
W. H. Mady
Department of Poultry Diseases, Animal Health Research Institute, Agriculture Research Center, 12611, Dokki, Giza, Egypt
A. A. Sanousi
Department of Poultry Diseases, Animal Health Research Institute, Agriculture Research Center, 12611, Dokki, Giza, Egypt
H. M. Amer
Department of Poultry Diseases, Animal Health Research Institute, Agriculture Research Center, 12611, Dokki, Giza, Egypt
I. M. El-Sabagh
Department of Virology, Faculty of Veterinary Medicine, Cairo University, 12211 Giza, Egypt
A. M. Khafagy
Department of Poultry Diseases, Animal Health Research Institute, Agriculture Research Center, 12611, Dokki, Giza, Egypt
M. S. Saber
Department of Virology, Faculty of Veterinary Medicine, Cairo University, 12211 Giza, Egypt
International Journal of Virology
Year: 2010 | Volume: 6 | Issue: 2 | Page No.: 73-81







ABSTRACT
In mid-February 2006, an outbreak of Highly Pathogenic Avian Influenza (HPAI) H5N1 affected the commercial poultry production sector and backyards in Egypt and resulted in magnificent socioeconomic losses. The present study was designed for detection, isolation and characterization of Avian Influenza Viruses (AIV) circulating among different poultry species during the first year of the outbreak. A total of 22 commercial poultry farms (16 chicken, 5 ducks and 1 quails) and 5 zoo birds were examined for such purpose. While, no influenza viruses were detected in zoo birds, 68% of the commercial poultry farms located in seven Egyptian governorates were positive to AIV and 53.3% of them were subtyped as H5. Virus isolation in SPF chicken embryos was successful for nine samples from the first egg passage. Molecular characterization of the viral RNA, in the harvested allantoic fluid by multiplex RT-PCR, confirmed the virus identity. On the basis of the OIE criteria of pathogenicity and the observed mortality profile, the AIV isolated during the first wave of the outbreak was identified as highly pathogenic, where the Intravenous pathogenicity index (IVIP) score was 2.83 and the mortality rate was 100%. In conclusion, this study outlines the major implication of subtype H5 HPAI in field affections during the first year of the outbreak. Additional involvement of other influenza A viruses and probably haemagglutinating respiratory agents can not be excluded.
PDF Abstract XML References Citation
How to cite this article
W. H. Mady, A. A. Sanousi, H. M. Amer, I. M. El-Sabagh, A. M. Khafagy and M. S. Saber, 2010. First Year of the Highly Pathogenic Avian Influenza H5N1 Outbreak in Egypt: Rapid Antigenic/Molecular Diagnosis and Virus Isolation. International Journal of Virology, 6: 73-81.
DOI: 10.3923/ijv.2010.73.81
URL: https://scialert.net/abstract/?doi=ijv.2010.73.81
DOI: 10.3923/ijv.2010.73.81
URL: https://scialert.net/abstract/?doi=ijv.2010.73.81
INTRODUCTION
Influenza A viruses are negative-sense, single-stranded enveloped RNA viruses with a segmented genome belonging to the family Orthomyxoviridae (Cox et al., 2000). On the antigenic bases of their surface glycoproteins; particularly hemagglutinin (HA) and neuraminidase (NA), influenza A viruses are divided into distinct subtypes. To date, sixteen HA (H1-H16) and nine NA (N1-N9) subtypes, which are found in many different combinations have been identified (Horimoto and Kawaoka, 2005).
Influenza A viruses have been isolated from a wide variety of hosts, including more than 90 bird species and various mammalian species, such as humans, pigs, horses, cats, minks and marine mammals. However, ecological studies suggest that their natural reservoirs are wild aquatic birds belonging to the orders Anseriformes (ducks, geese and swan species) and Charadriiformes (shorebird species, including gulls) (Webster et al., 1992).
Avian influenza viruses of all 16 subtypes can cause Low Pathogenicity Avian Influenza (LPAI) in susceptible birds, with mild respiratory symptoms and low mortality rates. However, in some cases, this infection may cause significant mortality rates, generally when it occurs in a combination with other bacterial or viral infections. Highly Pathogenic Avian Influenza (HPAI) is a systemic disease with high mortality rates approaching 100% in chickens and Turkeys. This disease is restricted to strains belonging to H5 and H7 subtypes (Johnson and Maxfield, 1976; Alexander and Gough, 1986; Alexander, 2000).
The outbreaks of HPAI H5N1 viruses have been recognized in Asia since 1996, when a fulminating and rapidly fatal systemic disease appeared and spread massively in domestic poultry (Shortridge, 1999). Although, the poultry depopulation policy adopted at that time had been succeeded in controlling the virus spread, it was not sufficient to prevent frequent episodes of the disease outbreaks in several Asian, African and European countries over the following few years. The current outbreak of HPAI H5N1 affected an unprecedented number of countries (No. = 61) extending over a wide geographical zone (OIE reports, 2008).
In mid-February, 2006 a devastating HPAI H5N1 infected the commercial poultry production sectors and backyards in Egypt (Aly et al., 2006 a, b). The outbreak caused great socioeconomic losses in poultry industry (Safwat, 2006) and still considered a renewable problem warning with a possible endemic.
Laboratory diagnosis of Influenza A virus infections is routinely performed by isolation and identification of the virus either in Embryonated Chicken Eggs (ECE) or in cell culture followed by typing the isolated virus by haemagglutination-inhibition test and/or RT-PCR. Virus pathogenicity is determined by estimating the intravenous pathogenicity index or by sequencing the multiple basic amino acids located at the cleavage site of haemagglutinin protein (OIE Manual, 2005).
In this study, we report the isolation and identification of AIV from the tracheal and cloacal swabs of infected chickens, ducks and quails located in seven different Egyptian governorates during the first year of the HPAI H5N1 outbreak.
MATERIALS AND METHODS
Specimens
Tracheal and cloacal swab specimens were collected from suspected cases of Avian Influenza in 22 different commercial poultry flocks located in seven Egyptian governorates (Sharkia, Giza, Dakahlia, Gharbia, Menofia, Ismailia and Menia) as well as from 5 flocks in Giza Zoo between October 2005 and March 2007. Affected birds included chickens (broilers, layers and broiler breeders); ducks (parents and meat type); Quails (parents) and zoo birds (White and black chickens, white and colored ducks and swans). Cotton swabs were used for specimens' collection and were subsequently immersed in 2 mL phosphate buffered saline, pH 7.2 containing 4000 IU penicillin, 400 μg streptomycin and 1000 IU mycostatin. The swab suspension was clarified by centrifugation at 2000 rpm for 10 min and the supernatant was stored at -20°C till use.
Rapid Antigen Detection Tests for Typing and Subtyping
Chromatographic immunoassay test kits for influenza type A and subtype H5 antigens (Anigen, Korea) were used for rapid detection and subtyping of avian influenza viruses in swab specimens (Gavin and Thomson, 2003). The test device combines a sample well with a reading well that contains a membrane strip for influenza A or H5 viral antigens. The tests were performed according to the manufacturer's instructions where, each sample was mixed with a specimen diluent containing buffered saline, detergent, mucolytic agent and preservative. Then, 200 μL were transferred by pipette into the middle of the test well of the device. A positive test was indicated by two color bands in the reading well, one in the Test (T) region and one in the Control (C) region. A negative test was indicated by only one color band in the C region. The absence of any color bands in the T or C regions represented an invalid test. Test readings were performed and recorded after 20-30 min of incubation.
Virus Isolation
The supernatant fluid of each swab suspension was inoculated into the allantoic cavity of five 9-11 days-old embryonating Specific-Pathogen-Free (SPF) chicken eggs. The eggs were incubated at 33-34°C for 4-7 days and were examined daily. Eggs containing dying embryos after 24 h of incubation and those remaining alive till the end of incubation period were chilled to 4°C and the allantoic fluid was harvested for further evaluation by haemagglutination test. Fluids that produced negative haemagglutination results passaged two further times before considered negative for virus isolation (OIE Manual, 2005).
Haemagglutination Test
Twenty-five microliters of PBS were added to all wells of a plastic V-bottomed microtiter plate. Twenty-five microliters of the infected allantoic fluid samples were added to the first wells and diluted serially two-fold across the plate. After dispensing, a further 25 μL of PBS to all wells of the plate, 25 μL of 1% (v/v) washed chicken RBCs were added. The plate was incubated at room temperature for 40 min by which the control RBCs usually settles to a distinct button. The HA titers of allantoic fluid samples were expressed as the reciprocal of the highest antigen dilution showing complete haemagglutination (Hierholzer et al., 1969).
RNA Extraction
Extraction of the total RNA from infected allantoic fluid samples was performed using TRIzol reagent (Invitrogen, San Diego, CA) according to the manufacturer's recommendations. Briefly, 400 μL of the allantoic fluid were mixed with 1 mL of the TRIzol reagent and incubated at Room Temperature (RT) for 10 min. After addition of 200 μL Chloroform, the tube was shacked vigorously before reincubated at RT for 15 min. Centrifugation of the tube at 14,000 rpm for 15 min at 4°C resulted in separation of the mixture into 3 phases. The upper phase containing the RNA was transferred to another tube, where 500 μL of isopropanol were added and the tube was incubated at RT for 10 min. After another cycle of centrifugation at 14,000 rpm for 10 min, RNA precipitated in the form of gel-like pellet. RNA was washed by ethanol 75%, air-dried for 10 min and dissolved in 30 μL nuclease-free water.
First Strand (cDNA) Synthesis
Reverse Transcription (RT) of the total RNA extract was carried out using RevertAidTM first strand cDNA synthesis kit (Fermentas, Germany). A reaction mixture of 5 μL RNA extract, 200 ng random hexamer primers and 6 μL nuclease-free water was prepared and incubated at 70°C for 5 min. The mixture was placed on ice for 5 min; then 4 μL of 5x reaction buffer; 1 μL of RNase inhibitor; 2 μL of 10 mM dNTP mix and 1 μL of MMuLV reverse transcriptase were added. The RT reaction was conducted in a thermal cycler (Gene-Amp 9700, Applied Biosystem Inc., CA) for one cycle at 25°C for 10 min; 42°C for 60 min and 70°C for 10 min. The synthesized cDNA was used for PCR amplification of influenza specific genes.
| Table 1: | Oligonucleotide primers for multiplex PCR |
 | |
Multiplex PCR
The PCR amplification of type A and H5 subtype AIV-specific sequences was optimized and performed using the oligonucleotide primers shown in Table 1. Multiplex PCR was carried out in a 50 μL reaction volume containing 1x PCR reaction mixture (Fermentas, Germany); 0.2 μM of each M-specific primer; 0.4 μM of each H5-specific primer and 5 μL of cDNA. The used thermal cycling profile was initial denaturation for 5 min at 94°C followed by 35 cycles of 94°C for 30 sec; 50°C for 30 sec; and 72°C for 1 min with a final extension of 10 min at 72°C. The PCR products were resolved by electrophoresis in 1.2% agarose gel stained with ethidium bromide.
Intravenous Pathogenicity Index (IVPI) Test
The test was performed as described by OIE Manual (2005), where fresh infective allantoic fluid samples of 4 log2 HA titers were diluted 1:10 in PBS. Ten eight-weeks-old SPF chickens were injected intravenously with 0.2 mL of the diluted allantoic fluid. Birds were examined for clinical signs and death at 24 h intervals for 10 days. At each observation, each bird is scored 0 if normal; (1) if sick (showing a single clinical sign); (2) if severely sick (showing multiple signs) and (3) if dead. The IVPI is the mean score per bird per observation over the 10 days period. An index of 3.00 means that all birds died within 24 h and an index of 0.00 means that no bird showed any clinical signs during the 10 days period. In general, any influenza virus regardless of the subtype, giving a value greater than 1.2 is considered to be highly pathogenic.
RESULTS
Rapid Detection and Subtyping of Avian Influenza Viruses
All the pooled tracheal and cloacal swab specimens, that represent 22 different poultry flocks (16 chickens, 5 ducks and 1 Quail) and 5 Zoo birds, were tested using the rapid Avian Influenza A antigen detection strips. With the exception of Zoo birds that was proved negative, the different tested domestic poultry species developed positive reactions in 13/16 (81%) of the chicken flocks, 1/4 (25%) of the duck flocks and 1/1 (100%) of the quail flocks. The affected flocks were distributed in 6 different Egyptian governorates including: Gharbia 2/2 (100%), Menia 2/2 (100%); Ismailia 1/1 (100%); Menofia 1/1 (100%); Giza 8/12 (66.7%) and Sharkia 1/3 (33.3%).
Only positive samples were subtyped as H5 using the rapid H5 antigen detection strip tests. While, 7 out of 13 of the chicken flocks (53.8%) and the sole quail flock reacted positively, the only AI-positive duck flock was not an H5 influenza virus (Table 2).
| Table 2: | A Collective data on the test samples and their reactivity in the different assays performed |
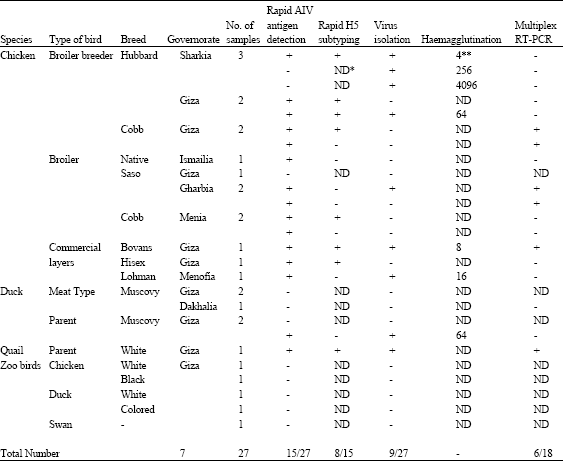 | |
| *ND: Not done, **Haemagglutination unit/25μL of the virus suspension | |
Isolation of Avian Influenza Viruses in Embryonated Chicken Eggs
The different collected samples were propagated in the allantoic cavity of SPF chicken embryos for virus isolation. Only 9 samples (7 from chickens, one from ducks and one from quails) could be isolated. Most of the isolated samples were recovered from the first egg passage. The isolated virus identity was further confirmed by the hae magglutination (HA) test, which showed variable HA titers ranging from 4 log2 to 12 log2 (Table 2).
Simultaneous Identification and Subtyping of the Isolated Viruses Using Multiplex RT-PCR
A multiplex RT-PCR assay was optimized for use in detection and subtyping of the H5N1 strains of Avian influenza virus. The oligonucleotide primers were chosen for amplification of fragments of the M and H5 gene sequences; analyzed and optimized in a standard reaction. The test was applied on 18 samples showed positive reactivity either in antigen detection or in virus isolation. Only six samples proved their identity as H5 virus strains by amplification of 2 specific bands of 244 and 456 bp (Table 2, Fig. 1).
Pathogenicity Testing
Application of IVPI test on ten eight-weeks-old SPF chicken showed that the isolated viruses have a mean score of 2.83 (Table 3). This result along with the death of all inoculated chicken confirmed the high pathogenicity nature of the isolated H5 influenza A viruses.
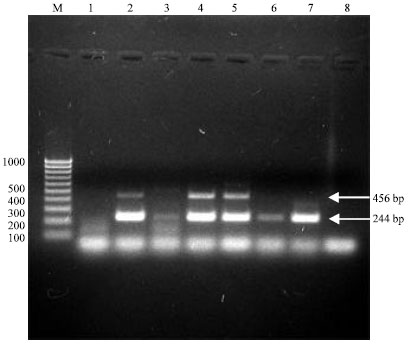 | |
| Fig. 1: | Multiplex RT-PCR products of selected samples for detection of HPAI H5N1 viruses as shown in ethidium bromide stained agarose gel electrophoresis: Lane M represents 100 bp DNA molecular weight ladder; Lanes 3 and 6 show positive amplification of a 244 bp fragment of M gene only; Lanes 2, 4 and 5 show positive amplification of 244 and 456 bp fragments of the M and H5 genes, respectively; Lane 7 represents a standard strain of Avian Influenza H9N2 and Lane 1 and 8 represents negative samples with no amplification observed |
| Table 3: | Intravenous pathogenicity index (IVPI) scores throughout the test period |
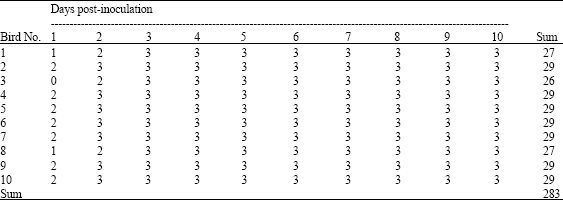 | |
| Scoring; 0: Normal, 1: Sick 2: Very sick (more than one sign) 3: Dead. IVPI: Sum/Total = 283/100 = 2.83 | |
DISCUSSION
Influenza A viruses containing HA of subtypes H5 and H7 are well known as highly pathogenic viral agents causing severe outbreaks in poultry worldwide (Alexander and Gough, 1986; Alexander, 2000). During the last few months of 2005, there was a rising anticipation of an H5N1 attack towards the Egyptian poultry industry following by a similar situation occurred in Turkey and Romania (Nicoll, 2005). At mid-February 2006, the expectations were changed into a reality, when the first isolates of highly pathogenic Avian Influenza H5N1 viruses were recovered from backyard birds. Soon, after the poultry farms of intensive breeding systems were affected (Aly et al., 2006a, b). By the end of March 2006, twenty Egyptian governorates officially reported the occurrence of HPAI H5N1 infection inside their range, a matter that caused great socioeconomic losses in poultry industry (Kilany, 2006; Safwat, 2006).
This study was started so early at October 2005, five months before the announcement of outbreak in Egypt and extended till March 2007 to cover the entire first year of the outbreak. It planned for detection, isolation and characterization of avian influenza viruses among domestic and wild birds in different Egyptian governorates and determination of their pathogenicity. During the course of the study, a total of 22 commercial poultry flocks (including chicken, ducks and quails), located into seven Egyptian governorates, were examined. All flocks showed either significant drop in production and/or high mortality rates. In addition, five wild birds located at Giza Zoo, suddenly died with no clinical symptoms, were also included in the study. Only tracheal or cloacal swabs were collected and used for virological examination.
Detection of influenza A viral antigens in clinical specimens by rapid immunochromatographic assays are widely used for diagnosis and typing of avian influenza viruses; because of their simplicity and ability for rapid diagnosis. However, the usefulness of such assays seems somewhat limited due to their low sensitivity. It is generally recommended that the negative reactions should be confirmed by virus isolation and/or RT-PCR (Gavin and Thomson, 2003; Peiris et al., 2004). In the current study, all the collected samples were tested using a rapid influenza A antigen detection kit. The positive samples were subtyped using a parallel H5 detection kit. Only, 15/27 (55.6%) of the samples were identified as influenza A viruses, from which 8 samples (53.3%) were subtyped as H5 (Table 2). These levels of infection justify the role of avian influenza viruses, particularly H5N1, in the field outbreaks among poultry flocks in Egypt during the period of study. Besides, the results of rapid antigen test kits indicate a sufficient level of detection for the positive cases and their ability to give fast response to the potential field problems (Davison et al., 1998). Further testing of the negative samples, by virus isolation and haemagglutination test, identified two more samples that are possibly containing avian influenza virus or a similar respiratory virus of potential haemagglutinating activity (e.g., Newcastle disease virus).
Virus isolation still remains the gold standard of diagnosis and indispensable for virus characterization (De Jong and Hien, 2006). In this study, avian influenza viruses were successfully recovered from 9 tested samples (four of H5 subtype and five of other subtypes). Only 4 samples failed in the isolation process even after 3 successive blind passages. This lower percentage of positivity comparing with the rapid antigen detection may demonstrate a degree of false positive reactivity in antigen detection assays, or due to the presence of inactivated viral antigens in such samples. Titration of the isolated viruses by haemagglutination test exhibited a widely variable titers ranging from 4 log2 to 12 log2 that reflect a distinct difference of viral load in the original inoculums.
In the last few years, RT-PCR was established as the most accurate and reliable assay for detection of H5N1 in clinical samples (Payungporn et al., 2004; Tran et al., 2004; Ng et al., 2006). This test combines the speed and simplicity of rapid antigen detection assays with the higher sensitivity and specificity of virus isolation. In this context, a one-step multiplex RT-PCR was developed and optimized for specific detection of H5 avian influenza viruses in the allantoic fluid collected from inoculated eggs. Unexpectedly, lower range of positive samples (6/18) was detected by the developed assay, which may indicate the necessity of further validation. However, several samples were detected by the multiplex RT-PCR although, they did not show any reactivity in antigen detection assays and/or virus isolation. Such degree of specificity and sensitivity promises with encouraging outputs of such assay, after complete validation, in rapid and accurate diagnosis of field outbreaks.
Pathogenicity assessment of the isolated avian influenza virus strains is usually indicated by the Intravenous pathogenicity index (IVIP), according to the OIE Manual (2005) and by the induced mortality level. To date, all HPAI viruses generate IVPI scores greater than 1.2 and induce mortalities exceeding 75%. In the current study, the recovered viruses showed clinical signs, gross lesions and death of 10 out of 10 of the inoculated chickens with a calculated IVPI of 2.83 during the observation period (Table 3). Based on this assessment, avian influenza virus isolates, obtained from infected poultry in Egypt during this study, are highly pathogenic.
CONCLUSION
Different virological tests were utilized for detection and characterization of avian influenza viruses that were circulating among poultry population in seven Egyptian governorates during the first year of the outbreak. These tests lead to an affirmed diagnosis of H5 HPAI in 50% of tested samples with a subsidiary existence of other subtypes of influenza A and probably respiratory haemagglutinating viruses. Therefore, the use of multiple diagnostic approaches is strongly recommended for an accurate and definite diagnosis of HPAI especially during the periods of the outbreak.
REFERENCES
- Alexander, D.J., 2000. A review of avian influenza in different bird species. Vet. Microbiol., 74: 3-13.
CrossRefPubMedDirect Link - Alexander, D.J. and R.E. Gough, 1986. Isolations of avian influenza virus from birds in Great Britain. Vet. Rec., 118: 537-538.
PubMed - Davison, S., A.F. Ziegler and R.J. Eckroade, 1998. Comparison of an antigen-capture enzyme immunoassay with virus isolation for avian influenza from field samples. Avian Dis., 42: 791-795.
PubMed - De Jong, M. and T.T. Hien, 2006. Avian influenza A (H5N1): A review. J. Clin. Virol., 35: 2-13.
CrossRefPubMedDirect Link - Hierholzer, J.D., M.T. Suggs and E.C. Hall, 1969. Standardized viral haemagglutination-inhibition test. II. Description and statistical evaluation. Applied Microbiol., 18: 824-833.
Direct Link - Horimoto, T. and Y. Kawaoka, 2005. Influenza: Lessons from past pandemics, warnings from current incidents. Nat. Rev. Microbiol., 3: 591-600.
CrossRefPubMedDirect Link - Johnson, D.C. and B.G. Maxfield, 1976. Occurrence of avian influenza virus infection in laying chickens. Avian Dis., 20: 422-424.
PubMed - Ng, L.F., I. Barr, T. Nguyen, S.M. Noor and R.S. Tan et al., 2006. Specific detection of H5N1 avian influenza A virus in field specimens by a one-step RT-PCR assay. BMC Infect. Dis., 6: 40-40.
CrossRefDirect Link - Nicoll, A., 2005. Avian influenza detected in Turkey and Romania. Euro Surveill, 10: 2807-2807.
PubMedDirect Link - Payungporn, S., P. Phakdeewirot, S. Chutinimitkul, A. Theamboonlers and J. Keawcharoen et al., 2004. Single-step multiplex reverse transcription-polymerase chain reaction (RT-PCR) for influenza A virus subtype H5N1 detection. Viral Immunol., 17: 588-593.
CrossRefDirect Link - Peiris. J.S., W.C. Yu, C.W. Leung, C.Y. Cheung and W.F. Ng et al., 2004. Re-emergence of fatal human influenza A subtype H5N1 disease. Lancet, 363: 617-619.
CrossRefDirect Link - Fouchier, R.A., T.M. Bestebroer, S. Herfst, L. Van Der Kemp, G. F. Rimmelzwaan and A.D. Osterhau, 2000. Detection of influenza A viruses from different species by PCR amplification of conserved sequences in the matrix gene. J. Clin. Microbiol., 38: 4096-4101.
Direct Link - Shortridge, K.F., 1999. Poultry and the influenza H5N1 outbreak in Hong Kong, 1997: Abridged chronology and virus isolation. Vaccine, 17: S26-S29.
PubMed - Tran, T.H., T.L. Nguyen, T.D. Nguyen, T.S. Luong and P.M. Pham et al., 2004. Avian influenza A (H5N1) in 10 patients in Vietnam. N. Engl. J. Med., 350: 1179-1188.
Direct Link - Webster, R.G., W.J. Bean, O.T. Gorman, T.M. Chambers and Y. Kawaoka, 1992. Evolution and ecology of influenza A viruses. Microbiol. Rev., 56: 152-179.
PubMedDirect Link