
A. Sinha
Department of Mycology and Plant Pathology,Institute of Agriculture Sciences, B.H.U., Varanasi-221005, India
M. Srivastava
Department of Botany, Harish Chandra P.G. College Varanasi-221001, India
International Journal of Virology
Year: 2010 | Volume: 6 | Issue: 3 | Page No.: 150-157







ABSTRACT
In present study effect of Mungbean yellow mosaic virus on the chlorophyll, total nitrogen, protein, total phosphorous and carbohydrate content in three varieties of mungbean plants HUM -2 (Malviya Jagriti), ML-192, Pusa Baisakhi were studied. Total chlorophyll, chlorophyll a, chlorophyll b content and carbohydrate content were lower in virus infected mungbean plant varieties. Total nitrogen, protein increased and total phosphorous content was found to be high in virus infected mungbean leaves. Estimation of chlorophyll in healthy and diseased leaves of mungbean plants at 15, 30, 45, 60 and 75 days were done and absorbance were recorded by spectrophotometer at 663 and 645 nm. Total nitrogen of leaves was measured by modified microkjekldahl method and total protein content was determined by multiplying the total nitrogen content with 6.25. Total phosphorous were estimated by digestion with Tri-acid mixture and reading was taken in spectrophotometer at 470 nm. Determination of carbohydrates were done by total water soluble sugars and starch content.
PDF Abstract XML References Citation
Received: June 08, 2010;
Accepted: July 23, 2010;
Published: September 07, 2010
How to cite this article
A. Sinha and M. Srivastava, 2010. Biochemical Changes in Mungbean Plants Infected by Mungbean yellow mosaic virus. International Journal of Virology, 6: 150-157.
DOI: 10.3923/ijv.2010.150.157
URL: https://scialert.net/abstract/?doi=ijv.2010.150.157
DOI: 10.3923/ijv.2010.150.157
URL: https://scialert.net/abstract/?doi=ijv.2010.150.157
INTRODUCTION
Green gram, commonly known as Mungbean or mung is an important pulse crop next to chickpea and pigeon pea in India. The major causes for low productivity are the incidence of insects and diseases. It was reported that Mungbean yellow mosaic virus (MYMV) alone could cause losses to the tune of 30 to 70% in green gram. In present study effect of MYMV on the chlorophyll nitrogen phosphorous, protein, carbohydrate content on three varieties of mungbean plant has been recorded. All various macroscopic and microscopic symptoms of disease must originate in biochemical aberrations induced directly or indirectly by the viruses (Matthews, 1970). Virus diseases provide simplified system for studies on physiology of host pathogen interaction since no metabolic activity has been associated with isolated plant viruses. The MYMV is the most destructive viral disease of mungbean and blackgram in the Indian subcontinent and adjacent areas of Southeast Asia. The MYMV was reported in 1955 in mungbean growing area in IARI, Delhi (Nariani, 1960). The MYMV infects practically all the major kharif pulses (Vigna radiata) (Mungbean) V. mungo (Urd bean ), V. aconotifolia (moth bean), V. lathyroides (Lathyrus), Glycine max (Soybean), Dolichos biflorus (Horse gram), Phaseolu vulgaris (French bean), Cajanus cajan (Pigeon pea) throughout India (Srivastava et al., 2010). The chlorophyll content was studied by Mali et al. (2000) in moth bean infected by Yellow mosaic virus (Gill and Singh, 2000) in mungbean. Various workers have made different prepostitions on the effect of various infections on nitrogen metabolism of the host (Farrag and Ramakrishnan, 1969; Singh and Bhargava, 1984). Total phosphorous content has been studied by Chakraborty (1993) and Chakraborty et al. (1995). Total carbohydrates in virus-infected plants has been reported in Cucumis melo L. leaves infected with Cucumber mosaic virus (Konishi et al., 2004) in Soybean leaves infected by Soybean mosaic virus (Xiaoyan et al., 2000).
MATERIALS AND METHODS
The whole experiment was done at Department of Mycology and Plant Pathology, Institute of Agricultural Sciences, B.H.U., Varanasi, India during January 2009 to June 2009 for studying the chlorophyll, protein, nitrogen, phosphorous and carbohydrate contents of three varieties HUM-2 (Malviya jagriti), ML-192 and Pusa Baisakhi of Mungbean plants were collected at 15, 30, 45, 60 and 75 days after inoculation.
Chlorophyll Content
For estimation of Chlorophyll healthy and diseased leaves of 15, 30, 45, 60 and 75 days were taken. The leaves were washed with distilled water and the water was soaked by butter paper. Then, the leaves were cutted into small pieces with scissors and 100 mg were taken for grinding into mortar and pestle with 80% acetone. The ground solutions were taken into test tubes and the final volume made to 10 mL by adding 80% acetone. The tubes were wrapped with black paper and were kept in refrigerator overnight. Next day the solutions were centrifuged at 5000 rpm for 10 min. The supernatant was taken in clean test tubes separately. The absorbance was recorded at 663 and 645 nm in a spectrophotometer (Bruinsona, 1963). Chlorophyll a, b and total chlorophyll were calculated by using following formula:
Chlorophyll a (mg g-1 tissue) = [12.7 (D663)-2.69 (D645)]xV / 1000xW Chlorophyll b (mg g-1 tissue) = [22.9 (D645) - 4.68 (D663)]xV/1000xW Total Chlorophyll (mg g-1 tissue) = [20.2 (D645) +8.02 (D663)]xV/1000 xW |
Where:
| D | = | Optical density at respective nm |
| V | = | Final volume of chlorophyll extract in 80% acetone |
| W | = | Fresh weight of the tissue extracted |
Estimation of Total Nitrogen and Protein
Primarily, the total nitrogen of leaves was measured by the modified microkjekldahl method (Lang, 1958). For this 100 mg of dried and uniformly powdered leaf sample was digested with 1-2 mL of concentrated H2SO4 (1 g salicylic acid was dissolved in 20 mL of H2SO4) in presence of 1-2 pieces of sodium thiosulphate and about 1-2 mL of 60% perchloric acid (0.05 g copper sulphate was added per 100 mL of perchloric acid). This mixture was heated till to get a clear solution .The solution was made up to 100 mL by adding distilled water. Nitrogen content was determined by Nesselerization and with the help of standard curve of NH4 (SO4)2 total protein content was determined by multiplying the total nitrogen content with 6.25.
Determination of Total Phosphorous
The plant samples were digested by digestion in Tri-acid mixture of concentrated HNO3: HCLO4:H2SO4 = (10:4:1). The phosphorous content of the digest was determined colorimetrically by vandomolybdate yellow colour method described by Jackson (1973). Two gram of 15, 30, 45, 60 and 75 days after inoculation both healthy and virus infected plant material and 15 mL of Tri-acid mixture were taken in digestion tubes. These tubes were placed in one litre beaker containing concentrated H2SO4 (LR grade) and digested on heater. After complete digestion, when the solution became clear white (in some cases, additional amount of HNO3 was added to make the solution clear) the digested material was filtered by No. 1 filter paper through repeated washing with distilled water and the volume was made up to 100 mL of volumetric flask. Ten milliliter of plant extract was taken in 50 mL volumetric flask and 10 mL of vandate molybdate solution was added and the volume was made up to 50 mL with distilled water and reading was taken in spectrophotometer at 470 nm.
For preparing standard P solution, 0.2195 g of potassium dihydrogen phosphate (KH2SO4) was dissolved in 400 mL of distilled water in one litre volumetric flask. Then 25 mL of 7 NH2SO4 was added in the solution and the volume was made up to the mark. The strength of the solution was 50 ppm. Then, 0, 1, 2, 4 and 5 mL of standard solution was taken in 50 mL volumetric flask and added 10 mL of vandate molybdate reagent. The volume was made up to mark with distilled water. Color was developed immediately but the reading was recorded after 10 min in the colorimeter at 470 nm. A standard curve was prepared by reading the standard P solutions. Then the amount of total phosphorus was determined.
Determination of Carbohydrates
Carbohydrate fraction consisted reducing and non-reducing sugars, total sugars and starch contents. Anthrone colorimetric method as described by Snell and Snell (1961) was followed to determine the reducing sugar content. The non-reducing sugars were measured by converting them to reducing sugars by the methods of Somogyi (1952). The sum of amounts of reducing and non-reducing sugars gave the value for total water-soluble sugars. The starch content was determined according to the method described by Snell and Snell (1961).
RESULTS
Results are presented in Table 1 indicate a loss of total chlorophyll, chlorophyll a and chlorophyll b content due to virus infection in mung bean varieties viz., HUM-2 (Malviya) b have been observed in the leaves of all varieties at 75th day of infection. Chlorophyll a ranged from 0.04 to 0.09 in healthy mungbean varities and in diseased it ranged 0.02 to 0.07 mg g-1. Chlorophyll b ranged from 0.04 to 0.09 mg g-1 in healthy mungbean varieties and in diseased it was 0.02 to 0.07 mg g-1. Total chlorophyll in healthy mungbean varieties were 0.09 to 0.18 mg g-1 and in diseased it was 0.03 to 0.12 mg g-1.
The results summarized in Table 2 shows that total nitrogen (N) are higher in the virus-infected plants in all 3 varieties viz., HUM-2 (Malviya Jagriti), ML-192 and Pusa Baisakhi that is it ranged from 1.78 to 2.69 mg/100 mg dry wt. in healthy varieties of mung bean plant and in diseased mungbean plant it was 2.43 to 3.22 mg/100 mg dry wt.
Results presented in Table 3 indicates an increase in total protein in virus infected mungbean plants than their healthy counterparts. In healthy mungbean varieties total protein was 1.11 to 1.59 mg/100 mg dry wt. and in diseased total protein was 1.64 to 1.99 mg/100 mg dry wt.
| Table 1: | Chlorophyll content (mg g-1) in var. HUM-2 (Malviya Jagriti), ML-192, Pusa Baisakhi leaves at different periods of MYMV infection |
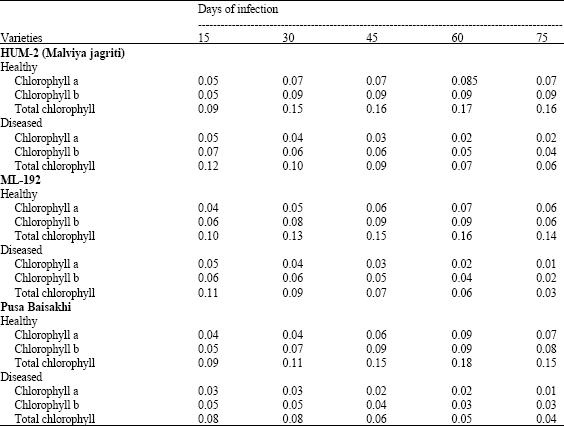 | |
| Table 2: | Effect of MYMV on total nitrogen content (mg 100 mg-1 dry wt.) of mungbean leaves at different periods of MYMV infection |
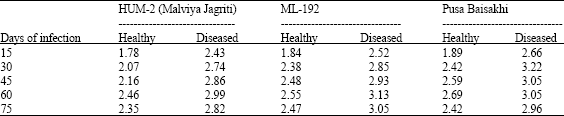 | |
| Table 3: | Effect of MYMV on total protein content (mg 100 mg-1 dry wt.) of mungbean leaves at different periods of MYMV infection |
 | |
An increased protein level in virus-infected plants has also been reported in several cases by previous workers (Erdiller and Ozyaner, 1983; Chakraborty et al., 1995; Shivaprasad et al., 2005; Hofius et al., 2001; Szczepanski and Redolfi, 2008).
The findings summarized in Table 4 revealed that total phosphorus (P) was higher in virus infected mungbean leaves than their healthy counterparts. Total Phosphorous in healthy mungbean ranged from 0.35 to 0.49 mg/100 mg dry wt. and in diseased it was 0.46 to 0.54 mg/100 mg dry wt.
| Table 4: | Effect of MYMV on total phosphorous content (mg 100 mg-1 dry wt.) of mungbean leaves at different periods of MYMV infection |
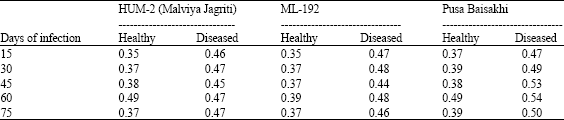 | |
| Table 5: | Total sugars content (mg 100 mg-1 dry wt.) of mungbean leaves at different periods of MYMV infection |
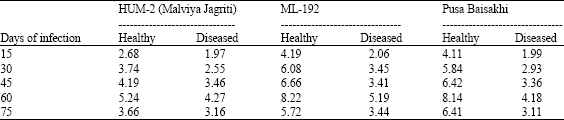 | |
| Table 6: | Starch content (mg 100 mg-1 dry wt.) of mungbean leaves at different periods of MYMV infection |
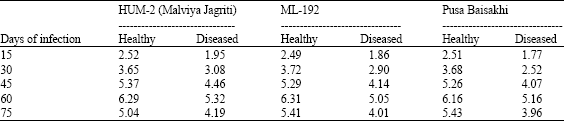 | |
General increases of the total Phosphorus in virus infected plants have been reported for a number of host virus combinations by earlier workers (Chakraborty, 1993; Chakraborty et al., 1995). The above findings clearly indicate that changes in levels of total P contents occur as a result of virus infection.
It is evident from the Table 5 three varieties viz., HUM-2 (Malviya Jagriti), ML-192 and Pusa Baisakhi showed that maximum total sugars were found in leaves of healthy plants which were 5.24, 8.22 and 8.14 mg /1000 mg dry weight on 60th day of inoculation but in MYMV infected leaves it decreased to 4.27, 5.19 and 4.18 mg respectively in all three varieties .The virus infection reduced the total sugars of the plant.
The result presented in Table 6 shows that starch content at 60th day of infection decreased in infected plants. The amount of starch in infected plant were 5.32, 5.05 and 5.16 mg as against 6.29, 6.31 and 6.16 mg in healthy plants of all three varieties, respectively at 60th day. The starch content was always lower in diseased leaves as compared to their healthy counterparts. However, the starch content decreased beyond 60th day. The carbohydrate contents (Total sugars and starch) were reduced in MYMV infected mungbean leaves.
DISCUSSION
Reduction in chlorophyll content has been reported in many host plants infected with different viruses by Pandey and Joshi (1989), Rathore and Agnihotri (1995), Thind et al. (1996), Dantre et al. (1996), Sutha and Rajappan (1998), Mali et al. (2000), Gill and Singh (2000), Milavec et al. (2001), Funayama-Noguchi and Terashima (2006), Pineda et al. (2008), Arora et al. (2009) and Singh and Shukla (2009). There are differences of opinion as to whether the virus destroys the chlorophyll or inhibits its synthesis. According to Sheffield (1933) the virus of tomato acuba mosaic did not affect the chlorophyll in fully developed leaves at the time of infection but did prevent the formation of plastids in young growing leaves. On the other hand, viruses like cucumber mosaic and tomato stripe produce chlorosis-showing destruction of already formed chlorophyll. However, stimulation of chlorophyllase which attack chlorophyll and inhibition of chloroplast development have been reported in virus infected plants. The main reason for increased activity is the disruption of chloroplast in chlorosis-induced tissues.
Concurrent with the present findings a general increase of the total N in virus infected plants have been reported for a number of host virus combinations by Chakraborty (1993), Chakraborty et al. (1995), Thind et al. (1996), Selman and Grant (2008). The main component of total N was probable insoluble protein, free amino acids, nucleic acids and amides. Alteration in any one of the components may disturb the nitrogen metabolism. A few viruses such as tobacco mosaic virus and potato virus protein contributes significantly to the total protein of the host and as a consequence there is an increase in total N. This might be one of the possibilities in the present case too.
The higher percentage seems due to virus multiplication which entails the synthesis of virus specific abnormal protein that accumulates an ultimately raises the percentage over healthy. Increase in protein contents observed in infected mungbean plant leaves may also be correlated with respiration. Increased nitrogen uptake by diseased plants associated with rapid respiration probably helps in the synthesis of more amino acids (Shivaprasad et al., 2005; Hofius et al., 2001; Szczepanski and Redolfi, 2008).
Since, phosphorus compounds have fundamental functions in many metabolic processes, virus induced disturbances of any of these fractions in many metabolic processes, virus induced disturbances of any of these fractions could have far reaching effects on the metabolic machinery of the cell. The rise in the contents of total P suggest that during the course of disease infected plants, a large fraction of inorganic phosphorus was absorbed from the soil and was subsequently converted in to the organic form which was probably incorporated in the synthesis of viral NA (Chakraborty, 1993; Chakraborty et al., 1995). Synthesis of viral NA is primary process of virus replication and its accumulation leads to and increase in the total phosphorous in comparison to healthy plants.
The findings related to carbohydrate are in concurrence with those reported by earlier workers (Ashraf and Zafar, 2000; Goncalves et al., 2005; Handford and Carr, 2007; Adomako and Hutcheon, 2008; Goodman et al., 2008; Singh and Shukla, 2009). Due to virus infection, there is an increased demand for abnormal protein production required for the rapid synthesis of virus particles and for this purpose there has to be increased diversion of assimilated carbon compounds towards protein synthesis resulting in the decreased production of carbohydrates level of plant leaves. The decrease in carbohydrate content may be explained on the basis of possible reduction and (or) faster breakdown of carbohydrates due to increased respiration and conversion of carbohydrates into amino acids, which are ultimately used for protein synthesis. It has been noticed that the various carbohydrate fractions rise till 60th day and later on their level falls in both healthy and MYMV infected plants. This decrease might be due to increased damage to the chlorophyll by increased age of virus infection of both.
REFERENCES
- Adomako, D. and W.V. Hutcheon, 1974. Carbohydrate metabolism and translocation in healthy and cocoa swollen shoot virus-infected cocoa plants. Physiol. Plant., 30: 90-96.
CrossRef - Arora, R., U.N. Joshi, P.P. Gupta and J.V. Singh, 2009. Effect of Yellow mosaic virus on pathogenesis related enzymes and chlorophyll content in mothbean (Vigna aconitifolia). Acta Phytopathologica Entomologica Hungarica, 44: 49-60.
CrossRefDirect Link - Ashraf, M. and Z.U. Zafar, 2000. Patterns of free carbohydrate and starch accumulation in leaves of cotton (Gossypium hirsutum L.) cultivars differing in resistance to cotton leaf curl virus. Arch. Agron. Soil Sci., 45: 1-9.
CrossRef - Chakraborty, S., A. Sinha and B.V.B. Reddy, 1995. Post infection changes in total nitrogen, total phosphorus and protein contents of cucurbit plants affected by cucumber mosaic viruses. Adv. Plant Sci., 8: 417-419.
Direct Link - Dantre, R.K., R.L. Keshwal and M.N. Khare, 1996. Biochemical changes induced by yellow mosaic virus in resistant and susceptible cultivars of soybean (Glycine max (L.) Merrill. Indian J. Virol., 12: 47-49.
Direct Link - Funayama-Noguchi, S. and I. Terashima, 2006. Effects of Eupatorium yellow vein virus infection on photosynthetic rate, chlorophyll content and chloroplast structure in leaves of Eupatorium makinoi during leaf development. Functional Plant Biol., 33: 165-175.
CrossRefDirect Link - Goncalves, M.C., J. Vega, J.G. Oliveira and M.M.A. Gomes, 2005. Sugarcane yellow leaf virus infection leads to alterations in photosynthetic efficiency and carbohydrate accumulation in sugarcane leaves. Fitopatol. Bras., 30: 10-16.
CrossRefDirect Link - Goodman, P.J., M.A. Watson and A.R.C. Hill, 2008. Sugar and fructosan accumulation in virus-infected plants: Rapid testing by circular-paper chromatography. Ann. Applied Biol., 56: 65-72.
CrossRefDirect Link - Handford, M.G. and J.P. Carr, 2007. A defect in carbohydrate metabolism ameliorates symptom severity in virus-infected Arabidopsis thaliana. J. General Virol., 88: 337-341.
PubMed - Hofius, D., K. Herbers, M. Melzer, A. Omid, E. Tacke, S. Wolf and U. Sonnewald, 2001. Evidence for expression level dependent modulation of carbohydrate status and viral resistance by the potato leaf roll virus movement protein in transgenic tobacco plants. Plant J., 28: 529-543.
PubMed - Jackson, M.L., 1973. Soil Chemical Analysis. 1st Edn., Prentice Hall Ltd., New Delhi, India, Pages: 498.
Direct Link - Konishi, T., Y. Ohmiya and T. Hayashi, 2004. Evidence that sucrose loaded into the phloem of a poplar leaf is used directly by sucrose synthase associated with various beta-glucan synthases in the stem. Plant Physiol., 134: 1146-1152.
PubMed - Mali, P.C., U. Burman and S. Lodha, 2000. Effect of planting dates and development of yellow mosaic viruson biochemical constituents of moth bean genotypes. Indian J. Phytopathol., 53: 379-383.
Direct Link - Milavec, M., M. Ravnikar and M. Kovac, 2001. Peroxidases and photosynthetic pigments in susceptible potato infected with potato virus YNTN. Plant Physiol. Biochem., 39: 891-898.
CrossRefDirect Link - Pineda, M., J. Soukupova, K. Matous, L. Nedbal and M. Baron, 2008. Conventional and combinatorial chlorophyll fluorescence imaging of tobamovirus-infected plants. Photosynthetica, 46: 441-451.
CrossRefDirect Link - Rathore, G.S. and J.P. Agnohotri, 1995. Studies of some of the biochemical and physiological components of moth (Vigana aconitifolia [Jacq.] Marechal) affected by yellow mosaic virus (Abstr.). Proceedings of the Global Conference on Advances in Research on Plant Disease and their Management, Feb. 12-17, RCA, Udaipur, India, pp: 99-100.
- Selman, I.W. and S.A. Grant, 2008. Some effects of nitrogen supply on the infection of tomato plants with tomato spotted wilt virus. Ann. Applied Biol., 45: 448-455.
CrossRefDirect Link - Shivaprasad, P.V., R. Akbergenov, D. Trinks, R. Rajeswaran, K. Veluthambi, T. Hohn and M.M. Pooggin, 2005. Promoters, transcripts and regulatory proteins of mungbean yellow mosaic geminivirus. J. Virol., 79: 8149-8163.
CrossRefDirect Link - Sheffield, F.N.L., 1933. The development of assimilatory tissue of solanaceous hosts infected with aucuba mosaic to tomato. Ann. Applied Biol., 20: 57-69.
CrossRef - Thind, S.K., P.K. Monga, N. Kaur and S.S. Cheema, 1996. Analysis of some biochemical and micronutrient constituents of Yellow mosaic virus infected moong. Indian J. Virol., 12: 157-159.
Direct Link - Lang, C.A., 1958. Simple micro determination of Kjeldahl nitrogen in biological materials. Annal. Chem., 30: 1692-1694.
CrossRef