
A. Ausama Yousif
Central Biotechnology Laboratory, College of Veterinary Medicine and Animal Resources, Bldg. 999, King Faisal University, Saudi Arabia
A. Abdelmohsen Al-Naeem
Department of Clinical Studies, Epidemiology Section, College of Veterinary Medicine and Animal Resources, Bldg. 999, King Faisal University, Saudi Arabia
International Journal of Virology
Year: 2011 | Volume: 7 | Issue: 4 | Page No.: 135-146







ABSTRACT
Camelpox virus (CMLV) belongs to the genus Orthopoxvirus (OPV) of Poxviridae. CMLV causes clinical disease in camel populations worldwide. CMLV has been known in Saudi Arabia for centuries. However, no attempts have been made to characterize the endemic CMLV genetically. This situation leads to the inability to monitor the molecular epidemiology of the virus in the environment. It also reduces our capability to evaluate vaccine-based control strategies. CMLV continues to persist in nature and re-emerge. We were able to isolate a CMLV from an outbreak in the Eastern Provence of KSA in 2009. The infection produced mild clinical signs in adults and camel calves. We were able to confirm that the isolate was indeed CMLV using PCR and partial sequencing of the A-type inclusion protein (ATIP) gene. The Orthopoxvirus (OPV) ortholog genes L1R, A27L, A33R and B5R of the Saudi enzootic CMLV were also investigated using a modified PCR assay. Our data explains some of the variation obtained with restriction analysis and underlines the need for a review of some of the vaccines used to control CMLV.
PDF Abstract XML References Citation
Received: June 20, 2011;
Accepted: July 27, 2011;
Published: October 22, 2011
How to cite this article
A. Ausama Yousif and A. Abdelmohsen Al-Naeem, 2011. Molecular Characterization of Enzootic Camelpox Virus in the Eastern Kingdom of Saudi Arabia. International Journal of Virology, 7: 135-146.
DOI: 10.3923/ijv.2011.135.146
URL: https://scialert.net/abstract/?doi=ijv.2011.135.146
DOI: 10.3923/ijv.2011.135.146
URL: https://scialert.net/abstract/?doi=ijv.2011.135.146
INTRODUCTION
Camelpox occurs in almost every country in which camel husbandry is practiced apart from the introduced dromedary camel in Australia and tylopods (llama and related species) in South America. Outbreaks have been reported in the Middle East (Bahrain, Iran, Iraq, Oman, Saudi Arabia, United Arab Emirates and Yemen), in Asia (Afghanistan and Pakistan), in Africa (Algeria, Egypt, Ethiopia, Kenya, Mauritania, Morocco, Niger, Somalia and Sudan) (Wernery et al., 1997) and in the southern parts of Russia and India. The first outbreak of CMLV was reported recently in Syria (AlZi’abi et al., 2007). The disease is endemic in these countries and a pattern of sporadic outbreaks occurs with a rise in the seasonal incidence usually during the rainy season (OIE, 2009).
In Saudi Arabia, the first report was in 1986 (Alhendi et al., 1994). Since then, CMLV was reported in the eastern region of Saudi Arabia in 1994 with 10% morbidity and no mortalities (Alhendi et al., 1994). In Al-Ahsa region, another outbreak of CMP was reported in 1999 with 100% morbidity and, again, no mortalities (Abu Elzein et al., 1999). More recently, CMLV was reported in Jazan with 41% morbidity and 3.6% mortality (Dafalla and Elfadel, 2007).
Camelpox is characterized by fever, local or generalized pox lesions on the skin and in the mucous membranes of the mouth, respiratory and digestive tracts. The clinical manifestations range from inapparent infection to mild, moderate and less commonly, severe systemic infection and death. Severe camelpox outbreaks in naïve young camelids cause high mortalities (OIE, 2009). The variation in the severity of clinical signs possibly reflects differences between the strains of CMLV (Wernery and Kaaden, 2002). The infection may also spread to the hands of camel drivers (Carter et al., 2005; AlZi’abi et al., 2007).
Camelpox is caused by a poxvirus. Members of Family Poxviridae, subfamily Chordopoxvirinae, genus Orthopoxvirus are among the best and longest known viruses to mankind and they are among the most feared viruses of livestock animals and humans. Viruses of the family Poxviridae have large and complex virions with a size of approximately 140-260 nmx220-450 nm that is large enough to be visible by light microscopy. The genome is a double-stranded DNA molecule of 130-375 kbp, with the genome size varying between genera and from strain to strain. Genetic studies showed that the central part (approximately 100 kbp) of the genome is highly conserved in gene order, content and sequences whereas the terminal regions vary both in length and patterns of restriction enzyme cleavage sites (Essbauer et al., 2010).
Viruses belonging to genus Orthopoxvirus (OPV) are antigenically closely related. Members/species include Variola major virus (VAR), Monkeypox virus (MPV), Camelpox virus (CMLV), Vaccinia virus (VAC), Cowpox virus (CPV) and six other species not known to be pathogenic for humans (Panning et al., 2004; Essbauer et al., 2010).
The presumptive preliminary diagnosis depends on clinical signs. However, early and mild clinical signs can be confused with a number of other disease conditions like contagious ecthyma (orf), papilloma virus and reaction to insect bites. Confirmatory diagnosis is essential for proper diagnosis. Diagnostic techniques include TEM, demonstration of CMLV antigens in tissues and scabs by immunohistochemistry, isolation on the chorioallantoic membrane of embryonated chicken eggs and detection of viral nucleic acid using PCR and restriction analysis (OIE, 2009). Today, PCR, real-time PCR and sequencing are the methods of choice for rapid identification and differentiation of poxviruses up to the species level (Mercer et al., 2007; Balamurugan et al., 2009). Diagnostic procedures based on sampling skin are preferred since scabs and other skin lesions are easily sampled, the virus is stable in scabs and lesions appear 1-3 days after the onset of fever and may be present long after fever subsides.
Despite the fact that CMLV has been known in Saudi Arabia for centuries, no attempts have been made to characterize the endemic CMLV genetically. This situation leads to the inability to monitor the molecular epidemiology of the virus in the environment. It also reduces our capability to evaluate vaccine-based control strategies. In this publication, we report the first molecular characterization of the enzootic CMLV in the KSA.
MATERIALS AND METHODS
Samples: Three Non-febrile adult camels and camel calves (6 and 8 months old) showing mild skin lesions were sampled. Animals sampled belonged to a single herd of fifteen camels that was partially fed by grazing in the Eastern Province of Saudi Arabia. Minute scabs loosely attached to the camel coat or crusted scabs which were harder to remove from the camel skin were collected in sterile cups and transported dry to the Central Biotechnology Laboratory, College of Veterinary Medicine, King Faisal University, Saudi Arabia. Samples were kept at -80°C until processing. Blood was collected from the jugular vein of two camel calves aged 6 and 8 showing very mild crusted pox lesions. Blood was aliquoted and preserved at -80°C until processing.
Total DNA extraction from skin lesions
The FNES technique: Ten milligrams (0.01 g) of the sampled scabs and negative camel tissue (negative control) were weighed, transferred to a sterile porcelain mortar and ground in 500 μL of freshly prepared extraction buffer AusEx3 [100 mM Tris pH 8.0, 50 mM EDTA pH 8.0, 500 mM NaCl, 1.5% SDS, 0.3% 2-ME]. AusEx3 is modified from EB described by Hosaka (2004). The 200 μL from the tissue homogenate of each sample were then transferred to a 1.5 mL microfuge and 64 μL of 3 M potassium acetate were added to precipitate the SDS- protein complex. The tubes were centrifuged at 20800x g for 20 sec and 120 μL (up to 200 μL can be used) of each extract were transferred to a fresh nuclease-free microfuge tube. Each sample is processed in less than three minutes. The technique was termed FNES technique because it is fast, non-enzymatic and safe. DNA extracts were kept at -20°C until they were analyzed by PCR and spectrophotometry. Each sample was extracted three times using the FNES technique to test reproducibility of the results.
DNA extraction using commercial kits: Qiagen DNeasy Blood and Tissue Kit (QIAGEN Sciences, Maryland, USA) was used for the extraction of DNA from scabs, blood and an experimental OPV vaccine (provided by the Ministry of Agriculture, Saudi Arabia). Extractions were performed according to the manufacturer’s recommendations using 20 mg of scabs, 50 μL of blood and 50 μL of reconstituted vaccine as starting materials. DNA extracts were kept frozen at -20°C until tested using PCR and spectrophotometry.
Extraction was performed according the manufacturer’s recommendations using 20 mg of scabs, 50 μL of blood and 50 μL of reconstituted vaccine as starting material. DNA extracts were kept at -20°C until use.
Primers: Primers that amplify a region of the Acidophilic-Type Inclusion Protein (ATIP) gene (Meyer et al., 1994) were used to detect the presence of OPV sequences in DNA from the skin lesions of sampled camels. The sequence of the forward primer (ATI up 1) was 5`- AAT ACA AGG AGG ATCT-3`and the reverse primer (ATI low 1) was 5`- CTT AAC TTT TTC TTT CTC-3`. In addition, two extracellular enveloped virus (EEV)-specific ortholog genes (A33R and B5R) were targeted for amplification. The primers used were A33R upper (5`-GGC ATA TGA TGA CAC CAG AAA ACG-3`), A33R lower (5`-GGC TCG AGT TAG TTC ATT GTT TTA ACA C-3`), B5R upper (5`-GGC ATA TGA AAA CGA TTT CCG TTG TTA CG-3`) and B5R lower (5`-GGC TCG AGT TAC GGT AGC AAT TTA TGG-3`) (Hooper et al., 2004). Also, two Intracellular Mature Virus (IMV) genes (L1R and A27L) were also targeted for amplification. The primers used were L1R upper (5`-GGC ATA TGG GTG CCG CAG CAA GC-3`), L1R lower (5`-GGC TCG AGT CAG TTT TGC ATA TCC G-3`), A27L upper (5`-GGC CAT GGA CGG AAC TCT TTT CCC CG-3`) and A27L lower (5`- GGC TCG AGC TCA TAT GGA CGC CGT CC-3`) (Hooper et al., 2004). A33R, B5R, L1R and A27L amplification reaction conditions were not described previously.
PCR: ATIP, A33R, B5R, L1R and A27L amplification reactions were carried out using the HotStar Taq® Plus Master Mix PCR kit (QIAGEN, Hilden, Germany). Each reaction tube contained one unit of HotStar Taq® Plus DNA polymerase in 1X PCR buffer containing (KCl and (NH4)2SO4), 200 μM of each dNTP, 0.5 μM of each of the forward and reverse primers, 1X CoralLoad dye mix and 3 μL of a 1:50 dilution of each of the samples and controls.
The reaction tubes were centrifuged at 2000x g briefly and placed in a BioRad MyCyclerTM thermal cycler (BioRad, Hercules, California, USA). ATIP PCR reactions conditions were described previously (Meyer et al., 1994). Thermal cycling conditions for amplification of the A33R and B5R ortholog genes were: an initial denaturation step (95°C for 4 min), 15 first-stage amplification cycles (95°C for 1 min, 44°C for 1 min and 72°C for 2.5 min), 25 sec stage amplification cycles (95°C for 30 sec, 46°C for 30 sec and 72°C for 2.5 min) and a final extension step (72°C for 5 min). Thermal cycling conditions for amplification of the L1R ortholog gene was: an initial denaturation step (95°C for 4 min), 15 first-stage amplification cycles (95°C for 1 min, 60°C for 1 min and 72°C for 2.5 min), 25 sec stage amplification cycles (95°C for 30 sec, 70°C for 30 sec and 72°C for 2.5 min) and a final extension step (72°C for 5 min). Thermal cycling conditions for amplification of the A27L ortholog gene was: an initial denaturation step (95°C for 4 min), 15 first-stage amplification cycles (95°C for 1 min, 50°C for 1 min and 72°C for 2.5 min), 25 sec stage amplification cycles (95°C for 30 sec, 55°C for 30 sec and 72°C for 2.5 min) and a final extension step (72°C for 5 min). After amplification, 15 μL were analyzed by electrophoresis on a 1.2% agarose gel stained with ethidium bromide. DNA bands were visualized by UV irradiation in a Gel Doc XR gel documentation system (BIO-RAD laboratories, Milan, Italy).
Sequencing: PCR products were purified from gels using QIAEX® Gel Extraction Kit (QIAGEN Sciences, Maryland, USA). Purified PCR products were sequenced in both orientations by the dideoxy chain-termination method using the amplification primers described earlier. The nucleotide and amino acid sequences of these genes were aligned using CLUSTAL W (Thompson et al., 1994) and BLAST 2.0 search program (National center for Biotechnology Information (NCBI) (Altschul et al., 1997).
Virus isolation on the chorioallantoic membrane of embryonated chicken eggs: Skin lesions were processed for viral isolation and inoculated on the Chorioallantoic Membrane (CAM) of 11-13 day old SPF Embryonated Chicken Eggs (ECE). The eggs were incubated at 35°C degrees and after 5 days, the eggs containing living embryos were refrigerated overnight. Inoculated eggs were then opened and the CAM examined for the presence of characteristic pock lesions: dense, grayish-white or lighter less elevated with hemorrhagic centers. Two passages were done on the CAM of ECE.
Histopathology: Paraffin sections from the tissue specimens were prepared and stained with Haematoxyline and Eosin according to Bancroft et al. (1996). Briefly, the slide were deparaffinize in 2 changes of xylene, 15 min each and hydrate to water by descending grades of alcohol (95, 80 and 70%) 10 min each. Mayer’s hematoxylin was added on slides for 15 min then washed in running tap water for 20 min and counterstained with eosin for 2 min. The sides were dehydrated in 95% and absolute alcohols, two changes of 2 min each until excess eosin was removed, then cleared in xylene (two changes of 2 min each) and mounted in Canada balsam.
RESULTS
PCR amplification of CMLV genes: Successful amplification of the three OPV genes tested was achieved. A27L gene PCR products (362 bp) were successfully amplified from extracts of skin lesions of camels with clear CMLV lesions (Fig. 1, lane 2). A27L was not amplified from the OPV control (Fig. 1, lane 1). A33R gene PCR products (568 bp) were successfully amplified from extracts of skin lesions of camels with clear CMLV lesions as well as the OPV control (Fig. 1, lanes 4 and 3, respectively). B5R gene PCR products (967 bp) were successfully amplified from extracts of skin lesions of camels with clear CMLV lesions as well as the OPV control (Fig. 1, lanes 6 and 5, respectively).
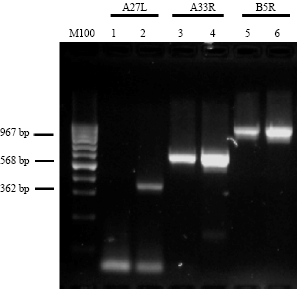 | |
| Fig. 1: | PCR product analysis of CMLV A27L, A33R and B5R orthologs.. A27L, A33R and B5R primers (Hooper et al., 2004) were used to amplify OPV sequences from 1:50 dilutions of DNA extracts of camel skin samples and an OPV vaccine. 100 bp DNA ladder is the far left of the figure. Lanes 1 and 2: A27L PCR products for OPV and CMLV, respectively. Lanes 3 and 4: A33R PCR products for OPV and CMLV, respectively. Lanes 5 and 6: B5R PCR products for OPV and CMLV, respectively. Notice that the OPV PCR for A27L was negative. All other gene amplifications produced comparable results for both OPV and CMLV |
ATIP gene PCR products (881 bp) were successfully amplified from extracts of skin lesions (Fig. 2, lane 2). ATIP gene PCR product of about 1.5 Kbp was amplified from extracts of the control OPV (Fig. 2, lane 1). Both control OPV and CMLV produced the 765 bp PCR product expected for the L1R gene amplification. The PCR products of A33R and B5R ortholog genes amplification from the FNES technique DNA extracts were comparable to those obtained from the spin-column technique extracts (data not shown).
Isolation on the CAM of SPF ECE: The first passage of CMLV of the CAM of SPF ECE at 35°C produced the characteristic pock lesions: light slightly elevated pock lesions with hemorrhagic centers (Fig. 3c). The second passage performed at 38°C produced numerous characteristic pock lesions: dense grayish-white with elevated centers (Fig. 3b). Seven to ten percent of embryos developed hemorrhagic pitichae upon inoculation with CMLV (Fig. 3a). Histopathological examination of the CAM of SPF ECE inoculated with CMLV showed localized regions of thickening in the CAM (Fig. 4a. Lower arrow) compared to the normal areas of the CAM (Fig. 4a. Upper arrow). In membranes with visible hemorrhagic pock lesions, there was evidence of an increase in small blood vessel supply to the membrane (Fig. 4b. Upper and lower arrows, B). The blood vessels are engorged with RBCs.
Sequence confirmation of the CMLV PCR products: The PCR products were sequenced to confirm the origin of the sequence amplified. The control OPV DNA amplified was shown to be an unexpected OPV.
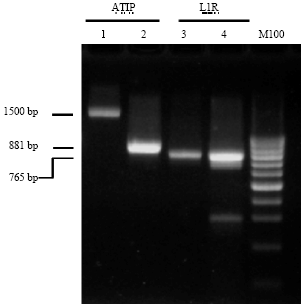 | |
| Fig. 2: | PCR product analysis of CMLV A27L, A33R and B5R orthologs.. A27L, A33R and B5R primers (Hooper et al., 2004) were used to amplify OPV sequences from 1:50 dilutions of DNA extracts of camel skin samples and an OPV vaccine. The 100 bp DNA ladder is the far right of the figure. Lanes 1 and 2: ATIP PCR products for OPV and CMLV, respectively. Lanes 3 and 4: L1R PCR products for OPV and CMLV, respectively. Notice that the OPV PCR product of the ATIP gene was not consistent with that of the Saudi CMLV. L1R gene amplifications produced comparable results for both OPV and CMLV |
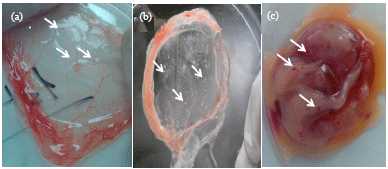 | |
| Fig. 3(a-c): | CMLV isolation of the CAM of SPF ECE. (a) Some embryos (7-10%) developed haemorrhagic pitichae upon inoculation with CMLV. (b) The second passage was incubated for 5 days at 38°C and (c) CMLV suspension prepared according to the standard procedures for virus isolation (OIE, 2009) was inoculated onto the CAM of SPF ECE and incubated for 5 days at 35°C |
That information is currently being held awaiting the approval for publication by the authorities.
The CMLV L1R gene ortholog resembles closely (100%) CMLV M-96 from Kazakhstan (GenBank accession No. AF438165.1) (Fig. 5). The CMLV A33R gene ortholog resembles closely (100%) CMLV M-96 from Kazakhstan (GenBank accession No. AF438165.1) (Fig. 6).
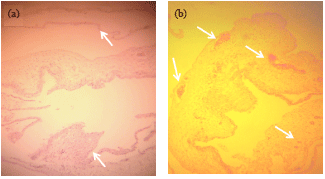 | |
| Fig. 4(a-b): | Histopathological examination of the CAM of SPF ECE inoculated with CMLV. Localized regions of the CAM were thickened (lower arrow, a) compared to the normal areas of the CAM (Upper arrow, a). In membranes with visible haemorrhagic pock lesions, there is evidence of an increase in small blood vessel supply to the membrane (upper and lower arrows, b). The blood vessels are engorged with RBCs |
 | |
| Fig. 5: | Initial BLAST analysis of CMLV L1R ortholog. The sequence obtained was 100% homologous to known CMLV sequence M-96 form Asia |
The CMLV B5R gene ortholog resembles closely (99%) CMLV M-96 from Kazakhstan (GenBank accession No. AF438165.1) (Fig. 7). Two gaps may have contributed to this homology percent. Review of the sequence is required to confirm that the gaps were not due to sequencing errors. The CMLV A27L gene ortholog resembles closely (98%) CMLV M-96 from Kazakhstan (GenBank accession No. AF438165.1) (Fig. 8). Review of the sequence is required to confirm that the two gaps observed were not due to sequencing errors. The CMLV ATIP gene resembles closely (99%) CMLV M-96 from Kazakhstan (GenBank accession No. AF438165.1). Part of the obtained sequence was submitted to GenBank (GenBank accession No. GU937751) (Fig. 9).
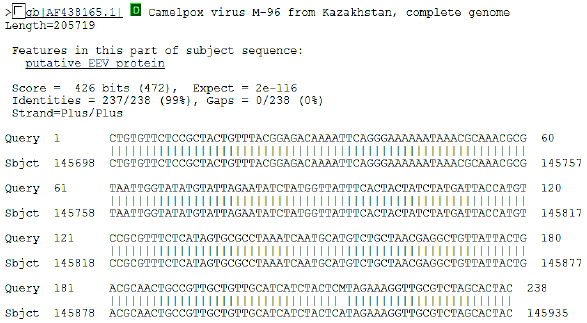 | |
| Fig. 6: | Initial BLAST analysis of CMLV A33R ortholog. The sequence obtained was 100% homologous to known CMLV sequence M-96 form Asia |
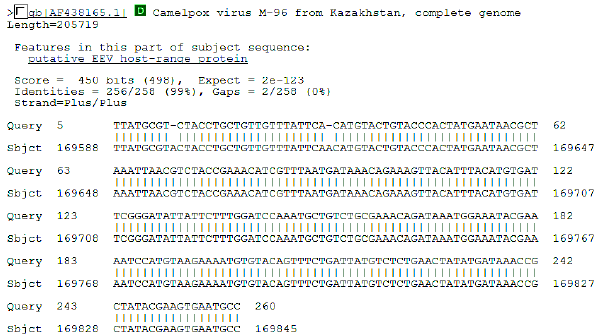 | |
| Fig. 7: | Initial BLAST analysis of CMLV B5R ortholog. The sequence obtained was 99% homologous to known CMLV sequence M-96 form Asia |
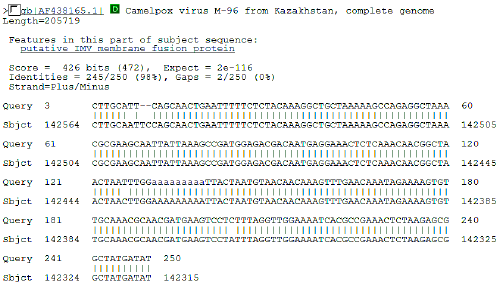 | |
| Fig. 8: | Initial BLAST analysis of CMLV A27L ortholog. The sequence obtained was 98% homologous to known CMLV sequence M-96 form Asia |
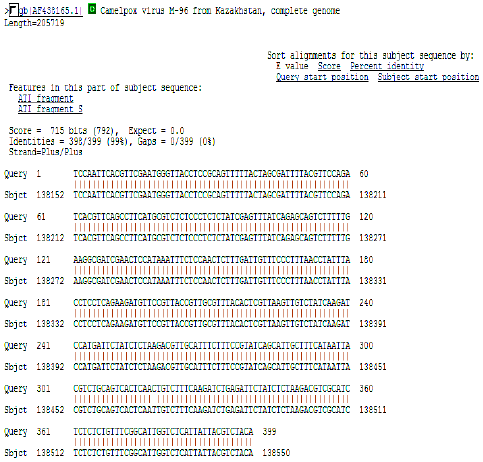 | |
| Fig. 9: | Initial BLAST analysis of CMLV ATIP. The sequence obtained was 99% homologous to known CMLV sequence M-96 form Asia |
DISCUSSION
Poxviridae is a very large family of dsDNA-containing viruses that infect mammals, birds and insects. Eleven genera are recognized, eight of which infect vertebrates and three of which infect invertebrates. Viruses that infect vertebrates are classified in the subfamily Chordopoxvirinae (Carter et al., 2005). Potentially pathogenic orthopoxviruses, including feral versions of vaccinia virus vaccine, persist in nature and re-emerge for reasons we do not fully understand. CMLVV is the most closely related to Variola virus (VARV), sharing all genes involved in basic replicative functions and the majority of genes involved in other host-related functions (Gubser and Smith, 2002).
Although, CMLV has been known for millennia in the Arabian peninsula, no real attempts have been made to characterize this virus. On record, only State Research Center of Virology and Biotechnology Vector, Koltsovo, Novosibirsk Region, former Russia and National Center for Infectious Diseases, Centers for Disease Control and Prevention, Atlanta, GA, USA, have ever attempted to extremely limited sequencing attempts of early isolates from the region (GenBank Acc. numbers U87839, GU937751.1, AY902251.1, AY315878.1, AY102984.1, Y299082.1 and AF375082.1). Apparently, their work was for biological war-fare purposes during the cold-war era.
This attempt for the molecular characterization of Al-Hassa isolate of CMLV was initiated by the possibility that this isolate may be suitable for production of a highly efficient local vaccine. The camel herd from which this virus was isolated had mild skin lesions and did not show the severe generalized form of CMLP that had infected camel herds less than 2 Km away. This meant that the low virulence strain can be used in vaccine-based control programs. This slow- spreading mild strain of the virus has probably been circulating for decades (Abu Elzein et al., 1999).
Rapid confirmation of OPV outbreaks in frontier posts and quarantine facilities is an essential component of national and regional preparedness plans. PCR-based diagnosis has proven superior to other techniques in terms of sensitivity and speed (Mercer et al., 2007). Skin lesions are considered the best source of specimens for laboratory diagnosis of poxvirus infections (OIE, 2009).
We have used a new extraction technique, the FNES technique developed in our lab, to produce DNA for this molecular characterization experiment. The PCR amplicons reported for CAMP ATIP gene using the primers used in this study was 881 bp (Meyer et al., 1994; Ali et al., 2009) indicating that the technology is valid for molecular epidemiological investigations and well as simple diagnostic reactions.
The PCR experiments have also characterized the amplification reactions for a multi-gene PCR panel that can be used for characterize viruses isolated from outbreaks in the region and around the world. This panel was instrumental in the discovery of serious contaminants in one of the animal vaccines.
Preliminary sequence analysis of the PCR products obtained from the CMLV ortholog genes demonstrated that Al-Hassa isolate was indeed highly similar to the Asian Kazakhestan isolate (Afonso et al., 2002).
To our knowledge, this is the first attempt to probe a region that is almost 89000 bp of the enzootic CMLV.
ACKNOWLEDGMENTS
This work was partially funded by the veterinary teaching hospital, College of Veterinary Medicine and Animal Resources and by grant number 110083 of the deanship of scientific research, King Faisal University, KSA. We would like to thank Ahmed Al- Qahtani, Ph.D. Head, Molecular Virology and Infectious Diseases Section, Department of Biological and Medical Research, Research Center, King Faisal Specialist Hospital and Research Center and his research team for providing an excellent sequencing service as a gift for the Central Biotechnology Laboratory, College of Veterinary Medicine and Animal Resources, Bldg. 999, King Faisal University.
REFERENCES
- Abu Elzein, E.M., A.A. Gameel, R.O. Ramadan and F.M. Housawi, 1999. An eruptive moderate form of camelpox infection in dromedary camels (Camelus dromedaries) in Saudi Arabia. Rev. Sci. Tech. Off. Int. Epiz., 18: 749-752.
Direct Link - Afonso, C.L., E.R. Tulman, Z. Lu, L. Zsak and N.T. Sandybaev et al., 2002. The genome of camelpox virus. Virology, 295: 1-9.
CrossRef - Alhendi, A.B., E.M.E. Abuelzein, A.A. Gameel and M.M. Hassanein, 1994. A slow-spreading mild form of camel pox infection. J. Vet. Med. B, 41: 71-73.
CrossRef - Al-Ziabi, O., H. Nishikawa and H. Meyer, 2007. The first outbreak of Camelpox in Syria. J.Vet. Med. Sci., 69: 541-543.
Direct Link - Balamurugan, V., V. Bhanuprakash, M. Hosamani, K.D. Jayappa, B. Chauhan, G. Venkatesan and R.K. Singh, 2009. A polymerase chain reaction strategy for the diagnosis of camelpox. J. Vet. Diagn. Invest., 21: 231-237.
CrossRefPubMedDirect Link - Bancroft, J., A. Stevens and D. Turner, 1996. Theory and Practice of Histological Techniques. 4th Edn., Churchill, Livingston, New York, London, San Francisco, Tokyo Pages: 740.
CrossRefDirect Link - Essbauer, S., M. Pfeffer and H. Meyer, 2010. Zoonotic poxviruses. Vet. Microbiol., 140: 229-236.
CrossRef - Gubser, C. and G.L. Smith, 2002. The sequence of Camelpox virus shows it is most closely related to variola virus, the cause of smallpox. J. Gen. Virol., 83: 855-872.
Direct Link - Hosaka, K., 2004. An easy, rapid and inexpensive DNA extraction method, one-minute DNA extraction for PCR in potato. Am. J. Potato Res., 81: 17-19.
CrossRef - Meyer, H., M. Pfeffer and H.J. Rziha, 1994. Sequence alterations within and downstream of the A-type inclusion protein genes allow differentiation of Orthopoxvirus species by polymerase chain reaction. J. Gen. Virol., 75: 1975-1981.
Direct Link - Panning, M., M. Asper, S. Kramme, H. Schmitz and C. Drosten, 2004. Rapid detection and differentiation of human pathogenic Orthopoxviruses by a fluorescence resonance energy transfer real-time PCR assay. Clin. Chem., 50: 702-708.
Direct Link - Wernery, U., H. Meyer and M. Pfeffer, 1997. Camel pox in the United Arab Emirates and its prevention. J. Camel Pract. Res., 4: 135-139.
Direct Link - Hooper, J.W., E. Thompson, C. Wilhelmsen, M. Zimmerman and M. Ait Ichou et al., 2004. Smallpox DNA vaccine protects nonhuman primates against lethal monkeypox. J. Virol., 78: 4433-4443.
CrossRefDirect Link - Thompson, J.D., D.G. Higgins and T.J. Gibson, 1994. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res., 22: 4673-4680.
CrossRefPubMedDirect Link - Altschul, S.F., T.L. Madden, A.A. Schaffer, J. Zhang, Z. Zhang, W. Miller and D.J. Lipman, 1997. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucl. Acids Res., 25: 3389-3402.
CrossRefPubMedDirect Link - Dafalla, M.A.O. and A.M.A. Elfadel, 2007. Epidemiologic and clinical features of camelpox in Jazan Region, Saudi Arabia. Vet. Res., 1: 66-67.
Direct Link - Ali, H.M.S., A.I. Khalafalla and A.H. Nimir, 2009. Detection o camelpox and vaccinia viruses by polymerase chain reaction. Trop. Anim. Health Prod., 41: 1637-1641.
CrossRef