
H. M. Mazyad
Department of Plant Virus and Phytoplasma Research, Plant Pathology Research Institue, ARC, P.O. Box 1261 9, Giza, Egypt
E. M. Khalil
Department of Vegetable, Horticultural Research Institue,
ARC, P.O. Box 12619, Giza, Egypt
A. A. Rezk
Department of Plant Virus and Phytoplasma Research, Plant Pathology Research Institue, ARC, P.O. Box 1261 9, Giza, Egypt
M.A. Abdel- Hakem
Department of Plant Virus and Phytoplasma Research, Plant Pathology Research Institue, ARC, P.O. Box 1261 9, Giza, Egypt
A.E. Aboul- Ata
Department of Plant Virus and Phytoplasma Research, Plant Pathology Research Institue, ARC, P.O. Box 1261 9, Giza, Egypt
International Journal of Virology
Year: 2007 | Volume: 3 | Issue: 2 | Page No.: 88-95







ABSTRACT
Five crosses derived from crossing between TYLCV-susceptible female, (Edkawy, Castle Rock, Strain-B, Peto-86 and Marmmande) and TYLCV-resistant male (Favi-9) were performed. Favi-9 is derived from Licopersecon hirsutum. Six population of each cross were planted in complete-random blot distribution (CRBD) with 3 replicates during 2002-2004 growing seasons at Kaha, ARC Research Station. Field and laboratory evaluation was done for each population individuals for finding out TYLCV resistance as the main goal. Evaluation process explained that parent hetorosis values were 5.72 to 14.9% and 112.7 to 128.7% for crosses of Marmmande x Favi-9 and Castle Rock x Favi-9, respectively. The heritability estimate was 55.76 and 88.38% for crosses of Edkawy x Favi-9 and Peto-86 x Favi-9, respectively. Potency Ratio Values showed that crosses of Marmmande x Favi-9 and Castle Rock x Favi-9 ranges between -0.11 and -0.30, respectively. Those values are the least ones obtained. The inbreeding depression has presented in most crosses under investigation. Its estimate was ranged between -4.82 and-27.66% for the crosses of Strain-B x Favi-9 and Castle Rock x Favi-9, respectively. Conclusion pointed out that effective gene for TYLCV resistance could be one to two pairs of genes.
PDF Abstract XML References
How to cite this article
H. M. Mazyad, E. M. Khalil, A. A. Rezk, M.A. Abdel- Hakem and A.E. Aboul- Ata, 2007. Genetic Studies on Tomato Yellow Leaf Curl Begomovirus (TYLCV) Resistance in Egypt: Six-population Analysis. International Journal of Virology, 3: 88-95.
DOI: 10.3923/ijv.2007.88.95
URL: https://scialert.net/abstract/?doi=ijv.2007.88.95
DOI: 10.3923/ijv.2007.88.95
URL: https://scialert.net/abstract/?doi=ijv.2007.88.95
INTRODUCTION
Tomato Yellow Leaf Curl Begomovirus (TYLCV) is a whitefly-transmitted geminivirus that affecting tomato plantations in many tropical and subtropical regions. Widespread infection has been confirmed using DNA and protein sequence comparison which indicate that tomato geminiviruses fall into three main clusters. There are only two distinguishable sub-clusters, within the Mediterranean and Middle East cluster (Czosnek and Laterrot, 1997). Moreover, the adult progeny of the viruliferous B. tabacci insects and their own progeny are able to infect tomato test plants, producing typical disease symptoms. So, TYLCV can be transmitted through the egg for at least two successive generations. Those statements leads to, absence of an available plant host, the whitefly may serve as a reservoir of virus between growing seasons (Ghanim et al., 1997). TYLCV has been first recorded during 1969 (Nour El-din et al., 1969) in Egypt. Since then ICM programs are being manipulated for facing TYLCV problem. Recently, DNA sequence of Egyptian TYLCV isolate has 98% homology with TYLCV Israeli isolate (Nakhla et al., 1993). As previously mentioned TYLCV has virulent and out breakable properties that make our goal of this study to looking for inheritance for TYLCV resistance.
MATERIALS AND METHODS
Plant Materials and Field Tests
Field experiments of this study were conducted during three successive growing seasons (2002-2004) at Kaha vegetable research station, Horticulture Research Institute (HRI), ARC which is located 30 Km North Cairo, in Qualubia governorate. Tomato varieties of TYLCV-susceptible female (Edkawy, Castle Rock, Strain-B, Peto-86 and Marmmande) which is commercially available, were obtained from Deptartment of vegetables, HRI, ARC. However, TYLCV-resistant male (Favi-9) was kindly forwarded by Volcani Center, Bet Degan, Israel as part of regional collaboration through MERC project # M21-037. Favi-9 is derived from Lycopersecon hirsutum. First season (2002), seeds of TYLCV-resistant male and TYLCV-susceptible females were sown in the field for confirmation on purity and stability before crossing process. Crossing between previously mentioned male and different females was curried out in the greenhouse to get F1 seeds of each. Seeds of parents and their F1’s were sown in the field to be evaluated and to get F2 seeds during 2003 growing season. Back Cross 1 (BC1) and Back Cross 2 (BC2) were carried out in the greenhouse and produced seeds as well as F2 seeds were sown in the field during 2004 growing season for evaluation.
Field Evaluation
Seeds of six population (P1, P2, F1, BC1, BC2 and F2) of each of five crosses i.e., 1-EdkawyxFavi-9, 2-Castle RockxFavi-9, 3-Strain-BxFavi-9, 4-Peto-86xFavi-9 and 5-Marmmande x Favi-9 were sown in the greenhouse to insure production of virus-free seedlings. Seedlings of P1, P2 and F1, were transplanted in three rows. Seedlings of BC1 and BC2 were transplanted in four rows. Seedlings of F2 were transplanted in 6 rows during growing season in a complete randomized block design with three replicates. Each row was 10.0 m long and 30.0 cm wide. Pesticides were not applied during the growing season to insure TYLCV-infection spreading out. Filed inspection is done once a week for virus infection using visual examination for viral symptoms (leaf curling, leaf yellowing, dwarfing and compact of vegetative growth). Score scale of Muniyappa et al. (1991) for field inspection was used. Scale degrees used are: 1 = no symptoms, 2 = mild symptoms, 3 = moderate symptoms, 4 = sever symptoms and 5 = very sever symptoms. As indicated by Vidavsky and Czosnek (1998), individuals were selected for TYLCV resistance (no symptoms without detectable viral DNA) and tolerance (no symptoms but with detectable viral DNA). Consequently, no symptoms (0-1 degrees) = highly tolerance, mild symptoms (1-2 degrees) = tolerance, moderate symptoms (2-3 degrees) = moderate susceptible, severe symptoms (3-4 degrees) = susceptible and very severe (4-5 degrees) = very susceptible.
Statistical Analysis of Genetic Components
Heterosis was estimated according to the mid-parent value method that was used by Sinha and Khanna (1975). Broad Sense Heritability (BSH) was measured using the method of Allord (1960). Potency ratio was measured according to Smith (1952) methodology. Estimation for number of genes those are controlling TYLCV-resistance character was made according to the method of Castle and Wright (1921). Mean scale for TYLCV severity was estimated as (dsxnp)/tp = mean degree of severity. ds = degree of severity, np = number of infected plants and tp = total plant examined.
Following are detailed equations used in this study:
Heritability
It was calculated using Broad Sense Heritability (BSH): It was calculated using the following equation (Allord, 1960)
| (1) |
Where:
| VG | = | Genetic variance which was calculated by subtracting the environmental variance from phenotypic variance (VE-VP), |
| VP | = | Phenotypic variance |
| VE | = | environmental variance which was calculated as geometric mean variances of the non-segregating population, i.e., parents and F1 |
The Number of Effective Factors Controlling the Measured Trait
It was calculated using: Weber’s formula that was modified by Castle and Wright (1921).
| (2) |
| (3) |
Where:
| P1 | = | Mean of susceptible parent |
| P2 | = | Mean of resistant parent |
| F1 | = | First generation mean |
| F2 | = | Second generation mean |
| VP1 | = | Phenotypic variance of P1 |
| VP2 | = | Phenotypic variance of P2 |
| VF1 | = | Phenotypic variance of F1 |
| VF2 | = | Phenotypic variance of F2 |
Potency ratio
The relative potency of gene set (P) was used to determine the direction of dominance according to the formula of Smith (1952):
F1-MP |
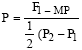 | (4) |
Where:
| F1 | = | First generation mean |
| P1 | = | Mean of the smaller parent |
| P2 | = | Mean of the larger parent |
| MP | = | mid parent value = ½ (P1 + P2) |
The absence of dominance is assumed when the difference between the parents was significant and F1-MP was not significant.
Complete dominance is assumed when potency ratio equal to ±1.0.
Partial dominance is considered when potency ratio is between + 1.0 and -1.0, but not equal zero.
Over dominance (Heterosis) is assumed when potency ratio exceeded ±1.0.
Heterosis
High parent Heterosis = (F1-HP)/HPx100 (Sinha and Khanna, 1975). |
| Where: | F1 | = | Mean of the first hybrid generation |
| H.P. | = | Mean of the high or better parent |
Variance of population mean, which are different from each other, reveals the importance of improving this character by means of recurrent selection. Moreover, coefficient of variance (CV) reveals also, population variability could be exploited in further breeding program by isolating and developing tomato with TYLCV-tolerance character.
RESULTS AND DISCUSSION
TYLCV disease severity and population reaction recorded for six populations (P1, P2, F1, BC1, BC2 and F2) of the five crosses are presented in Table 1 and Fig. 1-5. Parents (P1s and P2) had different degrees of TYLCV tolerance for all crosses (Fig. 1). It is obvious that the P1s are very susceptible whereas P2 is tolerant (Table 1 and Fig. 1). Mean severity of TYLCV for F1’s population was (3.432, 3.594, 3.343, 3.565 and 3.346 for all crosses, respectively) (Table 1) which lied between the mid-parent value (3.049, 3.128, 3.086, 3.271 and 3.165 for all crosses, respectively) and it was nearer to mean of the susceptible parent (Table 1 and Fig. 1). The population reaction curve of F1’s, against TYLCV infection in all crosses, has the same shape with a peak at scale °4 (susceptible). They never show very susceptibility reaction (scale °5) (Fig. 2) meaning TYLCV susceptibility character is moving towards tolerance. So, it could be suggested that there are some sort of partial dominance towered the lower parent of TYLCV-tolerance character for all crosses. Reaction of F2’s population has S-shape curve in all crosses except cross # 4 (Peto-86xFavi-9). Equal percentage of individuals has susceptibility reaction that expressed by scale 3 and °4 (Fig. 3). Moreover, mean severity of F2 was greatly decreased than those of F1’s individuals in all crosses except the cross # 3 (Strain -BxFavi-9) and cross # 4 (Peto-86xFavi-9) (Table 1 and Figs 3). F2’s population-reaction expresses, around 10%, are TYLCV tolerant and around 30% are TYLCV-susceptibility (Fig. 3). Population-reaction expression toward TYLCV tolerance has moved from 0.0-10% (Fig 1 and 3). Backcross one (BC1’s) is presented in Fig. 4. BC1 individuals reacted in S-shape curve in all crosses except Crosses # 4 (Peto-86xFavi-9) (Fig. 4). They were widely distributed with a High tendency towards this parent (susceptible one) (Table 1). Backcross two (BC2’s) is presented in Fig. 5. BC2 populations had the same S-shape reaction curve, in all crosses, against TYLCV infection except cross # 1 (Edkawi x Favi-9) (Fig. 5). They were widely distributed with a high tendency towards the tolerant parent (Favi-9) for all tomato crosses (Table 1).
Statistical analysis of quantitative genetic parameters for different previous five tomato crosses is presented in Table 2. Mid-parent heterosis (relative heterosis) had positive values (12.58, 14.9, 8.33, 8.98 and 5.72) for all five crosses respectively (Table 2). Better parent heterosis (heterobeltiosis) values were (118.4, 128.7, 112.7, 126.9 and 112.9) for the five crosses, respectively (Table 2). Those results mean that, there is a hybrid vigor towards the better parent. It has been confirmed, also, by potency ratio values. Low negative values of potency ratio (-0.2596,-0.2999,-0.16981,-0.1728 and -0.1137) indicating that TYLCV tolerance behaved as partial recessive toward better parent in this trait (Table 2). These findings were in accordance with those obtained by Hassan and Abdel-Ati (1999) as well as Vidavisky and Czosnek (1998) who reported that TYLCV resistance was a recessive trait. Heritability estimate showed that broad sense for TYLCV resistance were (55.79, 83.27, 59.31, 88.38 and 75.64), for all crosses, respectively (Table 2). This is indicating that there is a minor role of the
environment on this character as general except in crosses # 1 (Favi-9 x Edkawi) and # 3 (Favi-9 xStrain-B). This indication has been confirmed by the less number of minimum gene value obtained in Table 2. These results are coincided with those obtained by Hassan et al. (1984) who estimated BSH as 85 and 62% for TYLCV tolerance derived from of L. pimpinellifolium LA 121 and LA 373, respectively.
| Table 1: | Reaction of six-population individuals of five crosses against natural TYLCV infection during 2002-2004 at ARC, Research Station |
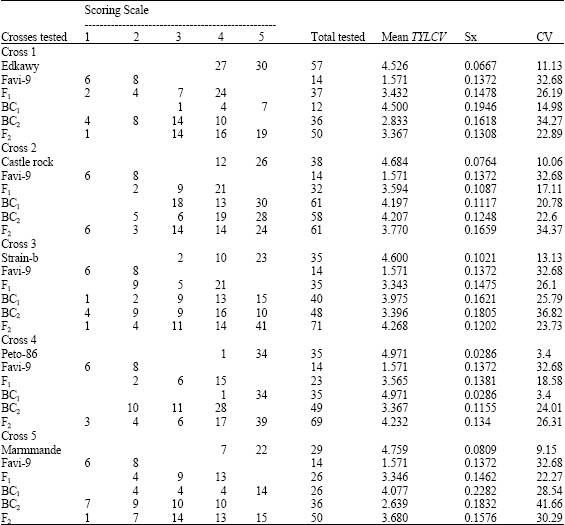 | |
| Sx-= Stander error; CV = Coefficient of variance | |
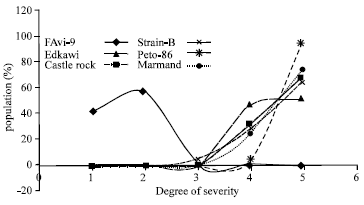 | |
| Fig. 1: | Reaction of susceptible and tolerant tomato parents against natural TYLCV infection at ARC Research Station during 2002 |
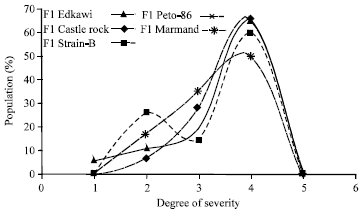 | |
| Fig. 2: | Reaction of F1 population of Different 5 crosses against natural TYLCV infection at ARC Research Station during 2003 |
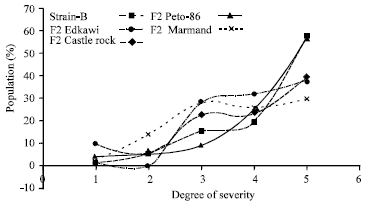 | |
| Fig. 3: | Reaction of F2 population of 5 crosses against natural TYLCV infection at ARC Research Station during 2004 |
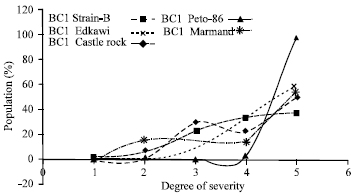 | |
| Fig. 4: | Reaction of BC1 population of different 5 crosses against natural TYLCV infection at ARC Research Station during 2003 |
The values of inbreeding depression (ID) are presented in Table 2. Those values are -17.7, -4.82, -27.66, -18.7and -9.98 in all five crosses, respectively (Table 2). Those large amounts of inbreeding depression for TYLCV trait in all crosses have been expected because of the large degree of heterosis showed, in this study, indicating presence of non-additive gene action (Allord, 1960).
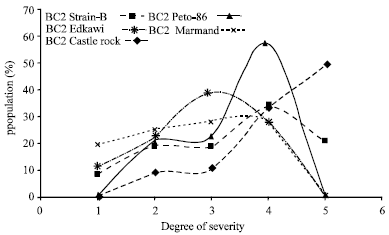 | |
| Fig. 5: | Reaction of BC2 population of different 5 crosses against natural TYLCV infection at ARC Research Station during 2004 |
| Table 2: | Genetic components i.e., heterosis, Board Sense Heritability (BSH) potency ratio, inbreeding depression and minimum number of genes for TYLCV tolerance in all crosses |
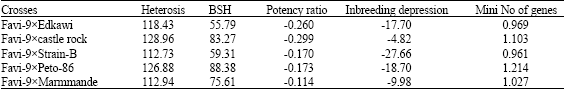 | |
One pair of genes could be estimated in crosses # 1 (Favi-9x Edkawi) and # 3 (Favi-9x Strain-B) (Table 2). Two pair of genes could be estimated in crosses # 2 (Favi-9xCastle Rock), # 4 (Favi-9xPeto-86) and #5 (Favi-9 xMarmmande) using the equation of Castle and Wright (1921) (Table 2). One pair of genes finding is coincided different reports (Kasrow, 1989; Laterrot, 1990; Pilowsky and Cohen, 1974) which indicated that TYLCV-resistance/tolerance is controlled by one pair of genes. Two pair of genes finding agree with those obtained by Banerjee and Kalloo (1987) who estimated that 2 pairs of genes are controlling this character. Mean while, Vidavisky and Czosnek (1998) reported that TYLCV-resistance is controlled by 2 to 3 pairs of genes. Also Nainar and Pappioha (2002) found that TYLCV-resistance in L. hirsutum is controlled by 3 pairs of genes. Vidavsky and Czosnek (1998) indicated that analysis of segregation of susceptibility, tolerance and resistance during the BC1F1 to BC1F4 crosses indicated that tolerance is controlled by a dominant major gene and resistance is controlled by 2 to 3 additive recessive genes.
ACKNOWLEDGEMENT
MERC project # M21-037 is acknowledged for supporting this study.
REFERENCES
- Banerjee, M.K. and Kalloo, 1987. Inheritance of tomato leaf curl virus resistance in Lycopersicon hirsutum f. glabratum. Euphytica, 36: 581-584.
CrossRefDirect Link - Czosnek, H. and H. Laterrot, 1997. A worldwide survey of tomato yellow leaf curl viruses. Arch. Virol., 142: 1391-1406.
CrossRefPubMedDirect Link - Ghanim, M., S. Morin, M. Zeidan and H. Czosnek, 1998. Evidence for transovarial transmission of tomato yellow leaf curl virus by its vector, the whitefly Bemisia tabaci. Virology, 240: 295-303.
CrossRefDirect Link - Hassan, A.A., H.M. Mazyad, S.E. Moustafa, S.H. Nassar, W.L. Sims and M.K. Nakhla, 1984. Genetics and Heritability of Tomato Yellow Leaf Curl Virus Tolerance Derived from L. pimpinellifolium. European Association for Research in Plant Breeding. Tomato Working Group, Wageningen, The Netherlands, pp: 81-87.
- Muniyappa, V., S.H. Jalikop, A.K. Saikia, Chennarayappa, G. Shivashankar, A.I. Bhat and H.K. Ramappa, 1991. Reaction of lycopersicon cultivars and wild accessions to tomato leaf curl virus. Euphytic, 56: 37-91.
CrossRef - Vidavisky, F. and H. Czosneki, 1998. Tomato breeding lines resistance and tolerant to tomato yellow leaf curl virus issued from Lycopersicon hirsutum. Phytopathol., 88: 910-919.
Direct Link