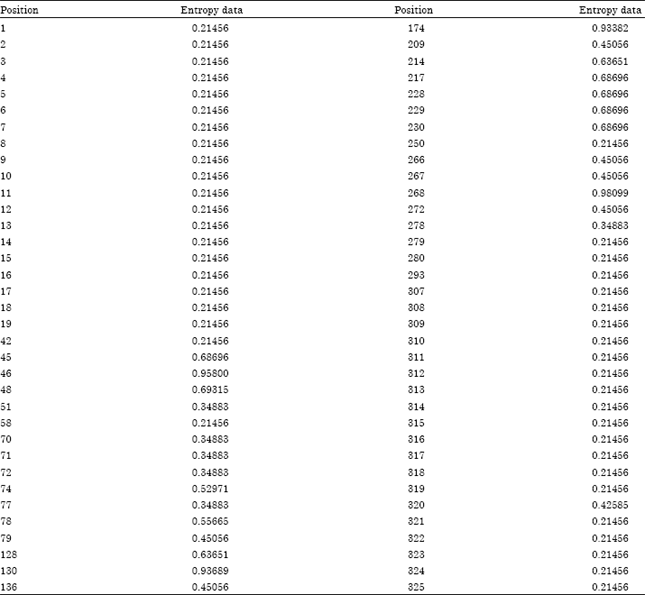
Entropy power explains the sequence mutates by the function of entropy plot using BIOEDIT. As shown in Fig. 7, there are several peaks which contains high variable regions. So entropy power can be used to screen the variable regions (Table 3).
A phylogenetic tree was constructed for showing the clustering relationship among HSVd-citrus populations (Fig. 8). It, generated using the Neighbour-Joining method and bootstrap analysis of 1000 repetitions. It produced two distinct groups. The first group is Asian strain with USA, Spain and Egypt. Second group is African including Egypt and South American with Japan and USA.
| Table 2: | Nucleotide distances and standard error between HSVd-EG strain (gummy bark agent) and 17 HSVd-citrus isolates published in GenBank |
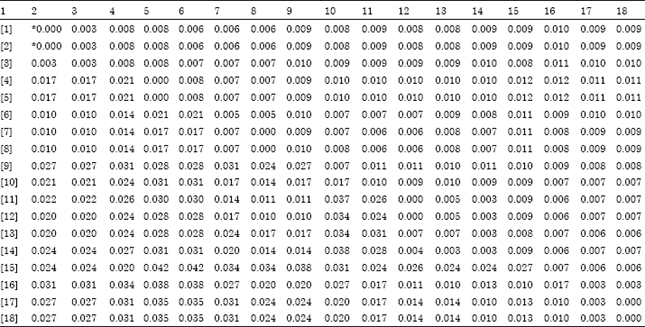 | |
| (*) Distances in lower left and Standard error in upper right (Bootstrap (1000 replicates; seed = 17114)).[1]: HSVd-EG_strain, [2]: CVd-IIb_or_Ca902_USA, [3]: Spain, [4]: CVd-IIc_or_Ca905_USA, [5]: Indian, [6]: Pakistan, [7]: China, [8]: Iran, [9]: USA, [10]: Ca909_USA, [11]: Egypt_(CCaVd), [12]: Ca903_USA, [13]: Sudan, [14]: Tunisia, [15]: Colombia, [16]: CVd-IIa_or_E819_USA, [17]: Cuba and [18]: Japan | |
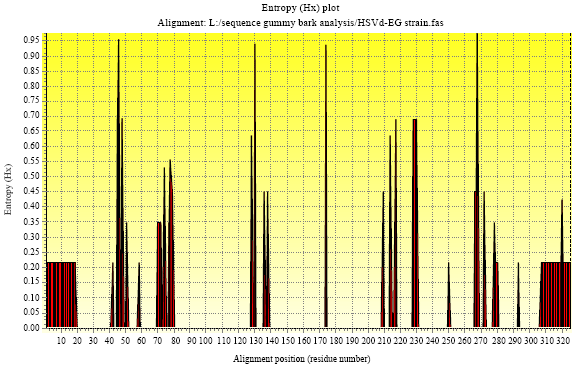 | |
| Fig. 7: | The entropy power result of HSVd-EG and 17 HSVd-citrus isolates by BIOEDIT |
| Table 3: | The entropy power data of the nucleotide variable positions of HSVd-EG and 17 HSVd-citrus isolates by BIOEDIT |
 | |
HSVd-EG (gummy bark agent) has 100, 97 and 96.5% homologous with CVd-IIb USA (accession No. aF131249), CCaVd isolate X-704-M Spain (accession No. AF213487) and CCaVd Egypt (accession No. AY513267), respectively.
Analysis of HSVd-EG-RNA structure: The primary structure of HSVd-EG-RNA is determined by the sequence. Secondary structure of HSVd-EG-RNA has been characterized by forming intramolecular base pairs among of its bases which include interior loops, hairpin loop, bulge loop and external loop (Fig. 9 and 10). On the other hand, the secondary structure of HSVd-EG-RNA differed than secondary structure of CCaVd-EG-RNA specially in pathogenic domain as shown in Fig. 9. The minimum free energy of a secondary structure for HSVd-EG-RNA is determined as-122.10 kcal moL-1 by summing the energy contribution of all base pairs, interior loops, hairpin loop, bulge loops and external loop at 37°C using MFOLD program. It is rod-shaped structure composed of alternating single-and double-stranded regions (Fig. 9).
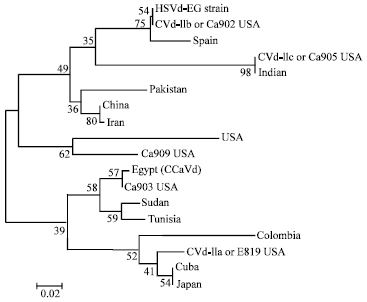 | |
| Fig. 8: | Neighbour-joining tree of HSVd-EG isolate (gummy bark agent) and 17 isolates of HSVd-citrus published in GenBank. Numbers represent bootstrap percentage values based on 1000 replicates |
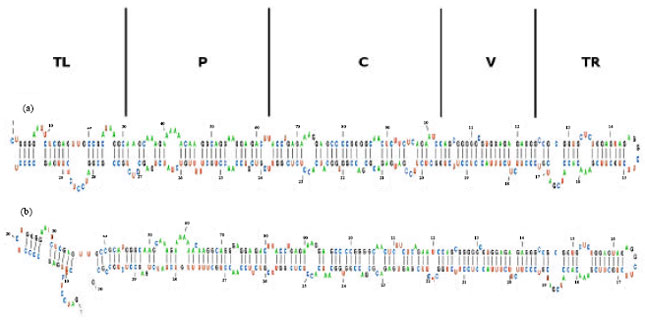 | |
| Fig. 9(a-c): | The secondary structure of (a) HSVd-EG-RNA (gummy bark agent) and (b) CCaVd-EG-RNA (cachexia agent) is rod-shaped structure. Domains: TL (terminal left), P (pathogenic), C (central), V (variable) and TR (terminal right) (Keese and Symons, 1985) |
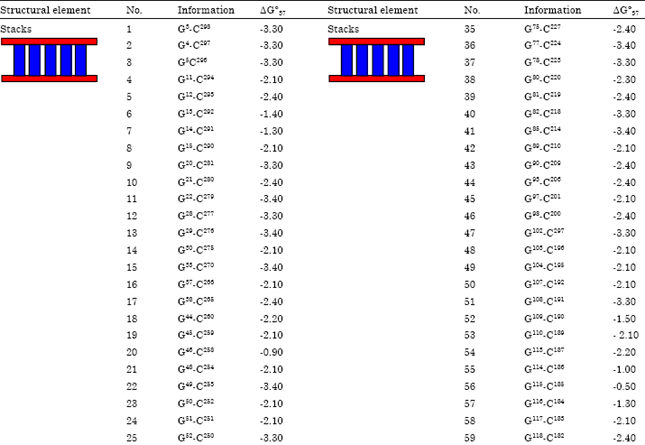
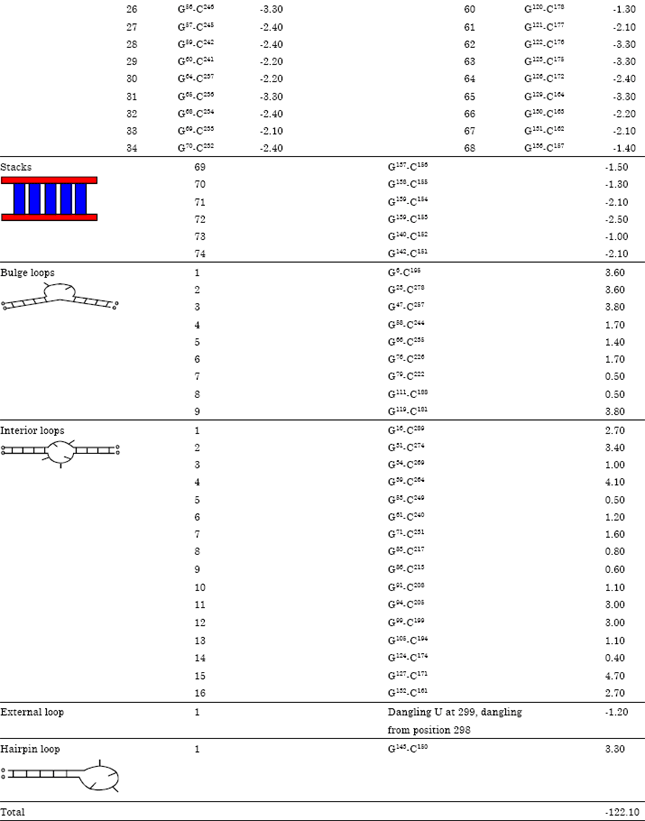
Table 4 illustrated ΔG°37 (Gibbs free energy) of stalking pairs and interior, hairpin and bulge loops depend on predicated free energy values (Kcal mol-1 at 37°C) for secondary structure of HSVd-EG-RNA. It was observed that base pairs lead to free energies <0 while bulges and loops lead to free energies >0. Consequently total free energy of HSVd-EG-isolate was -122.10 kcal moL-1. Partition function of HSVd-EG isolate (Q) sum all free energies of secondary structure. This can be calculated as base pairing probabilities for each pair of bases.
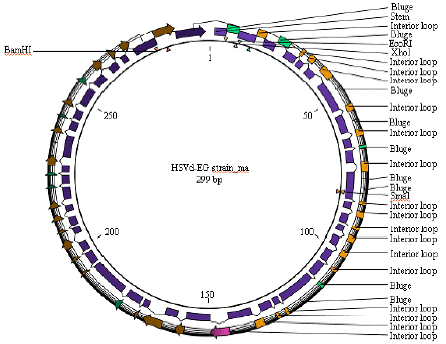 | |
| Fig. 10: | Diagram illustrated distribution of secondary structure on circular HSVd-EG-RNA |
| Table 4: | Thermodynamic of a secondary structure for HSVd-EG-RNA |
 | |
 | |
DISCUSSION
The etiology of gummy bark disease is unknown but viroids have been suggested as the causal agent. sPAGE and RT-PCR results indicated that, CVd-II could be associated with gummy bark disease as reported by Onelge et al. (2004). This study supported evidence that, the viroid associated with this disease was identified through biological and molecular characters as well as bioinformatics analysis.
Source material of this study (HSVd-EG) was isolated and propagated on healthy citrus seedlings Sour orange and Etrog citron by grafting inoculation. It was kept under greenhouse conditions 32-40°C (max. day)/27-30°C (min. night) as reported by Roistacher (1991), Duran-Vila et al. (1993) and El-Dougdoug et al. (2010). These results were confirmed by dot-blot hybridization using specific HSVd Dig-labeled cDNA probe. HSVd-EG isolate that was indexed by graft inoculation on Etrog citron plants gave petiole wrinkle and midvein browning symptoms appeared after 12-16 weeks. On the contrary, Duran-Vila et al. (1988) and Semancik et al. (1988) reported that the cachexia-xyloprorosis agents CVd-IIb and CVd-IIc are not symptomless on Etrog citron. On the other hand, Duran-Vila et al. (1993) reported that CVd-IIb and CVd-IIc induce mild symptoms in 6-9 months as incubation periods and 3 months at 23-32°C or 7 months at 18-25°C are necessary for the detection of all citrus viroids.
The HSVd-EG isolate was transmitted through grafting by eye and blind buds from naturally infected citrus cv. Navel trees to healthy Sour orange and Etrog citron plants where Nour-Eldin (1968), Roistacher (1991) and Onelge et al. (1996) reported that the causal agent of citrus gummy bark disease has been transmitted experimentally by bud inoculation from infected trees to sweet orange; however, symptoms are visible only after an incubation period of 5-10 years.
HSVd-EG-RNA was amplified by PCR after adding two primers oligo (dt) which amplify ~300 base fragment of RNA (Fig. 4a-c) and this is in harmony with Reanwarakorn and Semancik (1998) and El-Dougdoug et al. (2010). Sequence of HSVd-EG isolate is 299 nucleotides in length as reported by Sofy et al. (2010) where submitted to GenBank under accession No. FJ984562. The nucleotide sequence of complete genome for HSVd-EG strain (Accession No. FJ984562) revealed the highest content for Cytosine (C) 87 (29.1%) followed by Guanine (G) 82 (27.4%), then Thymine (T) 69 (23.1%) and Adenine (A) 61 (20.4%) (Table 1).
Genetic diversity and evaluation of entropy power for the Egyptian citrus gummy bark agent (Accession No. FJ984562) and HSVd-citrus populations registered in GenBank, were viewed against the phylogenetic background of known Cvd-II variants including the non-cachexia (CVd-IIa) and the causal agents of severe (CVd-IIb, CVd-IIc), more moderate (Ca903) and mild (Ca909) cachexia (Fig. 7 and 8) (Table 3) as reported by Reanwarakorn and Semancik (1999). The relationships between CVd-II related clone recovered from the citrus gummy bark symptomatic tree and HSVd-citrus sequence registered in GenBank were determined by nucleotide sequence homology (Fig. 5 and 6). Phylogenetic and molecular evolutionary analyses were conducted using MEGA software version MEGA 4.0 (Tamura et al., 2007) (Fig. 8). The minimum evolution based distance method, Neighbor Joining (NJ), was utilized (Table 2) due to limited number of parsimonious informative sites of the relatively small and conserved genomes of CVd-II variants. The Jukes-Cantor estimate the number of nucleotide substitutions per site (d) ranging between 0 = d = 0.042. It is supporting the use of p-distance model for the Neighbor Joining (NJ) method. Since the viroid-sequence alignment produces series of distinct insertion/deletion events, the gap sites were included in the phylogenetic analysis (Fig. 8). That could be done using the pairwise-deletion option as well as Bootstrap analysis (1000 replicates) which assess reliability of the constructed phylogenetic tree.
Egyptian gummy bark agent (Accession No. FJ984562; Sofy et al., 2010), is distinctly defined from the non-cachexia variant (CVd-IIa) as well as from the Citrus cachexia viroid Egyptian isolate (Accession No. AY513267 (Amin, 1999). This is supported by the Jukes-Cantor method Jukes and Cantor (1969) where the genetic distance calculated between Egyptian gummy bark agent (HSVd-EG) and CVd-IIb (d = 0), CVd-IIc (d = 0.017), Ca903 (d = 0.020), Ca909 (d = 0.021), CCaVd-EG (d = 0.022) as well as with the non-cachexia CVd-IIa (d = 0.031) variant. The relationship between HSVd-EG and CVd-IIb as determined by the estimated genetic difference is identical to the value between the two cachexia variants CCaVd-EG and Ca903-USA (Table 2 and Fig. 8).
The minimum free energy of a secondary structure for HSVd-EG-RNA was determined from its primary sequence by summing the energy contribution of all base pairs, interior loops, hairpin loop, bulge loops and external loop at 37°C by MFOLD analysis, since this is the temperature at which viroids symptoms are consistently observed by Roistacher (1991) and Duran-Vila et al. (1993). The HSVd-EG sequence appeared to fold into a rod-like structure with the minimum free energy -122.1 kcal moL-1 while CCaVd-EG with the minimum free energy -120.5 kcal moL-1. Serra et al. (1995) mentioned that RNA molecule folds into that structure with the minimum free energy (mfe). Free energy models typically assume that the total free energy of a given secondary structure for a molecule is the sum of independent contributions of adjacent, or stacked, base pairs in stems (which tend to stabilize the structure) and of loops (which tend to destabilize the structure).
Symptoms of citrus gummy bark, a disease specific to sweet orange (Nour-Eldin, 1956, 1959) bear similarities to cachexia, a disease specific to mandarins and tangelos Childs (1952) caused by variants of the Hop Stunt Viroid (HSVd) related CVd-II (Semancik et al., 1988). Thus, it has been suggested that a variant of this viroid may also be either the causal agent or a factor directly involved in citrus gummy bark disease expression Onelge et al. (2004). In addition, sequence analyses of the Egyptian gummy bark agent and CVd-IIb (Ca902) strain did not reveal any common nucleotide change or unusual mutations that might suggest a relationship to CVd-II with citrus gummy bark disease. While, the variant CVd-IIa does not induce any severe disease symptoms but bark cracking on Poncirus trifoliata rootstock has been attributed to CVd-IIa (Roistacher et al., 1993). This condition does not result in economic damage to trees or crop but to the contrary sweet orange with CVd-IIa exhibit an enhancement in commercial performance (Semancik et al., 2002). So, a case can be made for all CVd-II except CVd-IIa as the most probable agent in support of a viroid etiology for citrus gummy bark disease.
The previous data indicates that the high frequency of detection of Egyptian gummy bark agent as the principal CVd-II-related variant found in symptomatic Navel sweet orange and not found in asymptomatic trees suggests a possible relationship between that variant and citrus gummy bark but no highly distinct cachexia-like variant could be identified as a specific gummy bark disease inducing agent. With the virtual identical genome to CVd-IIb, the gummy bark symptom may constitute an expression of cachexia disease in sweet orange. But the cachexia disease symptoms which reported by Reichert and Perlberger (1934). Moreover, the Egyptian gummy bark agent differ from Egyptian isolate of Citrus cachexia viroid infecting mandarin in Egypt (Amin, 1999). So, we suggest for this Egyptian strain, the name of Citrus gummy bark viroid cachexia strain with taxon identifier 648750 and scientific name Citrus gummy bark viroid related to family pospiviroidae, genus Hostuviroid and species Hop stunt viroid (http://www.uniprot.org/taxonomy/648750.)
REFERENCES
- Candresse, T., G. Macquaire, V. Brault, M. Monsion and J. Dunez, 1990. 32P-and biotin-labelled in vitro transcribed cRNA probes for the detection of Potato spindle tuber viroid and Chrysanthemum stunt viroid. Res. Virol., 141: 97-107.
PubMed - Duran-Vila, N., C.N. Roistacher, R. Rivera-Bustamante and J.S. Semancik, 1988. A definition of citrus viroid groups and their relationship to the exocortis disease. J. Gen. Virol., 69: 3069-3080.
CrossRef - El-Dougdoug, K.A., M.E. Osman, S.A. Hayam, A. Rehab, Dawoud and R.M. Elbaz, 2010. Biological and molecular detection of HSVd-infecting peach and pear trees in Egypt. Austr. J. Basic Appl. Sci., 4: 19-26.
Direct Link - Keese, P. and R.H. Symons, 1985. Domains in viroids: Evidence of intermolecular RNA rearrangements and their contribution to viroid evolution. Proc. Natl. Acad. Sci. USA., 82: 4582-4586.
PubMedDirect Link - Onelge, N., A. Cinar, J.A. Szychowski, G. Vidalakis, J.S. Semancik, 2004. Citrus viroid II variants associated with Gummy Bark disease. Eur. J. Plant Pathol., 110: 1047-1052.
CrossRef - Palacio-Bielsa, A., J. Romero-Durban and N. Duran-Vila, 2004. Characterization of citrus HSVd isolates. Arch. Virol., 149: 537-552.
CrossRef - Reanwarakorn, K. and J.S. Semancik, 1998. Regulation of pathogenicity in Hop stunt viroid-related group II citrus viroids. J. Gen. Virol., 79: 3163-3171.
Direct Link - Reanwarakorn, K. and J.S. Semancik, 1999. Correlation of Hop stunt viroid variants to cachexia and xyloporosis diseases of citrus. Phytopathology, 89: 568-574.
PubMed - Rivera-Bustamante, R.F., R. Gin and J.S. Semancik, 1986. Enhanced resolution of circular and linear molecular forms of viroid and viroid-like RNA by electrophoresis in a discontinuous-pH system. Anal Biochem., 156: 91-95.
CrossRef - Semancik, J.S., T.J. Morris, L.G. Weathers, G.F. Rodorf and D.R. Kearns, 1975. Physical properties of a minimal infectious RNA (viroid) associated with the exocortis disease. Virology, 63: 160-167.
CrossRef - Semancik, J.S., C.N. Roistacher, R. Rivera-Bustamante and N. Duran-Vila, 1988. Citrus cachexia viroid, a new viroid of citrus: Relationships to viroids of the exocortis disease complex. J. Gen. Virol., 69: 3059-3068.
Direct Link - Serra, M.J., D.H. Turner and S.M. Freier, 1995. Predicting thermodynamic properties of RNA. Methods Enzymol., 259: 243-261.
CrossRefPubMedDirect Link - Sofy, A.R., A.M. Soliman, A.A. Mousa, S.A. Ghazal and K.H.A. El-Dougdoug, 2010. First record of Citrus viroid II (CVd-II) associated with gummy bark disease in sweet orange (Citrus sinensis) in Egypt. New Dis. Rep., 21: 24-24.
CrossRefDirect Link - Tamura, K., J. Dudley, M. Nei and S. Kumar, 2007. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol., 24: 1596-1599.
CrossRefPubMedDirect Link - Thompson, J.D., D.G. Higgins and T.J. Gibson, 1994. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res., 22: 4673-4680.
CrossRefPubMedDirect Link