
R. K. Gupta
Division of Entomology, Faculty of Agriculture Sher-e-Kashmir University of Agricultural Sciences and Technology, Jammu, 180002, India
J. C. Raina
Division of Entomology, Faculty of Agriculture Sher-e-Kashmir University of Agricultural Sciences and Technology, Jammu, 180002, India
R. K. Arora
Division of Entomology, Faculty of Agriculture Sher-e-Kashmir University of Agricultural Sciences and Technology, Jammu, 180002, India
K. Bali
Division of Entomology, Faculty of Agriculture Sher-e-Kashmir University of Agricultural Sciences and Technology, Jammu, 180002, India
International Journal of Virology
Year: 2007 | Volume: 3 | Issue: 2 | Page No.: 45-59







ABSTRACT
Helicoverpa armigera (Hubner) is a serious pest of tomato in hill and mountain ecosystem of North Western India. Larvae of this species are susceptible to a Nucleopolyhedrovirus (NPV) which has attracted interest as a potential biocontrol agent. Three strains of NPV were isolated from infected larvae of host insect. The comparative pathogenecity/speed of kill of various isolates was determined in term of median Lethal Dose (LD50) and median Lethal Time (LT50) in a bioassay by diet plug method to select a candidate isolate for field potential in tomato and chickpea against Helicoverpa armigera (Hubner). The order of virulence of each isolate was Samba>Udheywalla>Chenani on 3rd instar larvae with respective LD50 values of 89, 243 and 287 OBs/larva, For all the isolates, the LT 50 values were inversely related to doses and ranged from 134.4 to 175.2 h at 3 different concentrations of 1000, 5000 and 25000 OBs/larvae. The samba isolate was considered about 2.73 to 3.22 times more virulent than Udheywalla and Chenani, respectively. Existence of differential virulence was also revealed in pot experiments wherein the highest possible mortality (98.33%) in 3rd instar larvae was recorded in Samba isolate of HaNPV followed by Udhyewalla (86.11%) and Chenani (81.66%) after 9 days of inoculation, the difference being significant. The field effectiveness of the most virulent isolate (samba) was assessed on tomato and chickpea-the two major host crops of this pest. Application of virus alone @ 3.0x1012 OBs ha-1 in tomato and 1.5x1012 OBs ha-1 in chickpea resulted in significant suppression of pest, lowered the per cent fruit /pod damage and led to the desirable increase in marketable yields. The candidate virus was found compatible with the recommended insecticide endosulfan, Bacillus thuringiensis and egg parasitoid, Trichogramma pretiosum in combined as well as sequential manner. It was concluded that ecofriendly management of this pest on these crops is possible either by use of selected virus alone or their combined application with insecticides at reduced doses.
PDF Abstract XML References
How to cite this article
R. K. Gupta, J. C. Raina, R. K. Arora and K. Bali, 2007. Selection and Field Effectiveness of Nucleopolyhedrovirus Isolates Against Helicoverpa armigera (Hubner). International Journal of Virology, 3: 45-59.
DOI: 10.3923/ijv.2007.45.59
URL: https://scialert.net/abstract/?doi=ijv.2007.45.59
DOI: 10.3923/ijv.2007.45.59
URL: https://scialert.net/abstract/?doi=ijv.2007.45.59
INTRODUCTION
The bollworm, Helicoverpa armigera Hubner (Lepidoptera: Noctuidae) is a polyphagous pest of worldwide occurrence inflicting annual crop damage in India worth US $1 billion. The problem of this pest is magnified due to its direct attack on fruiting structures, voracious feeding habits, high mobility and fecundity, overlapping generations with facultative diapause, nocturnal behaviour, migration, host selection by learning and genetic variability on different hosts (Reed and Pawar, 1982; Cunningham et al., 1999; Sharma, 2005; Subramanian and Mohankumar, 2006). In recent years, it has assumed such a serious proportion in India that for the past one decade farmers and plant protection community have been virtually driven mad by this monstrous pest, creating an array of social, economical and political problems leading to several suicidal deaths in southern and western region. Although, nearly 30% of total insecticides are used for controlling this pest alone on different crops yet many of them do not prove effective as pest has been reported to have developed resistance to almost all kinds of insecticides to varying folds (Armes et al., 1996 ; Yaqoob et al., 2006) resulting into build up of some minor pests and environmental degradation. Transgenic plants having Bt and other genes at one stage appeared to be the answer to all the problems posed by this pest in different countries but before such plants could reach the field from laboratories, the insect developed resistance to them (Ferré and Van Rie, 2002 ; Tabashnik et al., 2003). Therefore, the demand in the present day scenario is the formulation of some eco-friendly means of pest control to minimize pesticide related problems and to ensure long term sustainable yield through sound ecological principles. Fortunately, there are many natural enemies of this pest, which ultimately regulate their population in nature. Among them, the insect viruses are of immense utility (Ignoffo and Couch, 1981). Owing to their often pronounced host specificity and their high virulence to susceptible insect hosts, baculoviruses are considered to be one of the most efficient biocontrol agents for insects. The virions are occluded in olyhedral Occlusion Bodies (OBs) and are protected against environmental conditions for years. Today at least 1300 insect viruses are known to infect insects (Martignoni and Iwai, 1986; Murphy et al., 1995; Moscardi, 1999). Among these, Nuclear Polyhedrosis Viruses (NPVs) are the most suitable and effective pest control agents due to their desirable attributes like infectivity, lethality, storage stability and environmental safety (Cunningham, 1995). They are particularly attractive as bio-insecticides because of two factors. They are safe for vertebrates and other non-target fauna and they are generally highly pathogenic, host death being the most likely outcome of an infection. Today several baculoviruses have been commercialized all over the world and used as possible alternative for toxic, environmentally disruptive chemicals. However, despite present and past successes, there is a continuing need to discover and develop new strains/races in respect of increased virulence, tolerance to photo inactivation and greater persistence on phylloplane. As the selection of virulent strain of NPV is key to the development of effective bioinsecticides, local strains are always preferred for sustainability, adaptability and efficacy under a given set of agro-ecosystem and hold an ample scope for their wide spread multiplication and commercial use as novel biopesticides in a particular region (Escribano et al., 1999; Gupta et al., 2007). Phenotypic variation among different isolates of insect viruses recovered from members of the same host species from different geographical locations has been reported in terms of survival time and pathogencity and their DNA differed in restriction endonuclease and protein patterns (Gettig and McCarthy, 1982; Shapiro and Robertson, 1991; Toprak et al., 2006). In addition, it is well recognized that factors such as the geographic origins of both virus and host can affect the characteristics of the dose-response curve and the period of survival of infected hosts (Maeda et al., 1990; Milks, 1997; Figueiredo et al., 1999). The specific objective of this research was to select an NPV isolate for use against H. armigera in a control program designed for small-scale tomato growers in northwestern region of India with varied climatic zones and to assess its field potential in tomato.
MATERIALS AND METHODS
Rearing of Host Insect
Laboratory rearing of Helicoverpa armigera was started by collecting the adults in light trap, which were allowed to mate and oviposit in a cage of cylindrical iron frame (50x30 cm diameter) covered with black muslin cloth for egg laying. The moths were fed on 10% sucrose solution. The eggs bearing cloth was removed, surface sterilized with 10% formalin for 10 min, rinsed in distilled water five times for 10 min for each working and replaced daily with new one. The cloth containing eggs was then dried and cut into small pieces which were placed on moist cotton in 500 mL conical flasks, the mouths of which was wrapped with aluminum foil. The phototropic neonate larvae after hatching were then transferred to semi synthetic diet. The same disease free culture was maintained in laboratory at 26±2°C and 70±10% relative humidity, 10:14 h (L:D) photoperiod.
Virus Isolates, Amplification and Enumeration of OBs
All of the isolates used in this study were originally collected from infected H. armigera larvae from traditionally pesticide free areas (Gupta, 2004). The isolates were given serial passages in the homologues host before propagation for further studies. Each isolate was propagated in 3rd instar larvae for further studies following adherence to strict microbiological procedures by harvesting the cadavers immediately after death to minimize bacterial contamination. Only 1 virus isolate was produced at a time to prevent cross-contamination Larvae that remained intact were only harvested and macerated with 3 mL of 0.1% Sodium Dodecyl Sulphate (SDS) and filtered through three-layered muslin cloth several times to remove the insect debris. Further, purification was done by centrifugation of filtrate at 500 rpm for 1 min and discarding the pellet of tissue debris. The supernatant thus obtained was subjected to centrifugation at 2500 rpm for 5 min and supernatant containing lipids and soluble material was discarded. The resuspended pellet containing the occlusion bodies was further purified and the fraction containing POBs was diluted in appropriate quantity of 0.1% SDS, pelleted and washed in sterile distilled water. Prior to enumeration, the contents were vortexed to facilitate equal distribution of polyhedra in the suspension and the stock suspension was diluted in sterilized distilled water to the desired level using a micropipette. Ten microliter of the viral suspension was expelled into a haemocytometer chamber and was left undisturbed for 2 min. Counting of POBs was done under a phase contrast microscope (Nikon Eclipse 600) using doubled ruled Neubauer Haemocytometer (Weber scientific) with a 0.1 mm depth. Ten replicated counts were made systematically across the grid after appropriate settings for each sample. The quantified stock suspensions were stored at 4°C for further use.
Bioassays
The comparative pathogenecity/speed of kill of various isolates was determined in term of median Lethal Dose (LD50) and median Lethal Time (LT50) in a bioassay by diet plug method. For viral inoculation, a modified diet without formaldehyde was prepared. A cube of diet, calculated to be consumed in 24 hrs was contaminated with a specific amount of virus i.e., 0 (control), 25, 50, 100, 200, 300, 400, 500, 1000, 2000 and 3000 OBs in each container. These suspensions were made in sterile distilled water to which 0.1% spreader was added. A newly molted third instar larvae starved for 24 h was then placed individually in 50 mL sterilized plastic containers .The larvae that consumed the entire plug within 24 hrs were then transferred to clean uncontaminated diet free of formaldehyde and incubated for the required period. Larvae were monitored at 24 h interval for mortality due to HaNPV infection until pupation. Thirty larvae were used for each dose and entire experiment was replicated three times. Dose-mortality data and median Lethal Times (LT50) were evaluated with the computer program SPSS 10.0 which is based on the probit analysis method. Differences in the slopes of regression lines and the non-overlapping of the 95% Confidence Limits (CL) were considered as evidence of significant statistical differences among the values being evaluated.
Laboratory Pot Experiment
A laboratory pot experiment was conducted to evaluate the efficacy of each isolate on tomato at virus concentration of 3x1012 OBs ha-1. A single replicate comprised of five plants of tomato var. Pusa Ruby (6 week old) encaged in acrylic cages (2x2 x2 ft) provided with mesh for aeration. The virus was sprayed on the plants uniformly with hand sprayer along with adjutants like Tinopal (0.1%) and jaggery (1%). The larvae were released on the plants immediately after drying of spray fluid on plants. The larval mortality was OBserved after 5, 7 and 9 days of application. Each experiment was repeated five times and mean larval mortality at different intervals was compared with one way ANOVA, tukey HSD test.
Field Efficacy
Small-scale field trials were conducted on tomato and chickpea to generate the efficacy data of most virulent samba isolate alone and in combination with the recommended biopesticides/bioagents against H. armigera in tomato. The first trials (Trial I) was conducted on tomato hybrid Sonali during spring 2002-2003 in Randomized Block Design (RBD) comprising 5 treatments, each replicated four times in plots of (3x2.5 m).The Standard treatments were Bacillus thuringiensis ar Kurstaki, Biobit WP; Rallis India Ltd. Bombay, India), Helicoverpa armigera NPV (HaNPV local Samba strain) and Trichogramma pretiosum (Biotech International Ltd., New Delhi). Since, in the first trial we could not bring the fruit damage below 10% as desired by local farmers, an another trial on potential implication of integration of virus with bioagents/safer recommended pesticide in combined/sequentia manner (Trial II) was conducted in the following season at Research Farm, Faculty of Agriculture, Udheywalla on tomato.Twelve treatments were assigned to three replicates in randomized block design in plots of 3 x 2.5 m. Similarly, an additional trial (Trial III) was also conducted on chickpea variety C-235, which is another major host of H. armigera in the prevailing agro ecosystem during the same year. Since, Trichogramma do not work in chickpea eco-system the number of treatments was reduced to eight from 12 as in Trial II. All insecticides/biopesticides tested in this study are registered in India.
Application
In all trials applications were made with a Knapsack sprayer equipped with a single cone nozzle. The spray was applied evenly along each row of plants from a height of approx. 40 cm. Virus treatments were applied using a clean sprayer to avoid cross contamination. The spraying was done at evening hours to protect NPV from sunlight as well as to avoid any adverse effect of pesticide on natural enemies. Adjutants like Tinopal (0.1%) and jaggery (1%) were added to NPV before spraying. In the treatments comprising combination of biopesticides, single spray was conducted by mixing two components. The egg parasitoid Trichogramma pretiosum was released 6 h after spraying biopesticides and a screen of muslin cloth was provided in the periphery of each treated plot in order to confine its effect in the same plot.
Observations
The data on larval density were recorded by randomly selecting 5 plants from alternate rows of each application before one day of treatment and after 7 and 14 days of treatment. Per cent fruit damage was calculated at each picking and pooled. Yields were determined by weighing all fruits at each picking from each plot.
Statistical Analysis
The data recorded were statically analyzed after performing appropriate transformations according to SPSS 10.0 version.
RESULTS
Biological Activity
Dose-Mortality Response
All the three tested isolates were found highly infectious to H. armigera with differential degree of virulence (Table 1). The value of χ2 did not differ significantly indicating the non heterogenous response for all the isolates.
| Table 1: | Dose mortality responses and LD50 values (OBs/larvae) of various isolates for third instar larvae of H. armigera |
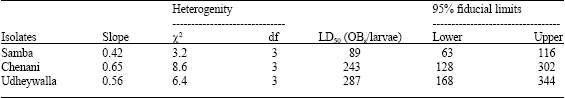 | |
| 30 larvae per isolate were exposed per virus dose. Bioassays were replicated three times.Data were analysed by probit analysis at α = 0.05 confidence level and fit probit model by χ2 tests at α = 0.05 | |
| Table 2: | LT50 values for 3rd instar H. armigera larvae infected with different doses of three isolates of HaNPV |
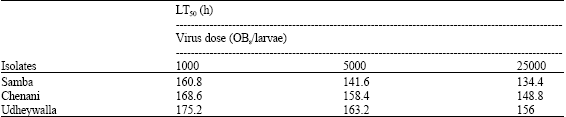 | |
| 30 newly ecdysed third-instar H. armigera is larvae were inoculated using a diet plug method. After the inoculation, mortality from NPV was examined every 6 h until death of larvae. Mortality data were analysed using probit analysis. Bioassays were replicated three times | |
| Table 3: | Efficacy of various indigenous isolates of [email protected] 12 against 3rd instar larvae of Helicoverpa armigera in pot experiments* |
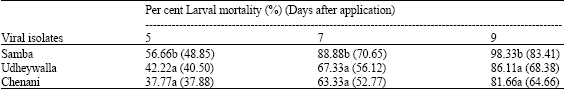 | |
| *Experiment was replicated five times, each replicate comprised of 5 plants encaged in acrylic cages in which 100 larvae @ 20/plant were released after treatment. Figures in the parentheses are angular transformed values. Within a column means followed by similar letters do not differ significantly, ANOVA, Tukey HSDa (p<0.05) | |
The slope values varied from 0.42 to 0.65. Samba isolate exhibited the lowest slope value reflecting less uniform response to the tested larva to virus isolate. According to the dose-mortality bioassays, the LD50 values of Samba, Udheywalla and Chenani isolate were 89, 243 and 287 OBs/larva, respectively, with non overlapping confidence limits for samba isolates in comparision to two other isolates. As such pathogenecity of samba isolate was considered significantly different from other isolates and it was considered about 2.73 to 3.22 times more virulent than Udheywalla and Chenani isolates, respectively.
Time-Mortality Response
Median Lethal Times (LT50) were calculated for each isolate as an indicator of their speed of kill. The LT50 values calculated for all isolates at 3 different concentrations of 1000, 5000 and 25000 OBs/larvae produced 100% mortality for both third-instar larvae. For all the isolates, the LT50 values were inversely related to doses and ranged from 134.4 to 175.2 h (Table 2). Based on a non-overlap of 95% confidence limits and the slopes of regression lines, Samba isolate was considered significantly superior over the other two isolates as it resulted in 50% mortality of third instar larvae of H armigera in comparatively short period. However, the differences among other two isolates were non significant.
Pot Experiment
Existence of differential virulence in indigenous strains from 3 different places was also indicated by varied larval mortality in pot experiments (Table 3). It was found that the order of virulence of each isolate was Samba>Udheywalla>Chenani on fifth, seventh and ninth days of inoculation.
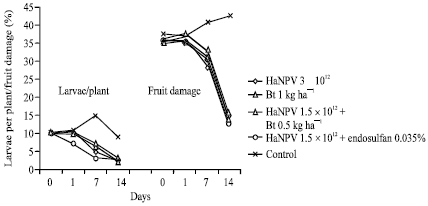 | |
| Fig. 1: | Larval density and fruit damage in tomato as influenced by various treatments at 0, 1, 7 and 14 days of application |
The highest possible mortality (98.33%) in 3rd instar larvae was recorded in Samba isolate of HaNPV followed by Udhyewalla (86.11%) and Chenani (81.66%) after 9 days of inoculation, the difference being significant (p<0.05, F = 35.11). This isolate also caused significantly higher larval mortality as compared to other isolates on fifth and seventh day after inoculation. Therefore, Samba isolate was considered most virulent among all and multiplied in laboratory for testing its efficacy in the field after standardization of OBs count mL-1.
Field Evaluation
Small-scale field trials were conducted at two locations viz., Udheywalla and Satwari to generate the efficacy data regarding potential utilization of selected isolate alone and in combination with the recommended insecticides, biopesticides and bioagents against H. armigera in tomato.
Trial 1
Trial 1 was conducted at farmer’s field at Satwari wherein all the treatments were targeted against high-density larval population of pest on hybrid tomato variety namely Sonali. While pretreatment population was almost uniformly distributed in all the treatments (Fig. 1), a significant reduction in larval density was OBserved after 7 and 14 days of application, the maximum being offered by the combined application of virus @ 1.5x1012 OBs ha-1+ 0.035% endosulfan followed by that of virus applied alone at full dose (3x1012 OBs ha-1). Similarly, It was found that the use of indigenously developed bio-pesticide at full doses or its combined application at reduced doses along with other bio-agents could provide significant protection to the fruits of tomato. While fruit damage continued to increase from 38.3-44.3% in untreated control, following treatment applications it declined remarkably from 37.5 to 15.5%. The maximum marketable yield of 28.75 kg plot-1 (3.0x2.5 m) was also obtained with combined application of virus @ 1.5x1012 OBs ha-1 + 0.035% endosulfan followed by that of virus applied at full dose (3x1012 OBs ha-1). The increase in marketable yield in all the treatments over control ranged from 60.70 to 101.70% (Fig. 2).
Trial II
In trial II, population of pest was almost uniformly distributed in all the treatments before spray application on the most commonly grown cultivar, Pusa Ruby. Following the treatment applications the larval density followed a declining trend in both the sprays (Table 4). After 24 h of treatment application the maximum reduction in pest population was OBserved in T6 (Endosulfan @ 0.07%) and T8 (NPV 1.5x1012+endosulfan @ 0.035%), whereas rest of the treatments were at par with the control (p<0.05, F = 5.42).
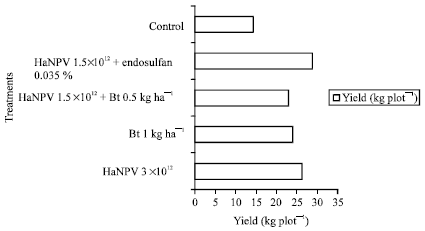 | |
| Fig. 2: | Marketable yield of tomato fruits as influenced by various treatment applications. *Pooled mean at various pickings |
| Table 4: | Field Efficacy of selected virulent Samba isolate of HaNPV alone and in combination with bioagents endosulfan on larval population in tomato |
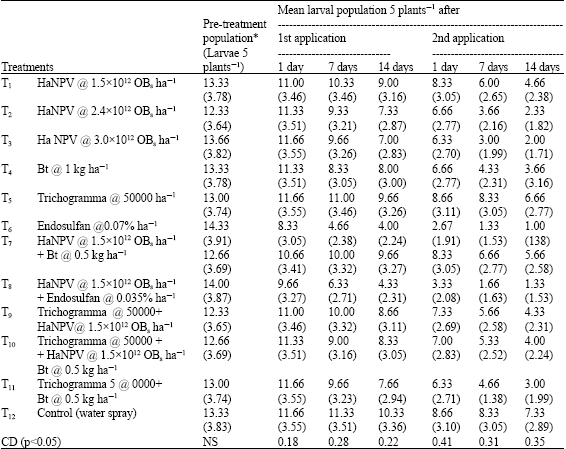 | |
| *Figures in the parentheses are √n+1 transformation; #HaNPV Helicoverpa armigera nuclear polyhedrosis virus; NS = Non Significant | |
The larval population recorded after 7 and 14 days of first treatment varied from 4.66 to 11.33/five plants and once again these two treatments proved more effective in comparison to rest of the treatments. After 14 days of 2nd treatment application, the plots sprayed with endosulfan 0.07% (T6) recorded least larval population (1 larva/5 plants), followed by plots sprayed with combined application of 1.5x1012 OBs ha-1 + endosulfan 0.035% (T8).
| Table 5: | Effect of of selected virulent Samba isolate of HaNPV alone and in combination with bioagents/endosulfan on fruit damage yield of tomato |
 | |
| Figures in the parentheses are arc sin percentage transformation; #HaNPV Helicoverpa armigera nuclear polyhedrosis virus; = *Plot size 3x2.25 m | |
However, both these treatments were statistically at par with each other. The treatments comprising the single release of T. pretiosum @ 50000/ha (T5) and HaNPV 1.5x1012 + Bt @ 0.5 kg ha-1 (T7) proved ineffective in checking the larval population even after 7 (p<0.05, F = 22.51) and 14(p<0.05, F = 14.77) days of 2nd treatment application.
Fruit Damage
Data analysis on fruit damage (Table 5) revealed that all the treatments of HaNPV alone or in combination with bioagents/endosulfan proved effective in checking the per cent fruit damage caused by the pest in tomato (p<0.05,F = 6.99). The lowest fruit damage (6.20%) was OBserved in plots treated with endosulfan 0.07% which was however at par with the plots treated with HaNPV @1.5x1012 OBs ha-1 + endosulfan 0.035% (7.4%), HaNPV @ 3x1012 OBs ha-1 (8.20%) and HaNPV @ 2.4x1012 OBs ha-1(9.13%). The other treatments also gave significant protection against fruit damage by this pest, but OBserved degree of protection was significantly low over above treatments
Marketable Fruit Yield
Maximum marketable fruit yield (15.24 kg plot-1) was recorded in the plots treated with endosulfan 0.07% followed by plots treated with HaNPV @1.5x10 12 OBs ha-1 + endosulfan 0.035%, HaNPV @ 3x1012 OBs ha-1 (14.09 kg plot-1) and HaNPV @ 2.4x1012 OBs ha-1 (13.18 kg plot-1). However, all these treatments were significantly at par with each other (p<0.05, F = 14.77). The rest of the treatments viz., HaNPV alone @1.5x1012 OBs ha-1, Bt alone (T4) and in combination with NPV (T7) and Trichogramma alone (T7) and in combination with biopesticides (T9, T10 and T11) although led to increase in the marketable yield over control yet their impact was significantly inferior to other treatments (Table 5).
Trial-III
Larval Population
The larval population recorded on 5 randomly selected plants of chickpea before one day of treatment application showed non-significant difference among all treatments (p<0.05, F = 0.788). However, after 1, 7 and 14 days of treatment applications, significant differences in terms of larval density were OBserved among various treatments (Table 6 ). After 24 h of 1st treatment application lowest larval density (4.33/5 plants) was OBserved on plots sprayed with endosulfan (0.07%) which was at par with the plots sprayed with combined application of HaNPV @ 1.5x1012 OBs ha-1 and endosulfan 0.035% (4.66/5 plants) whereas other treatments were statistically at par with control (p<0.05, F = 8.96).
| Table 6: | Effect of selected virulent samba isolate of HaNPV and in combination with bioagents endosulfan on larval population in chickpea. |
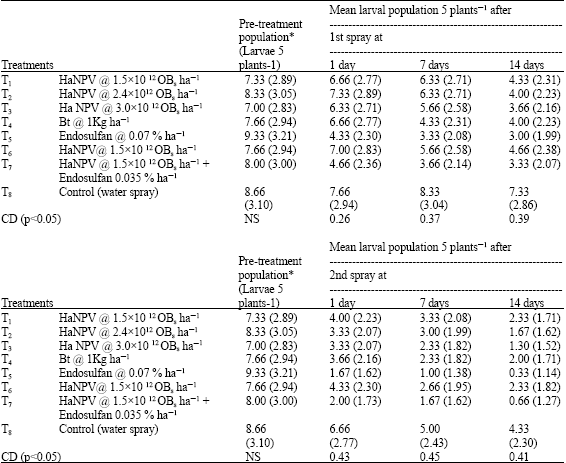 | |
| *Figures in the parentheses are √n+1 transformation; #HaNPV Helicoverpa armigera nuclear polyhedrosis virus; NS = Non Significant | |
The larval population recorded after 7 days of treatment varied from 3.33 to 8.33/5 plants and endosulfan 0.07%, HaNPV @1.5x10 12 OBs ha-1 + endosulfan 0.035%, HaNPV alone @ 3.0x1012 OBs ha-1, Bt 1 kg ha-1 and HaNPV alone @ 1.5x1012 OBs ha-1 + Bt 0.5 kg ha-1 proved more effective in suppressing the pest population in comparison to rest of treatments (p<0.05, F = 5.92). However, after 14 days, all the treatments proved significantly superior over control in checking the larval population and least larval population was found on the plots sprayed with endosulfan 0.07% (3 larvae/5 plants) and HaNPV @ 1.5x1012 OBs ha-1+ endosulfan 0.035% (3.33 larva/5 plants) (p<0.05, F = 4.66). More or less similar trend was formed in the 2nd spray and least larval population was recorded on the plots sprayed with endosulfan 0.07% (0.33 larvae/5 plants) and HaNPV alone @ 1.5x1012 OBs ha-1 + endosulfan 0.35% (0.66 larval/5 plants), which were statistically at par after 14 days of treatment application (p<0.05, F = 6.82).
Pod Damage
Data analysis on pod damage (Table 7) revealed that all the treatments of HaNPV alone or in combination with bioagents/endosulfan proved effective in checking the per cent pod damage caused by the pest in chickpea (p<0.05, F = 9.94). Lowest pod damage (6.60%) was OBserved in plots treated with endosulfan 0.07%, which was however, at par with the plots sprayed with HaNPV @1.5x1012 OBs ha-1+endosulfan 0.035% (7.93%) and HaNPV alone @ 3.0x1012 OBs ha-1 (11.73%).
| Table 7: | Effect of selected virulent Samba isolate of HaNPV alone and in combination with bioagents/endosulfan on pod damage and grain yield of chickpea |
 | |
| *Figures in the parentheses are arc sin percentage transformation; #HaNPV Helicoverpa armigera nuclear polyhedrosis virus; *Plot size 3x2.25 m | |
Yield
Maximum grain yield (1.223 kg plot-1) was recorded in the plots treated with endosulfan (0.07%) which was however, at par with yield (1.133 kg plot-1) obtained from the plots treated with HaNPV @ 1.5x1012 OBs ha-1 + endosulfan 0.035% and HaNPV alone @ 3.0x1012 OBs ha-1 (Table 7). The yield obtained in other treatments viz., HaNPV alone @ 2.4x1012 OBs ha-1 , HaNPV 1.5x1012 OBs ha-1, Bt @ 1 kg ha-1 and HaNPV alone @ 1.5x1012 OBs ha-1 + Bt @ 0.5 kg ha-1 were significantly superior over control, but proved inferior to the best treatments (p<0.05, F = 6.10).
DISCUSSION
The increased potential of microbial control is attributed to the discovery and development of new species and strains of entomopathogens. However, despite the successes of the past there is continuing need to discover and develop new strains/races if we are to meet the future needs of food and fibre production and concomitant reduction in the use of chemical pesticides (Hokkanen and Pimentel, 1984). There is no denying the fact that many insect viruses have already been isolated and used successfully (Moscardi, 1999) but the selection of virulent strain of NPV is key to the development of effective viral insecticides (Fuxa, 1991). In general regional viral strains with desirable traits hold an ample scope to overcome their disadvantages, such as their relatively slow speed of killing and narrow host range (Shapiro and Robertson, 1991; Blissard and Rohrmann, 1990; Escribano et al., 1999). This can either be achieved either by cloning the viruses by plaquing from uncloned wild populations that generally exhibit natural genetic heterogeneity (Lynn et al., 1993) or by isolation and selection of native isolates (Escribano et al., 1999; Toprak et al., 2006). In the present study, the comparative biological activity of the isolated strains was expressed in term of both LD50 and LT50 values which allowed direct comparision of their relative virulence among themselves or previously isolated strains. It was found that all the three different isolates of HaNPV exhibited differential degree of pathogenecity. The order of virulence of each isolate was Samba>Udheywalla>Chennai in term of both LD50 and LT50 values. The OBserved values are of more or less similar magnitude to those already reported earlier (Rabindra et al., 2003; Figueiredo et al., 1999). The occurrence of geographic variants has been well demonstrated in nuclear polyhedrosis virus of Heliothis sp. and several single and multiple NPV (HearMNPV and HearSNPV) have been isolated in different geographical regions from H. armigera larvae (Kelly et al., 1980; Gettig and McCarthy, 1982; McIntosh and Ignoffo, 1983; Sudhakar and Mathavan, 1999). Some of these isolates showing different DNA Restriction Endonuclease (REN) profiles also differed significantly in their insecticidal activity ( Hughes et al., 1983 ; Williams and Payne, 1984). Such studies generally have indicated that geographical variants are related strains of the same virus that show limited differences in the presence and distribution of restriction enzyme cleavage sites. However, minor genetic differences can have biologically significant consequences in terms of the phenotypic characteristics of virus (Blissard and Rohrmann, 1990; Possee and Rohrmann, 1997). These OBserved differences in biological activity are not unusual among virus isolates from distinct geographical regions (Hughes et al., 1983; Shapiro and Robertson, 1991; Caballero et al., 1992; Figueiredo et al., 1999) or among clonal variants derived from a single wild-type virus (Lynn et al., 1993). Even very small structural differences in DNA or Polypeptide can result in very large differences in virus infectivity (Monroe and McCarthy, 1984). This is attributed to the wide variety of key genes (Zhang et al., 2005) which are found in the polyhedrin gene region (Woo et al., 2006) or those involved in blocking host cell apoptosis (Clem, 1997) .
Once we were able to select a virulent isolate through lab and pot experiment, the next step is to validate its field potential through small scale trials conducted in different locations. One of the pre-requisite for the selection of an isolate as a biocontrol agent is that geographical variants be tested against an insect population from the locality in which the program is to be run (William et al., 1999). The successful utilization of this virus has already been established against this pest on various crops elsewhere in the country and abroad. However, our preliminary studies with commercially available strains of HaNPV did not bring desirable protection from pest in tomato and chickpea in prevailing ecosystem, probably due to the reason that these products were based on isolates obtained from southern region of country. The specific objective of this study was to select a naturally occurring native isolate for northwestern region of India from geographically varied location in terms of infectivity and virulence and to evaluate its field potential. The utilization of this virus under pot experiment showed its promising potential. Henceforth, the field experiments were undertaken to assess its field effectiveness. The prime aim of this study was to reduce the use of pesticides and to refine the application procedure for effective suppression under given set of agro-ecosystem coupled with the reduction of doses level of HaNPV/bioagents for cost effectiveness. The first trial was conducted in order to assess the field potential of virus alone at two different doses i.e., 1.5x1012 and 3x1012 OBs ha-1 in comparision to the recommended insecticide in tomato. Although its use alone at both the doses resulted in significant reduction in pest attack and increase in yield over control, yet the fruit damage could not be lowered below 10 %-a feature desired by progressive farmers in the vicinity. Singh (2001) have advocated the 6 weekly releases of T. pretiosum @ 50000/ha followed by the three applications of HaNPV @ 250 LE/ha for successful management of this pest in tomato. However, preliminary OBservations under given ecosystem reflected that soon after the appearance on tomato, abrupt increase of pest is recorded within 15-20 days. Therefore, we planned for the application of virus alone as well as in combined sequential manner with the safer insecticides bioagents in following season. The chemical pesticides were mixed along with the biopesticides at full doses, keeping in view their different periods of efficacy and to minimize the number of sprays required for the management of this pest as the pesticidal pressure has already been exerted on the pest in the tomato and chickpea growing areas of the state (Yaqoob et al., 2006).The validation of this technique as advocated by Singh (2001) with some refinement resulted in significant suppression of pest, lowered the per cent fruit damage and led to the desirable marketable yield of tomato besides, reducing the application cost for the bioagents. The suppression level of pest obtained in respect of reduction in larval population and fruit damage and increase in tomato yield in this investigation is comparable to the OBservations of (Pokharkar and Chaudhary, 2001) and Srinavasan et al. (1994). The use of virus alone @ 3x1012 OBs ha-1 or its sequential application after release of T. pretiosum @ 50000/ha also gave satisfactory results. This finding suggests that these bioagents can be effectively used for organic production of tomato. Similar protection was OBserved on chickpea wherein all the treatments of HaNPV alone or in combination with bioagents/endosulfan proved effective in checking the larval density and percent pod damage and increase in grain yield as demonstrated by Cherry et al. (2000) and Bhagwat and Wightman (2001). In general, it was found that the treatments comprising of endosulfan alone or in combination with HaNPV proved significantly superior to the treatments comprising bioagents alone or in combination with each other and gave maximum reduction in larval population, substantial reduction in pod/fruit damage and consequent increase in yield in chickpea and tomato. Bioagents alone or in combination with each other, although could reduce the larval population effectively after 7 and 14 days of spray, yet after one day of initial application, they proved inferior to endosulfan alone or the treatments comprising endosulfan in combination with bioagents. The OBserved differences in efficacy are attributed to the different modes of action of the products. As nerve poison, endosulfan is expected to have a relatively rapid action. The Bt toxin is a stomach poison, whose mode of action is to damage the cells of the insect gut, causing death through starvation. The mode of action of NPV is through systemic infection following ingestion of the virus particles. The virus replicates within all host cells leading to disintegration of the host tissues, culminating in death. The rate of this process is a function of the dose ingested, temperature and larval size, but can vary between 4 and 14 days (Jones et al., 1994). It is well established that the mortality due to NPV in field is initiated after 7 days which is incubation period for virus to replicate in host insect. However, when it is combined with the insecticide, the speed of kill is increased partially due to direct effect of insecticide or its sub lethal action or synergism (Geetha et al., 1999). This synergistic action is attributed to the suppression of activity of detoxifying enzymes such as Mixed Function Oxidases (MFO), Carobxylestrases (CarE) and Glutathione S-transferases. Since this pest is acquiring differential degree of resistance coupled with increased ratio of resistance through various generations (Yaqoob et al., 2006), this finding could be of vital importance to evolve resistance management strategy against this pest. However, a combination of NPV with Bt. did not give better results than their application alone. Santharam et al. (1993) ascribed it to differential mode of action by these microbes, which may compete for living cells for their multiplication replication. The present findings, suggests that ecofriendly management of this pest on these crops is possible either by use of selected viral strain alone or its combined application with insecticides at reduced doses. It was interesting to note that HaNPV even at 1.5x1012 OBs ha-1 was found to be effective in checking larval population in chickpea whereas in tomato crop HaNPV at higher doses i.e., 3.0x1012 OBs ha-1 was required to bring equitable results. The effectiveness of NPV depends upon the crop phenology and the factors like protein binding, pH, plant allelochemicals and passage time of food through the gut all might affect the susceptibility of insects to NPV on different hosts (Fuxa, 2004). As such varied rates of application on different crops have been recommended (Singh, 2001). The results also showed that pesticides, which are environment friendly and are compatible with the bioagents, should remain an important component of IPM.
ACKNOWLEDGMENTS
We are deeply grateful to Dr Nor chejonsky, Agriculture Research Organization, Israel for critically reading this manuscript and providing the helpful suggestions. This research was funded by the National Agriculture Technology Project, ICAR, India (project number CGPII-108).
REFERENCES
- Armes, N.J., D.R. Jadhav and D.K.R. Souza, 1996. A survey of insecticide resistance in Helicoverpa armigera in the Indian subcontinent. Bull. Entomol. Res., 86: 499-514.
Direct Link - Bhagwat, V.R. and J.A. Weightman, 2001. NPV based management of Helicoverpa armigera (Hub.) in chickpea (Cicer arietinum L.). Ann. Plant Prot. Sci., 9: 209-212.
Direct Link - Cherry, A.J., R.J. Rabindra, M.A. Parnell, N. Geetha, N.S. Kennedy and D. Grzywacz, 2000. Field evaluation of Helicoverpa armigera nucleopolyhedrovirus formulations for control of chickpea pod borer, H. armigera (Hubn.), on chickpea (Cicer arietinum var. Shoba) in Southern India Crop. Prot., 19: 51-60.
Direct Link - Cunningham, J.P., M.P. Zalucki and S.A. West, 1999. Learning in Helicoverpa armigera (Lepidoptera: Noctuidae): A new look at the behaviour and control of a polyphagous pest. Bull. Entomol. Res., 89: 201-207.
CrossRef - Escribano, A., T. Williams, D. Goulson, R.D. Cave, J.W. Chapman and P. Caballero, 1999. Selection of a nucleopolyhedrovirus for control of Spodoptera frugiperda (Lepidoptera: Noctuidae): Structural, genetic and biological comparison of four isolates from the Americas. J. Econ. Entomol., 92: 1079-1085.
Direct Link - Ferre, J. and J. Van-Rie, 2002. Biochemistry and genetics of insect resistance to Bacillus thuringiensis. Annu. Rev. Entomol., 47: 501-533.
PubMed - Hokkanen, H. and D. Pimentel, 1984. New approach for selecting biological control agents. Can. Entomol., 16: 1109-1121.
CrossRefDirect Link - Milks, M.L., 1997. Comparative biology and susceptibility of cabbage looper (Lepidoptera: Noctuidae) lines to a nuclear polyhedrosis virus. Environ. Entomol., 26: 839-848.
Direct Link - Moscardi, F., 1999. Assessment of the application of baculoviruses for control of Lepidoptera. Annu. Rev. Entomol., 44: 257-289.
PubMedDirect Link - Rabindra, R.J., N. Sathiah, K.P. Jayanth and R.K. Gupta, 2003. Improving Commercial Scale Production and Formulation Technology for Nuclear Polyhedrosis Viruses of Helicoverpa armigera and Spodoptera litura. In: Advances in IPM, Singh, A., T.P. Trivedi, H.R, Sardana, O.P. Sharma and N. Sabir (Eds.), National Centre for Integrated Pest Management., IARI., New Delhi, India, pp: 33-40.
- Sudhakar, S. and S. Mathavan, 1999. Electron microscopical studies and restriction analysis of Helicoverpa armigera nucleopolyhedrosis virus. J. Biosci., 24: 361-370.
CrossRefDirect Link - Tabashnik, B.E., Y. Carriere, T.J. Dennehy, S. Morin and M.S. Sisterson et al., 2003. Insect resistance to transgenic Bt crops: Lessons from the laboratory and field. J. Econ. Entomol., 96: 1031-1038.
Direct Link - Toprak, U., S. Bayram and O.M. Gurkan, 2006. Comparative biological activities of a plaque-purified variant and a Turkish native isolate of SpliNPV-B against Spodoptera littoralis (Lepidoptera: Noctuidae) Pest. Manage. Sci., 62: 57-63.
CrossRefDirect Link - Williams, T., D. Goulson, P. Caballero, J. Cisneros, A.M. Martinez, J.W. Chapman, D.X. Roman and R.D. Cave, 1999. Evaluation of a baculovirus bioinsecticide for small-scale maize growers in Latin America. Biol. Control, 14: 67-75.
CrossRefDirect Link - Woo, S.D., J.Y. Choi, Y.H. Je and B.R. Jin, 2006. Characterization of the Helicoverpa assulta nucleopolyhedrovirus genome and sequence analysis of the polyhedrin gene region. J. Biosci., 31: 329-338.
PubMedDirect Link - Zhang, C.X., X.C. Ma and Z.J. Guo, 2005. Comparison of the complete genome sequence between C1 and G4 isolates of the Helicoverpa armigera single nucleocapsid nucleopolyhedrovirus. Virology, 333: 190-199.
CrossRefDirect Link