
Sabry Y.M. Mahmoud
Department of Agriculture Botany (Microbiology), Faculty of Agriculture, Sohag University, Sohag 82786, Egypt
Sabah A. Abo-El Maaty
Department of Botany, Faculty of Science, Banha University, Banha, Egypt
Ali M. El-Borollosy
Virology Lab., Department of Agriculture Microbiology, Faculty of Agriculture, Ain Shams University, Cairo, Egypt
Mamdouh H. Abdel-Ghaffar
Virology Lab., Department of Agriculture Microbiology, Faculty of Agriculture, Ain Shams University, Cairo, Egypt
International Journal of Virology
Year: 2008 | Volume: 4 | Issue: 1 | Page No.: 1-13







ABSTRACT
Onion yellow dwarf potyvirus (OYDV) causes mosaic in garlic plants, together with other Potyviruses, Carlaviruses and Alexiviruses. A garlic isolate of OYDV (OYDV-G) presenting typical yellow stripes was isolated from naturally garlic mixed infected plants and then biologically purified by single local lesion method on leaves of Chenopodium amaranticolor plants. The obtained isolate was mechanically transmitted and caused chlorotic local lesions in C. a maranticolor but necrotic local lesions to C. quinoa and C. Album. The virus had narrow host range, restricted mainly in some plants belonging to family Alliaceae and Chenopodiaceae. The OYDV-G was aphid transmitted in a non-persistent manner through Myzus persicae. Cytoplasmic Cylindrical Inclusions (CI) of pinwheels and laminated aggregates were observed in ultrathin sections of infected garlic leaf cells. The virus was purified from OYDV-infected garlic tissues by modified procedure involved clarification with chloroform and carbon tetrachloride then concentrated by two cycles of centrifugation, first on a 20% sucrose cushion and second on a 0-40% cesium chloride. Ultraviolet absorption spectrum of the purified virus showed a typical curve of nucleoprotein with A260/280 of 1.450-1.560. The yield of purified virus was 12-15 mg kg-1 infected garlic leaves. The model length of the purified virus was 725-750 nm. The molecular weight of viral coat protein was 30 kDa when determined by SDS-PAGE. The polyclonal antibodies against OYDV-G were produced and the antiserum titer was determined using repeat-indirect enzyme linked immunosorbant assay (R-ELISA). OYDV-RNA was detected in the purified virus preparation and infected garlic samples by immunocapture/reverse transcription-polymerase chain reaction (IC/RT-PCR) using two oligonucleotide specific primers to amplify the common central region of OYDV coat protein gene with size length of approximately 601 bp. The concentration of the virus in the apical and youngest leaves was relatively low comparing with that in the more mature leaves of garlic plants. In addition, the virus isolate was successfully detected in cloves of garlic.
PDF Abstract XML References Citation
How to cite this article
Sabry Y.M. Mahmoud, Sabah A. Abo-El Maaty, Ali M. El-Borollosy and Mamdouh H. Abdel-Ghaffar, 2008. Identification of Onion yellow dwarf potyvirus as One of the Major Viruses Infecting Garlic in Egypt
. International Journal of Virology, 4: 1-13.
DOI: 10.3923/ijv.2008.1.13
URL: https://scialert.net/abstract/?doi=ijv.2008.1.13
DOI: 10.3923/ijv.2008.1.13
URL: https://scialert.net/abstract/?doi=ijv.2008.1.13
INTRODUCTION
Commercially grown garlic (Allium sativum L.) is a species that can only be vegetatively propagated by bulbs and this condition favors a process of virus accumulation overtime in plant material. More than eight viruses have been detected in garlic, including members of the Potyvirus, Carlavirus and Allexivirus genera, which often found forming a viral complex that causes a disease known as garlic mosaic (Van Dijk, 1993a, b; Dovas et al., 2001a; Dovas and Vovlas, 2003). Members of the genus Potyvirus cause significant losses in garlic (Lot et al., 1998). Among these, Onion yellow dwarf virus (OYDV) and Leek yellow stripe virus (LYSV) were the first two viruses described and characterized in garlic plants (Bos, 1976, 1981). Several estimates have proposed that garlic mosaic can generate up to 88% losses in bulb weight (Walkey, 1989; Canavelli et al., 1998; Lot et al., 1998). Onion yellow dwarf virus was found at a higher frequency than LYSV and being the main cause of losses. The OYDV has been distributed worldwide (Brunt et al., 1996; Barg et al., 1994) including Egypt (El-Kewey et al., 2004) in garlic plants. The effect of other members in the mosaic complex is largely unknown. The OYDV is a member of family Potyviridae, genus Potyvirus composed of the aphid-borne viruses with Potato virus Y (PVY) as the type species that characterized by a monopartite, positive-sense, single stranded RNA genome encapsidated in flexuous filaments particles (Brunt et al., 1996).
In Egypt, relatively little attention has been paid to characterize this virus. Therefore, this study aimed to identify an Egyptian of OYDV garlic isolate (OYDV-G) based on the biological and molecular properties together with production of antiserum for virus detection.
MATERIALS AND METHODS
Source of Virus Isolate
The virus isolate was isolated from naturally infected garlic (Allium sativum cv. Chinese) plants cultivated in the open fields of the Faculty of Agriculture, Sohag University, Sohag, Egypt. These plants were exhibited virus-like symptoms, i.e., severe yellow stripe, stunting and malformation.
Virus Isolation and Host Range Test
Young leaves of naturally infected garlic plants were triturated in 0.05 M borate buffer, pH 8.1 containing activated charcoal (1:100 w/v) and sodium diethyldithiocarbamate (1:1000 w/v). Extracted sap was mechanically inoculation on leaves of Chenopodium amaranticolor. After local lesions appearance, the virus isolate was biologically purified by three cycles of single local lesion isolation and then mechanically transmitted to virus-free garlic seedlings cvs. Balady and Chinese, which kindly obtained from tissue culture Lab, ARC, Giza, Egypt. The inoculated garlic plants were maintained under aphid-free conditions in the greenhouse and all subsequent work was done with material from this source. Seventeen plant species belonging to five families, i.e., Alliaceae, Chiniopodiaceae, Solanaceae, Cucurbitaceae and Leguminaceae were used for studying the host range of the virus isolate.
Aphid Transmission
In non-persistent manner, Myzus persicae was tested for transmission OYDV from garlic to garlic according to Lunello et al. (2002). Five aphids per plant and 20 plants were used in this study. Thirty days later, all plants were checked for the presence of the virus by back inoculation on C. amaranticolor.
Types of Inclusions
To identify types of inclusions induced by OYDV, systematically OYDV-infected leaves of garlic cv. balady plant 30 days post-mechanical inoculation were subjected to ultrathin sections according to Lunello et al. (2002). Sections were stained with 2% uranyl acetate then lead citrate and examined in Jeol-1010 transmission electron microscopy at EM unit, Sohag University, Sohag, Egypt.
Virus Purification
The purification procedure developed for OYDV was based on the method of Helguera et al. (1997) with some modifications. Infected garlic (Allium sativum cv. Chinese) leaves were harvested 35 days post virus mechanical inoculation. The infected leaf tissues were homogenized in 2 volumes (w/v) of extraction buffer (0.05 M borate buffer pH 8.1 containing 0.2% thioglicolic acid, 50% carbon tetrachloride and an equivalent volume of chloroform). The resulting slurry was centrifuged at 9000 rpm for 10 min and the supernatant was layered on a 20% sucrose cushion and centrifuged at 23,000 rpm for 2 h at 4°C. Precipitates were resuspended in 0.05 M borate buffer pH 8.1 containing 0.5 M urea and layered on a 0-40% CsCl gradient prepared in the same buffer plus 20% sucrose. The gradient was centrifuged at 28,000 rpm for 5 h at 7°C. A visible band was collected and diluted in resuspension buffer and concentrated by centrifuging for 2 h vat 50,000 rpm at 4°C. The virus concentration was estimated spectrophotometrically by using extinction coefficient of 2.4 for 1 mg mL-1 solution (Stace-Smith and Tremaine, 1970). Virus particles were also observed in purified preparations negatively stained with 2% uranyl acetate according to Kitajima and Nome (1999) and the modal length was calculated after print micrographs.
Estimation of the Molecular Weight of the Viral Coat Protein
The molecular weight of the OYDV-coat protein was estimated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) on a 3.5% stacking gel and a 12% resolving gel. A mixture of an equal amount of virus preparation and dissociation buffer (0.12 M Tris-HCl, pH 6.7, 3% SDS, 20% glycerol and 10% 2-mercaptoethanol) was boiled for 5 min. After electrophoresis for 2 h at 120 V at room temperature, protein bands were visualized by staining with Coomassie Brilliant Blue. The molecular weight of the OYDV-coat protein was estimated by comparison with those of protein molecular weight standards (14.3-200 kDa).
Production of OYDV-Polyclonal Antibodies
The polyclonal antibodies to OYDV were prepared by injecting two adults New Zealand white female rabbits with purified viral preparation. Before starting the injection schedule, the rabbits were bled to obtain the normal serum. Initially, 500 μg mL-1 of purified virus was emulsified with 1 mL of Freund`s complete adjuvant (Sigma Chemical Co., USA) and then injected subcutaneously, followed after 2 weeks by four intramuscular booster injections at weekly intervals using incomplete adjuvant. The rabbits were bled three times, started one, two and three weeks after the last injection. The antisera were separated and collected, then overnight-cross absorbed at 4°C with 5% (w/v) extracted sap of healthy Allium sativum cv. Chinese plants prepared in TBS-Tween buffer (10 mM Tris-HCl buffer, pH 8.0, 150 mM NaCl, 0.05% Tween 20). The mixture was then frozen for 3-4 h, thawed and centrifuged at low speed. The supernatants; cross absorbed antiserum from the three bleeding were tittered by repeat-indirect enzyme linked immunosorbant assay (R-ELISA) based on the method of Baunoch et al. (1992).
OYDV-immunoglobulin G (OYDV-IgG) was isolated and purified from the crude antiserum; which gave a high titer by indirect R-ELISA, according to the method of Perosa et al. (1990) with some modifications. The antiserum was dialyzed against sodium acetate buffer, pH 4.8 and then the caprylic acid was added followed by centrifugation at a low speed. The supernatant containing the IgG was collected and dialyzed against potassium phosphate buffer, pH 7.2, then precipitate by saturated ammonium sulfate solution and low speed centrifugation. The IgG-pellets were resuspended in distilled water followed by dialyzing against phosphate buffer, pH 7.2. The concentration of IgG was adjusted to 1 mg mL-1 when the Optical Density (OD) at wavelength 280 nm was 1.46. The OYDV-IgG was stored at -20°C until use.
Immunocapture/Reverse Transcription-Polymerase Chain Reaction (IC/RT-PCR)
IC/RT-PCR was performed to confirm the specificity and high sensitivity detection between the OYDV particles and its IgG that obtained from this study. IC/RT-PCR was done in a one-tube single step method using 200 μL MicroAmp optical tubes, previously coated for 2 h at 37°C with 50 μL of an OYDV-IgG, diluted 1/500 in 50 mM carbonate buffer (pH 9.6). OYDV-infected and healthy garlic leaves were ground 1:10 (w/v) in extraction buffer according to Clark and Adams (1977) and subjected to IC at 4°C overnight as described by Nolasco et al. (1993). The RT-PCR was carried out based on the method modified from Sellner et al. (1992). The forward and reverse oligonucleotide specific primers were designed after a search of sequences available in the GenBank database (www. rcbi.nlm.nih.gov) and applying the Primer Express software (PE Biosystem) for amplify the common central region of OYDV cp gene (Takaichi et al., 1998). The nucleotide sequences of those primers were as follows; forward 5` CGA AGC AAA TTG CCA AGC AG 3` and reverse 5` CGA TTA GCT GCC CCT CTA AC 3`. RT-PCR conditions used were 30 min at 48°C of retrotranscription, 10 min at 95°C as an initial polymerase activation step, followed by 27 amplification cycles of 1 min denatured at 95°C, 1 min annealed at 57°C, 1 min extended at 72°C and one final cycle of 5 min at 72°C. The resulting reaction mixture was analyzed by 1% agarose gel electrophoresis.
Virus Distribution Within Garlic Plant
In this study, variations in viral concentration within a plant (green leaves and clove storage leaves) was determined using indirect DAS-ELISA based on the method of Clark and Adams (1977). Three of OYDV-infected garlic plants cultivars Balady and Chinese were used for such purpose. All leaves per plant were sampled one month before harvesting and numbered according to position, from top leaf (No. 1) to older one (No. 6). Also, the presence of OYDV was tested in dormant cloves of garlic at harvest.
RESULTS
Mechanical Isolation of OYDV and Symptoms on Test Plants
The local lesions showed in the inoculated C. amaranticolor and C. quinoa provided an evidenced that more than a single virus had existed. Necrotic and chlorotic local lesions (NLL and CLL) were developed in C. amaranticolor leaves inoculated with symptomatic garlic extract. Developed two types of local lesions were different in size and shape (Fig. 1A). The two virus isolates were isolated and separated via single local lesion isolation on C. amaranticolor. On the other hand, the infected garlic sap was exposed to 60°C for 10 min and then inoculated to C. amaranticolor. These process results in one type of necrotic local lesions as shown in C. amaranticolor and C. quinoa Fig. 1B, C, respectively. Single local lesions were cut from C. amaranticolor and inoculated in virus-free garlic plants cv. Balady; the symptoms occurred as numerous striking, short yellow streaked spots in the basal of leaves and then became completely chlorotic, partially flattened, crinkled and wilted (Fig. 2A, E). On the other hand, sever mosaic was induced on garlic plants cv. Chinese Fig. 2F, G, yellow spot, streaking, zigzag leaves were occurred on inoculated onion plants Fig. 2H and yellow spots were induced on Narcissus sp. Fig. 2I.
 | |
| Fig. 1: | The local lesions i nduced by inoculation of infected garlic sap, (A) from untreated sap on C. amaranticolor. After sap exposure to 6 0°C for 10 min on C. amaranticolor (B) and C. quinoa (C) |
 | |
| Fig. 2: | Symptoms induced on test plants after mechanical inoculation by OYDV-garlic isolate (B-E) Development of symptoms on garlic plants cv. Balady compared with control in (A F , G) Garlic plants cv. Chinese; (H) Onion leaves and (I) Narcissus leaves |
The obtained results indicated that the OYDV isolate can be easily transmitted by sap inoculations to 60% garlic plants. The plants used for host range test belonging to five different families are shown in Table 1. Results indicated that the tested virus isolate has a narrow host range restricted in some species and cultivars of garlic and onion plants as well as the diagnostic plants, i.e., Chenopodium sp., which were infected and positively reacted with the virus isolate (Fig. 1, 2), but the virus isolate could not be mechanically transmitted to leek plants. All other inoculated plant species belonging to families Solanaceae, Cucurbitaceae and Leguminaceae showed negative results. These results were confirmed by back inoculation to C. quinoa.
Aphid Transmission
Results showed that the OYDV was transmitted in a non-persistent manner by M. persicae from infected garlic to healthy garlic plants. When checked for the presence of OYDV 30 days post-aphid transmission by characteristic symptoms and back inoculation on C. amaranticolor, only 35% were found to be infected.
Ultrathin Sections
The electron micrographs of ultrathin sections of A. sativum cv. Chinese leaves, exhibited the characteristic symptoms of OYDV, showed cylindrical Cytoplasmic Inclusions (CI) as pinwheels and laminated aggregates (Fig. 3A, B) in infected mesophyll cells.
| Table 1: | Reactions of test plants mechanically inoculated with OYDV-Garlic isolate |
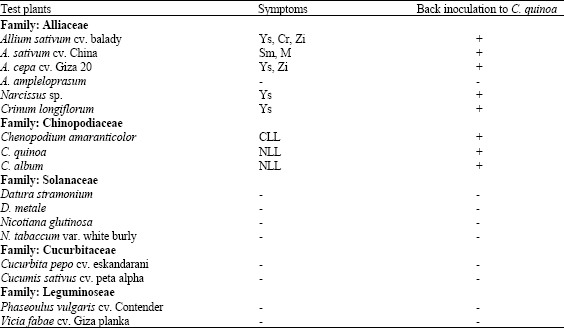 | |
| -: No symptoms, CLL: Chlorotic Local Lesion, Cr: Crinkle, M: Mosaic NLL: Necrotic Local Lesion, Sm: Severe mosaic, Ys: Yellow streak, Zi: Zigzag leaf | |
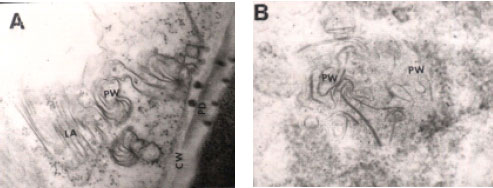 | |
| Fig. 3: | Electron micrographs for the mesophyll cells of A. sativum cv. Chinese leaf plant, 30 days post mechanical inoculation. Note, cylindrical cytoplasmic inclusions; pinwheels (PW) and laminated aggregates (LA), Cell Wall (CW), plasomodesmata (PD). (A, x 30,000; B, x 25,000) |
Virus Purification
The Ultraviolet-absorption spectrum of purified OYDV (Fig. 4), showed a typical curve for nucleoprotein with A260 peak and A245 minimum. The relation of Amax/min was 1.133-1.158 and A260/280 was -1.450-1.560. The purification method yielded 12-15 mg virus purified/1kg infected leaf tissues based on the extinction coefficient of 2.4 for 1 mg mL-1 solution. Electron micrographs of purified preparation showed numerous flexuous filamentous particles (Fig. 5) with a length of 725-750 nm and 15 nm in width, when negatively stained with 2% aqueous uranyl acetate and examined in transmission electron microscope.
Coat Protein Analysis
The coat protein of virus particles was detected in 12% SDS-PAGE and migrated as a single band with a molecular weight of ~ 30 kDa (Fig. 6).
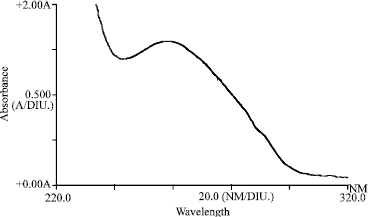 | |
| Fig. 4: | Ultraviolet-absorption spectrum of OYDV purified preparations |
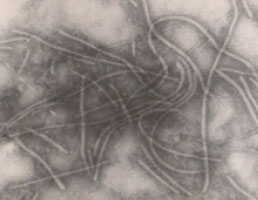 | |
| Fig. 5: | Electron micrograph of purified OYDV, stained with 2% uranyl acetate, (x 60,000) |
 | |
| Fig. 6: | Electrophoresis of OYDV-coat protein in 12% SDS-PAGE. Lane M: marker proteins, Lanes 1 and 2: OYDV- coat protein |
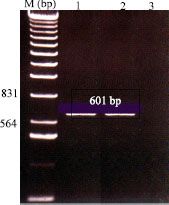 | |
| Fig. 7: | Agarose gel electrophoresis analysis of IC/RT-PCR product of central region of OYDV cp gene, stained with ethidium bromide. Lane 1, positive reaction from OYDV-infected garlic plants. Lane 2, positive reaction from purified OYDV preparation. Lane 3, healthy control (reaction from healthy garlic plants). It should be noted that the obtained amplicons have the expected size (ca. 601 bp) |
| Table 2: | Determination of OYDV antiserum titer by R-ELISA |
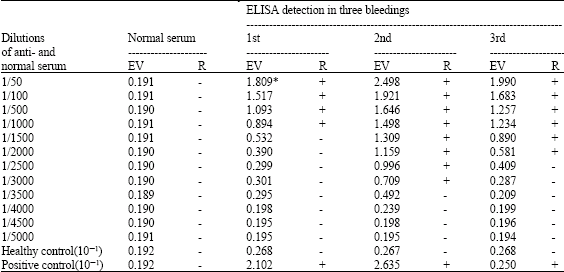 | |
| *: ELISA Value (EV) at 405 nm was an average of 4 replicates, Healthy control: Healthy garlic tissues, Positive control: OYDV-infected garlic tissues Note: The ELISA value that over two folds of healthy control was considered as a positive (+) results | |
The polyclonal antiserum against OYDV isolate was produced and its titer was determined by indirect R-ELISA. Totally three bleedings were obtained from the rabbits immunized with OYDV. Three bleedings varied in their titers. Positive ELISA values were obtained up to dilutions of 1/1000, 1/3000 and 1/2000 for the 1st, 2nd and 3rd bleeding respectively (Table 2). According to titer of different bleedings of OYDV polyclonal antiserum, the 2nd bleeding was used for isolation of IgG. The concentration of IgG was adjusted to 1 mg mL-1 when OD at 280 nm was 1.46.
IC/RT-PCR
IC/RT-PCR was done in a one-tube single step method to confirm the specificity between the purified virus and its IgG. Results indicated that the OYDV particles in both infected garlic tissue and purified virus preparation were immunocaptured by specific OYDV-IgG, thus, the genomic viral RNA could be released and used as template. When primers that amplify the central region of OYDV cp gene were used, a single band with the expected size length (ca. 601 bp) was amplified (Fig. 7) (Lane 1 and 2). A negative result of IC/RT-PCR was obtained with a healthy garlic tissue (Fig. 7; Lane 3).
| Table 3: | ELISA readings for OYDV concentrations in different leaf positions and cloves in garlic plants |
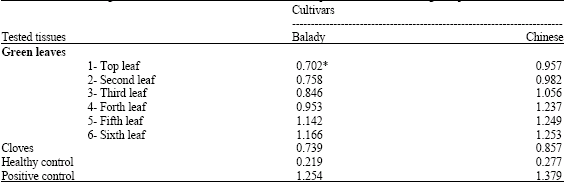 | |
| *: ELISA Value (EV) at 405 nm was an average of 4 replicates, Healthy control: Healthy garlic tissues, Positive control: OYDV-infected garlic tissues, Note: The ELISA value that over two folds of healthy control was considered as a positive (+) results | |
OYDV Distribution Within Garlic Plant
Data Table 3, showed comparison in OYDV concentrations among leaves of garlic plant. OYDV level was low in the apical leaf and then increased up to about the sixth leaf below the apex. Virus concentration was higher in older leaves compared with apical ones. On the other hand, the OYDV was successfully detected in cloves of garlic.
DISCUSSION
Garlic is an economically important crop for several Egyptian agricultural regions. Egyptian growers traditionally produce their own garlic propagative material. This fact accounts for the observed heavy viral infection and implies a potentially high reduction in yield and quality of this crop. To face this problem, a strategy is currently under development in Egypt, for the production of virus-free garlic propagative material. Because limited data are currently available, this study aimed to the identification of some A. sativum infecting viruses. The results showed that garlic in Egypt is heavily infected with viruses. OYDV, probably the most important virus of garlic, which found to be the most abundant and widespread virus (Dovas et al., 2001b). Results of detailed host range and serological studies by Van Dijk (1993a, b) on isolates from a large number of Alliums sp. suggested that, in addition to OYDV, LYSV seems to be widely spread in garlic growing in many countries around the world causing significant degeneration of the crop after only a few multiplications under field conditions (Fajardo et al., 2001; Takaichi et al., 2001).
In present study, OYDV-G was isolated from garlic via serial local lesion transfer on C. amaranticolor and then back inoculated on virus-free garlic plants. The results indicated presence one virus isolate; the evidence for this is biological reactions. Also, the absence of other viruses was confirmed by differential hosts which produced one type of local lesions compared with original samples from nature infections (mixed infections). On the other hand, OYDV-G isolate and OYDV-onion isolate (OYDV-O) are much like in biologically properties. The OYDV-G isolate was mechanically transmitted and caused chlorotic and necrotic local lesions in C. amaranticolor and C. quinoa, respectively and can be transmitted with difficulty to onion. This indicates that OYDV-G is a newly reported isolate of OYDV, since it was mentioned that OYDV-O was easily transmitted to onion (Bos, 1976; El-Kewey and Sidaros, 1996).
Aphid transmission of OYDV-G from garlic to garlic was confirmed by M. persicae that could transmit the virus isolate in non-persistant manner. M. persicae was confirmed as a vector for OYDV (Bos et al., 1978). M. persicae are polyphagous and widely distributed in Egypt. It forms large colonies in numerous horticultural and field crops beside garlic fields. This becomes important since OYDV-G can easily transmit by aphids from garlic to garlic. This should be considered when deciding on control measures.
The cylindrical inclusions of pinwheels and laminated aggregates were observed in OYDV-infected garlic mesophyll cells. This result characterized OYDV as a member of potyvirus subdivision II according to Edwardson´s classification (1974). This result agrees with earlier reports by Verhoyen and Horvat (1973) Paludan (1980) and Lunello et al. (2002).
In the present study, the purification procedure described here was efficient method for the extraction and purification of OYDV-G free of cellular plant components. Extraction and clarification with borate buffer pH 8.1 containing thioglicolic acid, carbon tetrachloride and an equivalent volume of chloroform, then precipitation of the virus particles by two cycles of ultracentrifugation, first on a 20% sucrose cushion and second on a 0-40% cesium chloride were found to be satisfactory without much loss of virus particles. The purified preparation of OYDV-G had ultraviolet absorption spectrum characteristic of nucleoprotein with A260/280 of 1.450-1.560. The estimated yield of purified virus was 12-15 mg kg-1 infected garlic leaves. These results relatively agreed with previous finding by Helguera et al. (1997). The particle model length was 725-750 nm, similar to the 750 nm reported by Helguera et al. (1997).
The molecular weight of viral coat protein was 30 kDa when determined by SDS-PAGE. This data was consistent with that found by Brunt et al. (1996).
In the present study, the polyclonal antibody was successfully raised to the OYDV-G and three bleedings were obtained and showed that the 2nd bleeding was the highest one in its titer using R-ELISA. The OYDV-IgG was purified from the 2nd bleeding using the caprylic acid procedure and use it in ELISA and IC/RT-PCR detection.
In contrast to other virus genera, serology is not a very good parameter for virus differentiation among viruses of the genus Potyvirus, because serological cross reactions often cause misinterpretation of results (Conci et al., 1999) and it is not suitable for potyvirus taxonomy (Shukla and Ward, 1988). These observations supported the application of molecular techniques for characterization of the garlic viruses as demonstrated by others (Nagakubo et al., 1994; Kobayashi et al., 1996; Lot et al., 1998; Tsuneyoshi et al., 1998; Van der Vlugt et al., 1999). In the present study, the positive IC/RT-PCR reaction indicated the specificity between the OYDV-G particle and its IgG, also it confirmed the specificity primers that used to amplify the common central region of OYDV coat protein gene with expected size length of approximately 601 bp. Several papers have reported the use of RT-PCR as highly sensitive technique for detection of viruses in Allium (Dovas et al., 2001a; Shiboleth et al., 2001). RT-PCR using specific primers can be helpful in the detection of new garlic virus`s not yet characterized (Sumi et al., 1993). PCR fragments from OYDV-G and LYSV can be labeled and used as probes. No cross-reaction observes between coat protein sequences of these two potyviruses. It showing that the virus isolates belonged to distinct Potyvirus species, the specific probes can be used for virus differentiation, providing an accurate method for detection of mixed infections of garlic viruses.
The distinct detection methods can be successfully used to monitor a high quality program of virus-free garlic production associated with an efficient program of virus eradication from garlic by meristem-tip culture and thermotherapy (Torres et al., 2000). Because the concentration of OYDV in the apical, physiologically youngest, leaf is low, testing dormant cloves of garlic for presence of OYDV was conducted (Vunsh et al., 1991). This study suggested that the present system is sensitive enough to detect virus in clove storage leaf than in the leaf initials of the dormant plantlets. This test could be used to screen during the summer season, before the bulb is separated into planting material. This procedure may save testing of all resulting plants, since in a normal field planting it is impossible to tell which plants are derived from the same bulb. Since one bulb yields about eight to 10 cloves suitable for planting, testing in the dormant cloves would reduce the number of tests to be carried out.
REFERENCES
- Barg, E., D.E. Lesemann, H.J. Vetten and S.K. Green, 1995. Identification, partial characterization and distribution of viruses infecting Allium crops in South and Southeast Asia. Acta Hortic., 358: 251-257.
CrossRefDirect Link - Baunoch, D.A., P. Das, M.E. Browning and V. Hari, 1992. R-ELISA: Repeated use of antigen-coated plates for ELISA and its application for testing of antibodies to HIV and other pathogens. Biotechniques, 12: 412-417.
Direct Link - Bos, L., N. Huijberts, H. Huttinga and D.Z. Maat, 1978. Leek yellow stripe virus and its relationships to Onion yellow dwarf virus; Characterization ecology and possible control. Netherlands J. Plant Pathol., 84: 185-204.
CrossRef - Brunt, A.A., K. Crabtree, M.J. Dallwitz, A.J. Gibbs and L. Watson, 1996. Viruses of Plants. Descriptions and Lists from the VIDE Database. 2nd Edn., CAB International, Wallingford, UK., ISBN: 0-85198-794-X, Pages: 1484.
Direct Link - Clark, M.F. and A.N. Adams, 1977. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol., 34: 475-483.
CrossRefPubMedDirect Link - Dovas, C.I., E. Hatziloukas, R. Salomon, E. Barg, Y. Shiboleth and N.I. Katis, 2001. Comparison of methods for virus detection in Allium sp. J. Phytopathol., 149: 731-737.
CrossRefDirect Link - Dovas, C. I., E. Hatziloukas, R. Salomon, E. Barg, Y. Shiboleth and N.I. Katis, 2001. Incidence of viruses infecting Alliums sp. in Greece. Eur. J. Plant Pathol., 107: 677-684.
Direct Link - Dovas, C.I. and C. Vovlas, 2003. Viruses infecting Alliums sp. in Southern Italy. J. Plant Pathol., 85: 135-135.
Direct Link - Fajardo, T.V.M., M. Nishijima, J.A. Buso, A.C. Torres, A.C. Avila and R.O. Resende, 2001. Garlic viral complex: Identification of Potyviruses and Carlavirus in Central Brazil. Fitopatologia Brasileira, 26: 619-626.
Direct Link - Helguera, M., P.A. Lunello, C. Nome and V.C. Conci, 1997. Advances in the purification of filamentous viruses from garlic and in antisera production. Acta Hortic., 433: 623-630.
Direct Link - Kobayashi, K., P. Rabinowich, F. Brsvo-Almonacid, M. Helgura, V. Conci, H. Lot and A. Mentaberry, 1996. Coat protein gene sequences of garlic and onion isolates of the Onion yellow dwarf potyvirus (OYDV). Arch. Virol., 141: 2277-2287.
PubMed - Lot, H., V. Chovelon, S. Souche and B. Delecolle, 1998. Effect of Onion yellow dwarf and leek yellow stripe viruses on symptomatology and yield loss of three French garlic cultivars. Plant Dis., 82: 1381-1385.
Direct Link - Lunello, P., D.A. Ducasse, M. Helguera, C.F. Nome and V.C. Conci, 2002. An Argentinean isolate of Leek yellow stripe virus from leek can be transmitted to garlic. J. Plant Pathol., 84: 11-17.
Direct Link - Nagakubo, T., M. Kubo and K. Oeda, 1994. Nucleotide sequences of the 3 regions of two major viruses from mosaic-diseased garlic: Molecular evidence of mixed infection by Potyvirus and a Carlavirus. Phytopathology, 84: 640-645.
CrossRef - Nolasco, G., C. Deblas, V. Torres and F. Ponz, 1993. A method combining immunocapture and PCR amplification in a microtiter plate for the detection of plant viruses and subviral pathogens. J. Virol. Methods, 45: 201-218.
Direct Link - Paludan, N., 1980. Virus attack on leek: Survey, diagnosis, tolerance of varieties and winterhardiness. Tidsskrift Planteavl, 84: 371-385.
Direct Link - Perosa, F., R. Carbone, S. Ferrone and F. Dammacco, 1990. Purification of human immunoglobulins by sequential precipitation with caprylic acid and ammonium sulphate. J. Immunol. Methods, 128: 9-16.
CrossRef - Sellner, L.N., R.J. Coelen and J.S. Mackenzie, 1992. Reverse transcriptase inhibits Taq polymerase activity. Nucleic Acids Res., 20: 1487-1490.
Direct Link - Shiboleth, Y.M., A. GalOn, M. Koch, H.D. Rabinowitch and R. Salomon, 2001. Molecular characterization of Onion yellow dwarf virus (OYDV) infecting garlic (Allium sativum L.) in Israel: Thermotherapy inhibits virus elimination by meristem tip culture. Ann. Applied Biol., 138: 187-195.
Direct Link - Shukla, D.D. and C.W. Ward, 1988. Amino acid sequence homology of coat proteins as a basis for identification and classification of the Potyvirus group. J. Gen. Virol., 69: 2703-2710.
CrossRef - Stace-Smith, R. and J.M. Tremaine, 1970. Purification and composition of Potato virus Y. Phytopathology, 60: 1785-1789.
Direct Link - Sumi, S., T. Tsuneyoshi and H. Furutani, 1993. Novel rod-shaped viruses isolated from garlic, Allium sativum, possessing a unique genome organization. J. Gen. Virol., 74: 1879-1885.
CrossRefDirect Link - Takaichi, M., M. Yamamoto, T. Nagakubo and K. Oeda, 1998. Four garlic viruses identified by reverse transcription-polymerase chain reaction and their regional distribution in Northern Japan. Plant Dis., 82: 694-698.
Direct Link - Takaichi, M., T. Nagakubo and K. Oeda, 2001. Mixed virus infections of garlic determined by a multivalent polyclonal antiserum and virus effects on disease symptoms. Plant Dis., 85: 71-75.
Direct Link - Torres, A.C., T.V. Fajardo, A.N. Dusi, R. de Oliveira Resende and J.A. Buso, 2000. Shoot tip culture and thermotherapy for recovering virus-free plants of garlic. Horticultura Brasileira, 18: 192-195.
Direct Link - Tsuneyoshi, T., T. Mtsumi, K.T. Natsuaki and S. Sumi, 1998. Nucleotide sequence analysis of virus isolated indicated the presence of three potyvirus species in Allium plants. Arch. Virol., 143: 97-113.
PubMed - Van der Vlugt, V.R.A.A., P. Steffens, C. Cuperus, D.E. Lesemann, L. Bos and H.J. Vetten, 1999. Further evidence that Shallot yellow stripe virus (SYSV) is a distinct Potyvirus and reidentification of welsh Onion yellow stripe virus as a SYSV strain. Phytopathology, 89: 148-155.
CrossRef - Van Dijk, P., 1993. Carlavirus isolates from cultivated Allium species represent three viruses. Netherlands J. Plant Pathol., 99: 233-257.
CrossRefDirect Link - Van Dijk, P., 1993. Survey and characterization of Potyviruses and their strains of Allium species. Netherlands J. Plant Pathol., 99: 1-48.
CrossRefDirect Link - Vunsh, R., A. Rosner and A. Stein, 1991. Detection of bean yellow mosaic virus in Gladioli corms by the polymerase chain reaction. Ann. Applied Biol., 119: 289-294.
CrossRef - Walkey, D.G.A. and D.N. Antill 1989. Agronomic evaluation of virus-free and virus-infected garlic (Allium sativum L.). J. Hortic. Sci., 64: 53-60.
CrossRefDirect Link
Siti Shofiya Nasution Reply
Hi. my name is shofi. I am using primers OYDV that you designed. I need your journal to my research. thank you. I hope you can help me.