
A.M. Soliman
Plant Pathology Research Institute, Agriculture Research Center, Giza, Egypt
B.N. Barsoum
Department of Chemistry, Faculty of Science, Cairo University, Giza, Egypt
G.G. Mohamed
Department of Chemistry, Faculty of Science, Cairo University, Giza, Egypt
A.A. Rezk
Plant Pathology Research Institute, Agriculture Research Center, Giza, Egypt
A.E. Aboul-Ata
Plant Pathology Research Institute, Agriculture Research Center, Giza, Egypt
H.M. Mazyad
Plant Pathology Research Institute, Agriculture Research Center, Giza, Egypt
International Journal of Virology
Year: 2008 | Volume: 4 | Issue: 1 | Page No.: 14-25







ABSTRACT
This study, aims at determination of efficiency of micro interfering RNA (miRNA) to develop ability of virus resistance against Egyptian PVX isolate (PVX-Eg2) in both potato (Solanum tuberosum L. cv. Spunta) and tobacco (Nicotiana benthamiana). RNA constructs of Sense (PVX-Eg2cpVs), antisense (PVX-Eg2cpCs) and sense/antisense were designed, cloned and sub- cloned for gene transfection using Agrobacterium inoculation technique. Two to 3 leaf-stage seedlings of potato (Solanum tuberosum L. cv. Spunta) and tobacco (Nicotiana benthamiana) were inoculated with the three previous constructs. The construct-treated plants were mechanically inoculated with the PVX-Eg2 isolate. Bioassay and PCR amplification have been able to evaluate transfected-plant resistance against PVX-Eg2 that is caused by siRNA of PVX-Eg2cp. PCR amplification has been able to detect PVX viral genome in all challenged plants those infiltrated with either pFGC5491 vector without insert, or with sense construct and also with antisense construct. Bioassay has confirmed same previous statement. Nine out of 10 sense/antisense-transfected potato plants were negatively reacted with both bioassay and PCR amplification. Same negative reaction has been viewed using both bioassay and PCR for sense/antisense transfected-tobacco plants. Seven out of 10 proved they are PVX-Eg2 resistant.
PDF Abstract XML References Citation
How to cite this article
A.M. Soliman, B.N. Barsoum, G.G. Mohamed, A.A. Rezk, A.E. Aboul-Ata and H.M. Mazyad, 2008. siRNA Silencing of PVX Coat Protein Gene Affects Accumulation of Viral RNA in Potato and Tobacco Plants. International Journal of Virology, 4: 14-25.
DOI: 10.3923/ijv.2008.14.25
URL: https://scialert.net/abstract/?doi=ijv.2008.14.25
DOI: 10.3923/ijv.2008.14.25
URL: https://scialert.net/abstract/?doi=ijv.2008.14.25
INTRODUCTION
Potato (Solanum tuberosum L.) is one of the most important economic crops in Egypt concerning the production, farm income and exportation. It is used for human consumption, animal feed and as a source of starch and alcohol for bio fuels (Horton, 1992). It is affected by more than 20 different viral diseases (Smith, 1931). Potato X potxvirus (PVX) was first recognized on Solanum tuberosum in UK (Smith, 1931). The most important viruses affecting potato crop in Egypt are Potato X potxvirus (PVX), Potato Y potyvirus (PVY) and Potato leaf roll luteovirus (PLRV). These viruses have been shown to cause great economic losses and considered to be the major limiting factors of potato production (El-Amrety, 1970). Potato virus X can be found wherever potatoes are grown in Egypt (El-Amrety, 1970). The yield reduction, caused by this virus, is usually less severe unless PVX occurs together with other viruses (Salazar, 1996). PVX, which has long history in Egypt, was isolated and studied by El-Amrety (1970), Fahmy and Mohamed (1984), Yousef (1992), Abdel-Salam et al. (1994) and Soliman et al. (2000).
PVX-Eg2 isolate was isolated from diseased potato plants in 2006. PVX-like symptoms i.e. mild mosaic, mottling and stunting were shown. Isolated virus was maintained on Nicotiana tabaccum (L.
cv. White Burley) in Department of Plant Virus and Phytoplasma Research, Plant Pathology Research Institute, ARC (Soliman et al., 2006). Nucleotide sequence of a selected fragment of the coat protein gene of the previous isolate was submitted in the GeneBank under the name of PVX (PVX-Eg2) and Accession No. DQ412730 (Soliman et al., 2006).
PVX has a positive-sense genomic RNA consists of five open reading frames (ORFs) numbered 1 to 5. Twenty nucleotides (59-to-39 direction encode polypeptides of 166 kDa (viral replicase), 24 kDa (p24), 12 kDa (p12), 8 kDa (p8) and 25 kDa viral coat protein) (Orman et al., 1990). Post-transcriptional gene silencing (PTGS) is a universal mechanism by which plants are able to switch off the expression of target genes via the reduction of steady state levels of specific RNAs. Double-stranded RNA (dsRNA) can silence genes by developing degradation of homologous RNA in the cytoplasm and by directing methylation of homologous nuclear DNA sequences (Bass, 2000; Hammond et al., 2001; Voinnet, 2001; Waterhouse et al., 2001).
MATERIALS AND METHODS
Design and Synthesis of Oligonucleotide Primers Total RNA was isolated from the PVX-Eg2-infected potato plants, maintained at Dept. of Plant Virus and Phytoplasma Research, Plant Pathology Research Institute, ARC, Egypt in 2002, using RNeasy® Plant Mini Kit obtained from QIAGEN according to manufacturer`s instructions. Pair of primers specific for a part of the coat protein gene of PVX (selected fragment) was designed according to available sequences from the GenBank. The forward primer (PVXcpVs) with XbaI and AscI restriction enzyme sites (underlined) is as follow: 5` TCTAGA GGCGCGCC AGTGGTATGGAACTGGATGC 3`. The complementary primer (PVXcpCs) with BamHI and SwaI restriction enzyme sites (underlined) is as follow: 5`GGATCC ATTTAAAT TCGTTGGATTGTGCCCTGGC 3`. All the restriction sites were added at 5` of the designed primers to facilitate the cloning of the selected domains in the binary vector. The expected size of the RT-PCR product is 259 bp. Total RNA extracted from PVX-Eg2-infected potato plants was used as template for each RT-PCR amplification reactions using one-step RT-PCR reaction kit as instructed from QIAGENTM. PCR-amplified fragments were separated by gel electrophoresis using 1.5% agarose gels in 0.5X TBE buffer, stained with ethidium bromide and visualized with UV illumination using Gel Documentation System.
Cloning and Sequencing of RT-PCR Product (Selected Fragment) RT-PCR product was ligated into PinPoint™ Xa-1 T-Vector (PinPoint™ Xa-1 T-Vector System I, PROMEGA). The recombinant plasmids were introduced into E. coli strain DH5α as described by manufacturer`s instructions. DNA was prepared from selected white colonies, digested with BamHI and fractionated on agarose gels using 1 kb DNA ladder (Promega). The nucleotide sequence of clones having a 259 bp insert were selected for dideoxy sequencing [ABI PRISMTM Dye Terminal Cycle Sequencing Ready Reaction Kit with AmpliTaq DNA Polymerase, FS (Biotechnology Department University of Wisconsin, Madison, USA) and performed on 377 XL automated DNA Sequencer (Applied Biosystem Co.). The nucleotide sequence was compared and analyzed using DNAMAN Sequence Analysis Software (Lynnon BioSoft. Quebec, Canada) with those of PVX isolates available in GenBank.
Sub-Cloning of the Selected Fragment into Binary Vector The selected fragment was cloned into the binary vector pFGC5941 (Fig. 1) (supplied from the University of Arizona, USA) in both orientations once by the SwaI and AscI to give the sense
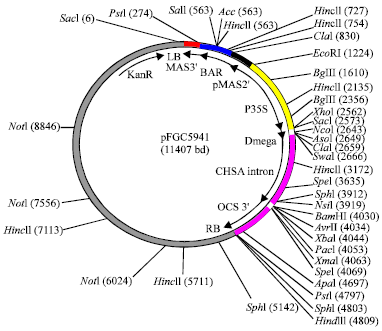 | |
| Fig. 1: | pFGC5941 vector supplied from the University of Arizona, USA. Description of this binary vector could be found at http://www.chromdb.org/5941seq.html |
orientation and another time with BamHI and XbaI to give the antisense orientation leaving the Chalcon Synthase (ChaS) intron in the middle.
Preparation of Sense, Antisense and Sense/Anti-Sense Constructs Prepare the sense construct, 40 μL of DNA plasmid (pPVXcpGS) were digested using 50 U of SwaI (FERMENTAS), 10 μL of 10X buffer O and dH2O was added to final volume of 100 μL. The reaction tube was incubated at 30°C in water bath for 4 h then 50 U of AscI (FERMENTAS) and 10 μL of 10 X buffer Tango™ were added to the reaction tube and incubated at 37°C for 4 h. Prepare the antisense construct, 40 μL of DNA plasmid (pPVXcpGS) were digested using 50 U of BamHI (PROMEGA), 10 μL of 10 X buffer E, 5 μL of BSA (100 μg μL-1), 50 U of XbaI, 10 μL of 10 X buffer D and dH2O was added to final volume of 100 μL. The reaction tube was incubated at 37°C in water bath for 4 h. Prepare the sense/antisense construct, 50 μL of DNA plasmid (pPVXcpGS) were digested using 50 U of each of BamHI and XbaI enzymes, 10 μL of multicore buffer (PROMEGA), 5 μL of BSA (100 μg μL-1) were added and dH2O was added to final volume of 100 μL. The reaction tube was incubated at 37°C in water bath for 4 h. Prepared DNA was electrophoresed in 2% agarose gel prepared in 0.5% TBE buffer, stained with ethidium bromide and visualized by using UV transilluminator. The digested band was cut out with a clean sharp scalpel for gene cleaning. The binary vector pFGC5941 was digested with same two restriction enzymes in each case. One hundred nanogram of digested DNA vector were added to a clean tube and then 17 ng of the digested DNA of selected fragment were added (binary vector and selected fragment were digested with the same restriction enzymes). One microliter of 10 X ligase buffer and 1 U of T4 DNA ligase were added to the ligation mixture. Nuclease-free water was added to the ligation mixture to final volume of 10 μL, the mixture was incubated at 4°C for overnight.
Transformation of the Constructs into E. coli Competent Cells Competent cells E. coli (strain DH5α) were thawed on ice for 30 min and then 100 μL transferred to a pre-chilled 5 mL tube and 5 μL of ligated DNA added and incubated on ice for another 30 min. Cells were heat-shocked for 2 min at 42°C by immersing the tube into a water bath for 2 min and then incubated on ice for 2 min. One milliliter of LB medium was added and cells grown for 1 h at 37°C in shaking water bath. Transformed cells were plated on the surface of prepared plates (LB solid medium with 50 μg mL-1 kanamycin) for overnight at 37°C and bacterial colonies containing recombinant plasmids were selected for further characterization. The enzyme digestion for the DNA minipreps of the selected colonies of constructs (psPVXcpGS [sense], pasPVXcpGS [anti-sense], ps/asPVXcpGS [sense/anti-sense]) were carried out using the restriction enzymes SwaI and AscI (sense), BamHI and XbaI (anti-sense) and AscI and XbaI (sense/anti-sense).
Transformation into Agrobacterium tumefaciensLBA4404 One microgram of plasmid DNA was added to the cells. Freezing the cells in liquid nitrogen was preformed. The cells were thawed by incubating the tubes in a 37°C water bath for 5 min. One milliliter of YEP medium was added to the tube and incubated at 28°C for 2-4 h with gentle shaking. This period allowed the bacteria to express the antibiotic resistance genes. The tubes were centrifuged for 30 sec in a table centrifuge. The supernatant solution was discarded and the cells were resuspended in 0.1 mL YEP medium. The cells were spread on a YEP agar plate containing 50 μg mL-1 kanamycin and 50 μg mL-1 streptomycin. The plate was incubated at 28°C. Transformed colonies should appear in 2-3 days.
Gene Delivery Using the Syringe-Spotting Technique After Agrobacterium tumefaciens LBA4404 has been transformed with vectors harboring different constructs using chemical transformation, 5 mL culture was grown overnight at 28°C in the appropriate antibiotic selection medium. The next day, the culture was inoculated into a 50 mL LB medium containing the selective antibiotics. The culture was grown overnight in a 28°C shaker. Agrobacterium cells were harvested and resuspended in infiltration media [10 mM MgCl2, 10 mM MES (2-N-morpholino ethane sulfonic acid) and 20 μM acetosyringone (4`-Hydroxy-3,5-dimethoxyacetophenone)], adjusted to OD600 of 2.0 and left at room temperature for 3 h. Potato (Solanum tuberosum L. cv. Spunta) and tobacco (Nicotiana benthamiana) plants (10 plants in each treatment) in 2-3 leaf-stage seedlings were infiltrated with Agrobacterium with the constructs psPVXcpGS, pasPVXcpGS, ps/asPVXcpGS separately and pFGC5941 (without construct as a control) using syringe without needle 5 mL syringe (Syringe-Spotting Technique (SST) developed by Johansen and Carrington (2001).
Constructs Used for Plant Transformation The following vectors and constructs were used in this study (Fig. 2):
| • | Construct A (binary vector pFGC5941 without any insert). This construct was used as negative control. |
| • | Construct B (binary vector pFGC5941 with the sense orientation of the silencing trigger sequence). |
| • | Construct C (binary vector pFGC5941 with the antisense orientation of the silencing trigger sequence). |
| • | Construct D (binary vector pFGC5941 with sense/antisense orientation of the silencing trigger sequence separated by the ChaS intron to create the dsRNA inside the host cells). |
Inoculation with PVX-Eg2 Potato and N. benthamiana plants previously transformed with different constructs were infected with Egyptian isolate of PVX-Eg2 using mechanical transmission as follows: sap from infected plants was extracted from young potato leaves with neutral or weakly basic 0.05 M phosphate buffer
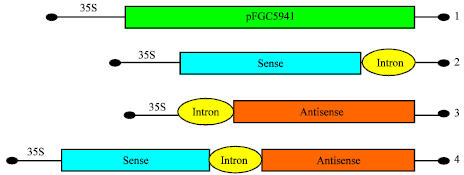 | |
| Fig. 2: | Constructs used to transform potato and N. tabaccum plants. 1: a plant binary vector, 2: 35S with sense orientation of silencing construct, 3: 35S with antisense orientation of silencing construct and 4: 35S with sense/antisense orientations of silencing construct |
(pH 7.2). The sap was clarified through two layers of cheesecloth and the plants were dusted with carborundum. The crude sap was used as an inoculum into 2-3 leaf-stage healthy seedlings. All inoculated plants were kept under controlled conditions in the growth chamber with 16 h of light and 8 h of darkness and at 25°C.
Evaluation of the Resistance Developed Against PVX-Eg2 Screening Using Viral Symptoms
Resistance developed against PVX-Eg2, in both potato and N. benthamiana plants those were transformed with different constructs and inoculated with the Egyptian isolate of PVX-Eg2, was evaluated by daily observation for the development of virus symptoms.
Screening with RT-PCR Total RNA was used as template for each RT-PCR amplification reactions using QIAGENTM One-Step RT-PCR Kit. PVX-Eg2cp gene was detected by PCR using primers for CP gene (Shalaby et al., 2002). Forward primer (PVXCPvEcoRI) sequence was: 5`-GATAGAATTCAGAT GACTACACCAGCCAACACC-3', with EcoRI restriction enzyme site (underlined) at the 5` end and the complementary primer (PVXCPcNcoI) sequence was: 5`-TACGCGTCGGTTCCATGGACGT AGTTATGGTGG-3`, with NcoI restriction enzyme site (underlined) at the 5` end. PCR-amplified fragments were separated by gel electrophoresis using 1% agarose gels in 0.5 X TBE buffer, stained with ethidium bromide and visualized with UV illumination using Gel Documentation System. The expected size of the PCR product was 750 bp.
RESULTS
Amplification of the Selected Fragment RT-PCR amplification of viral RNA was carried out on the total RNA isolated from PVY-Eg2-infected plants using specific primers designed to amplify the selected fragment of the coat protein gene of PVX-Eg2cp. Electrophoresis analysis of RT-PCR product of the selected fragment showed a single amplified fragment of 259 bp while no fragment was amplified from potato healthy plant (Fig. 3).
Enzyme Digestion, Cleaning and Cloning of the Selected Fragment into PinPointâ„¢ Xa-1 T-Vector Several white colonies resistant to kanamycin were selected to test for recombinant plasmids containing the selected fragments. Enzyme digestion for PinPointâ„¢ Xa-1 T-Vector carrying the DNA
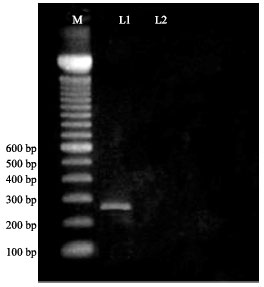 |
| Fig. 3: | Agarose gel electrophoresis analysis of RT-PCR amplified products M: 100 bp DNA ladder (Gibco BRL); L1: total RNA extract from potato leaf sample infected with PVX-Eg2, L2: healthy potato sample |
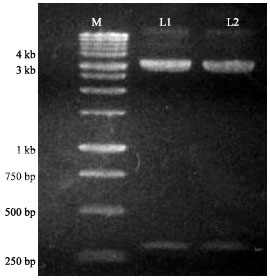 |
| Fig. 4: | Enzyme digestion of the recombinant plasmids from the cloning of the selected fragment. M: 1 Kb DNA ladder (PROMEGA); L1 and L2: DNA minipreps digested with BamHI |
minipreps of the selected colonies of the three constructs i.e., (psPVXcpGS) for sense and (pasPVXcpGS) for anti-sense and (ps/asPVXcpGS) for sense/anti-sense was done using restriction enzymes SwaI and AscI for having sense orientation and BamHI and XbaI for having anti-sense orientation as well as AscI and XbaI for having sense/anti-sense orientation. Cleaning was done, for the cut band using sharp scalpel, using QIAEX II Agarose Gel Extraction Protocol. It is shown in Fig. 4 (lower bands with size 259 bp).
Nucleotide Sequence Analysis Nucleotide sequencing of the RT-PCR product in the recombinant plasmid for the selected fragment of PVX-Eg2cp was completed to confirm PCR-produced fragment is belonged to Potexvirus
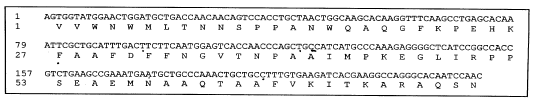 |
| Fig. 5: | Nucleotide sequence of the selected fragment of the coat protein gene of the Egyptian isolate of PVX (PVX-Eg2) and the predicted amino acids below |
 |
| Fig. 6: | PVX symptoms appeared on tobacco (a) and potato (c) plants treated with sense or antisense constructs after challenging with PVX-Eg2 compared with no symptoms on tobacco and potato plants those were treated with sense/antisense construct (b and d, respectively) |
group sequence available at GeneBank. Nucleotide sequence is shown in Fig. 5. Sequence of PVX-Eg2cp fragment was submitted in the GenBank under Accession No. DQ412730.
Sub-Cloning of the Selected Fragment into Binary Vector The selected fragment was sub-cloned into the binary vector pFGC5941 in both orientations (sense and antisense) once by the SwaI and AscI to direct the sense orientation and another time with BamHI and XbaI to direct the antisense orientation leaving the Chalcon Synthase (ChaS) Intron. The Intron (ChasS) was left in the middle of the third construct (sense/antisense).
Evaluation of the Resistance Developed Against PVX-Eg2 Screening Using Bioassay Typical PVX symptoms were appeared in all positive control (non-infiltrated) plants three weeks after PVX-Eg2 inoculation. Symptoms were also recorded on all of infiltrated plants with the following: a) pFGC5491 vector without any insert, b) sense construct, c) antisense construct. Agro-infiltrated plants with sense/antisense construct did not produce viral symptoms (Fig .6)
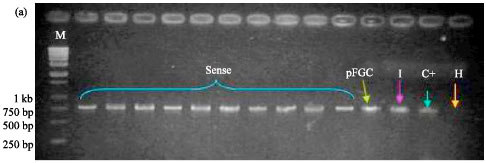 |
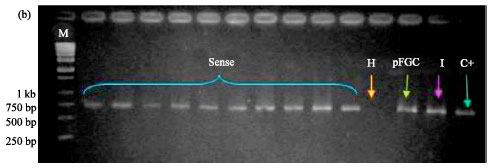 |
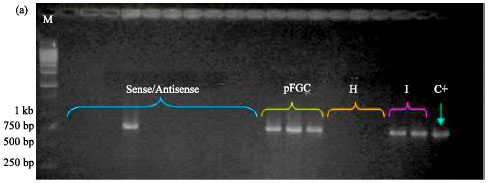
| Fig. 7: | PCR amplification for the potato (a) and tobacco (b) plants agro-infiltrated with sense construct. M: 1 Kb DNA ladder (PROMEGA). Sense: 10 extracts of potato plants agro-infiltrated with sense construct, pFGC: pFGC5491 vector without insert, I: PVX-Eg2-infected plant, C+: positive control, H: healthy plant (negative control) |
Screening Using RT-PCR PCR was carried out, as more sensitive test, to confirm previous bioassay results. Figure 7a and 8a show positive amplification of the expected size (750 bp) fragment that was obtained with the following: a) non-infiltrated potato plants, b) infiltrated potato plants with (b1) pFGC5491 vector without any insert, (b2) sense construct indicating the presence of viral genome in challenged plants. Figure 7b and 8b show same positive amplification of the expressed size (750 bp) fragment that was obtained with the following: a) non-infiltrated tobacco plants, b) infiltered tobacco plants with (b1) pFGC5491 vector without any insert, (b2) antisense construct; indicating the presence of viral genome.
Infiltrated tobacco and potato plants with sense/antisense construct gave negative amplification indicating the absence of the viral genome in 9 out of 10 potato plants and in 7 out of 10 tobacco plants (Fig. 9).
DISCUSSION
Twenty one to twenty three nt RNA known as siRNA is a key component of gene silencing (Voinnet, 2002; Llave et al., 2002). This study has used 21 nt as siRNA. One conserved, un-translated sequence from the coat protein gene of PVX-Eg2 (PVX-Eg2cp) was selected to develop resistance against PVX-Eg2. Successful application of gene silencing technique to develop resistance against potato virus X (PVX-Eg2) is shown in Fig 6 and 9. Previous studies on virus resistance by post-transcriptional gene silencing depended highly on parts of one gene to switch off or eliminate the viral replication in the host plants (Beachy, 1997). PTGS with the subsequent production of virus-derived siRNAs in the case of potato virus X-infected plants was published by Hamilton and Baulcombe (1999). This induced defense is characterized by sequence specific resistance against virus infection
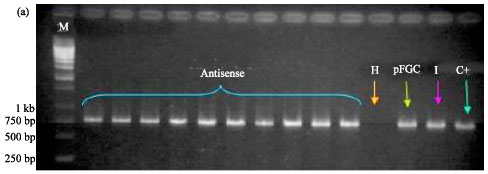 |
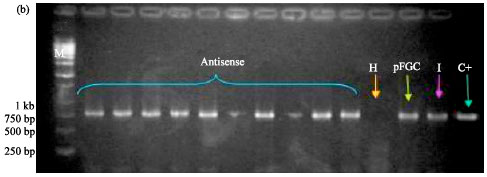 |
| Fig. 8: | PCR amplification for the potato (a) and tobacco (b) plants agro-infiltrated with antisense construct. M: 1 Kb DNA ladder (PROMEGA), Antisense: 10 potato plants agro-infiltrated with antisense construct, pFGC: pFGC5491 vector without any insert, I: PVX infected plant, C+: positive control, H: healthy plant (negative control) |
(Ratcliff et al., 1999). PTGS using anti-sense or co-suppression constructs usually results in only a modest proportion of silenced individuals (Wesley et al., 2001). This study showed the potential of sense/antisense construct encoding self complementary hairpin RNA (hpRNA) to efficiently silence PVX-Eg2 genes. Inclusion of an intron in these constructs (Fig. 3) has a consistently enhanced effect. This technique is supported by Wesley et al. (2001) who explained that Intron-containing constructs (ihpRNA) generally gives 90-100% of independent transgenic plants showing silencing. Agrobacterium tumefaciens-mediated transient expression system, which was used in this study, is a versatile tool to rapidly introduce genes into plant tissue. This system has also been used effectively by Johansen and Carrington (2001) as a mean to deliver RNA silencing inducers and suppressors into transgenic plants that express a silencing reporter gene. Moreover, results of this study has been approved by Feng et al. (2003) who explained that modified PVX-CP gene and wild-type CP gene, those were inserted into binary vector under the control of a promoter and these constructs were transferred into tobacco genomes via Agrobacterium mediated method, have been analyzed using Northern blot of RNA isolated from these plants showed that the silencing ratio of the modified gene in transgenic tobaccos was higher than that of the wild gene. Syringe-Spotting Technique that was used in this study has been efficient to deliver the different constructs might be due to the fact that small leaf tissues of newly developed seedlings posses a large amount of meristematic cells, which provide the best host-cells for both the bacterium to attached and the DNA to express this was supposed by Eckardt (2002). Moreover, amplicon-mediated gene silencing provides an important new strategy for the consistent activation of gene silencing in transgenic plants (Angell and Baulcombe, 1997). Transfection of siRNA is commonly transported through the phloem (Ruiz-Medrano et al., 2001; Lucas et al., 2001) and that regulation of RNA trafficking plays an important role in resistance development and to its role in
 |
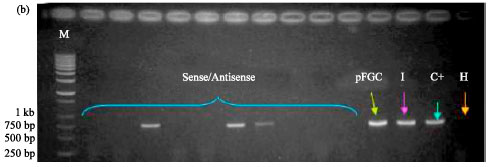 |
| Fig. 9: | PCR amplification for the potato (a) and tobacco (b) plants agro-infiltrated with sense/antisense construct. M: 1 Kb DNA ladder (PROMEGA), Sense/Antisense: 10 potato plants agro-infiltrated with sense/antisense construct, pFGC: pFGC5491 vector without any insert, I: PVX infected plant, C+: positive control, H: healthy plant (negative control) |
PTGS (Vance and Vaucheret, 2001; Haywood et al., 2002). Potato and N. benthamiana plants transformed with the empty vector and later inoculated with PVX developed diseases symptoms 2-3 weeks after inoculation (Fig. 6-8) indicating that the binary vector alone was unable to develop resistance against PVX. Similarly, all potato and N. benthamiana plants transformed with sense construct (Fig. 7) were susceptible to PVX-Eg2 infection indicating that mRNA resulted from the transcription of the sense silencing did not code for any protein and did not anneal to the viral mRNA during the viral infection cycle. Also, both potato and tobacco plants those were transformed with antisense construct, had mild symptoms with PVX-Eg2 infection suggesting that the rate of disease development in these plants was lower (Fig. 8). High level of resistance was induced in potato and N. benthamiana plants against PVX-Eg2 (Fig. 6, 9) when they were transformed with sense/antisense construct. These results demonstrate that the sense/antisense orientation could produce dsRNA folding by the complementarities between the sense and the antisense orientations of the silencing genes after the splicing of the ChaS intron post-transcriptionally.
siRNA technology could be used as significant virus-control measure. As indicated high rate (nine potato plants out of 10) were resistant to PVX-Eg2 when they were transformed with sense/antisense construct. Also, seven tobacco plants out of 10 ones were resistant to PVX-Eg2 when they were transformed with the previous construct.
REFERENCES
- Angell, S.M. and D.C. Baulcombe, 1997. Consistent gene silencing in transgenic plants expressing a replicating potato virus X RNA. EMBO. J., 16: 3675-3684.
CrossRef - Bass, B.L., 2000. Double-stranded RNA as a template for gene silencing. Cell, 101: 235-238.
CrossRefDirect Link - Beachy, R.N., 1997. Mechanisms and applications of pathogen-derived resistance in transgenic plants. Curr. Opin. Biotechnol., 8: 215-220.
PubMed - Feng, D., X. Liu, K. Meng, L. Liao, X. Wei, H. Xu and Z. Zhu, 2003. Silencing of potato virus X coat protein gene in transgenic tobaccos by codon replacement that confers resistance to PVX infection. Chin. Sci. Bull., 48: 1592-1598.
CrossRef - Hamilton, A. and D.C. Baulcombe, 1999. A species of small antisense RNA in posttranscriptional gene silencing in plants. Science, 286: 950-952.
PubMedDirect Link - Hammond, S.M., A.A. Caudy and G.J. Hannon, 2001. Post-transcriptional gene silencing by double-stranded RNA. Nat. Rev. Genet., 2: 110-119.
Direct Link - Haywood, V., F. Kragler and W.J. Lucas, 2002. Plasmodesmata: Pathways for protein and ribonucleoprotein signaling. Plant Cell, 14: 303-325.
Direct Link - Johansen, L.K. and J.C. Carrington, 2001. Silencing on the Spot. Induction and suppression of RNA silencing in the Agrobacterium-mediated transient expression system. Plant Physiol., 126: 930-938.
Direct Link - Llave, C., K.D. Kasschau, M.A. Rector and J.C. Carrington, 2002. Endogenous and silencing-associated small RNAs in plants. Plant Cell, 14: 1605-1619.
Direct Link - Lucas, W.J., B.C. Yoo and F. Kragler, 2001. RNA as a long-distance information macro-molecule in plants. Nat. Rev. Mol. Cell Biol., 2: 849-857.
CrossRefDirect Link - Orman, B.E., R.M. Celnik, A.M. Mandel, H.N. Torres and A.N. Mentaberry, 1990. Complete cDNA sequence of a South American isolate of potato virus X. Virus Res., 16: 293-306.
CrossRef - Ratcliff, F.G., S.A. McFarlane and D.C. Baulcombe, 1999. Gene silencing without DNA. RNA-mediated cross protection between viruses. Plant Cell, 11: 1207-1215.
Direct Link - Ruiz-Medrano, R., B. Xoconostle-Cazares and W.J. Lucas, 2001. The phloem as a conduit for inter-organ communication. Curr. Opin. Plant Biol., 4: 202-209.
CrossRefPubMedDirect Link - Shalaby, A.A., M.K. Nakhla, A.M. Soliman, H.M. Mazyad, A. Hadidi and D.P. Maxwell, 2002. Development of a highly sensitive multiplex reverse trans-cription-polymerase chain reaction (m-RT-PCR) method for detection of three potato viruses in a single reaction and nested PCR. Arab J. Biotechnol., 5: 275-286.
Direct Link - Smith, K.M., 1931. On the composite nature of certain potato virus diseases of the mosaic group as revealed by the use of plant indicators and selective method of transmission. Roy. Soc. London Proc. Ser. B., 109: 251-267.
Direct Link - Soliman, A.M., B.N. Barsoum, G.G. Mohamed, A.K. El-Attar and H.M. Mazyad, 2006. Expression of the coat protein gene of the Egyptian isolate of potato virus X in Escherichia coli and production of polyclonal antibodies against it. Arab J. Biotechnol., 9: 115-128.
Direct Link - Vance, V. and H. Vaucheret, 2001. RNA silencing in plants-defense and counterdefense. Science, 292: 2277-2280.
Direct Link - Voinnet, O., 2001. RNA silencing as a plant immune system against viruses. Trends Genet., 17: 449-459.
Direct Link - Voinnet, O., 2002. RNA silencing: Small RNAs as ubiquitous regulators of gene expression. Curr. Opin. Plant Biol., 5: 444-451.
Direct Link - Waterhouse, P.M., M.B. Wang and T. Lough, 2001. Gene silencing as an adaptive defense against viruses. Nature, 411: 834-842.
Direct Link - Wesley, S.V., C.A. Helliwell, N.A. Smith, M. Wang, D.T. Rouse, Q. Liu and P.S. Gooding, 2001. Construct design for efficient, effective and high-throughput gene silencing in plants. Plant J., 27: 581-590.
CrossRef