
Lydie-Stella Koutika
Centre de Recherche sur la Durabilite et la Productivite des Plantations Industrielles (CRDPI), B.P. 1291, Pointe-Noire, Republic of the Congo
LiveDNA: 682.13136
George Ayaga
Kenya Agricultural and Livestock Research Organization, KALRO, P.O. Box 278, Busia, Kenya
Philip Brookes
Department of Soil Science, Rothamsted Research, Harpenden, Herts. AL5 2JQ, United Kingdom
International Journal of Soil Science
Year: 2017 | Volume: 12 | Issue: 3 | Page No.: 97-103







ABSTRACT
Background and Objective: Phosphorus (P) deficiency is mainly due to P occlusion in the iron and aluminum oxides in the highly weathered soils of many African countries. The objective of the study was to evaluate the impact of combined manure and inorganic fertilizer on soil P availability for plant uptake, enhanced soil microorganism activity and improved crop yields. Materials and Methods: To test whether increasing microbial biomass protects immobilized phosphorus from fixation and ultimately increases P availability to plants, 33P turnover 63 days and DNA concentrations were measured using the Multi terminal restriction fragment length polymorphism procedure in the two Kenyan soils (high and low P fixing soil) with 4 treatments: (1) control, (2) + Farmyard manure (FYM) (10g kg–1), (3)+ Superphosphate (g kg–1) and 33Phosphorus and (4) FYM+superphosphate (g kg–1) and 33Phosphorus. The one-way analysis of variance (ANOVA) was conducted on different treatments of both studied soils to evaluate 33P turnover and difference in DNA concentrations. Results: The 33P specific activity of Soil Microbial Biomass P (SMB-P) was much higher in the high P fixing soil (KH2PO4 and FYM+KH2PO4) than in the low P fixing soil at Day 21 and 63, showing an enhanced 33P turnover. DNA concentrations were higher in the low P fixing soil (>14 ng μL–1) relative to the high P fixing soil (<10 ng μL–1) at 63 days suggesting the difference in the microbial groups of the two studied soils. Combination of KH2PO4 and FYM benefited to P turnover and DNA concentrations of the high fixing Kenyan soil. Conclusions: This study highlighted the useful effects of combining manure with inorganic fertilizer to both reduce P adsorption and increase P availability i.e., improve soil fertility, to increase crop growth and yields in weathered soils.
PDF Abstract XML References Citation
Received: October 10, 2016;
Accepted: July 24, 2017;
Published: October 02, 2017
How to cite this article
Lydie-Stella Koutika, George Ayaga and Philip Brookes, 2017. Changes in Phosphorus Turnover and Microbial Communities of Two Kenyan Soils. International Journal of Soil Science, 12: 97-103.
DOI: 10.3923/ijss.2017.97.103
URL: https://scialert.net/abstract/?doi=ijss.2017.97.103
DOI: 10.3923/ijss.2017.97.103
URL: https://scialert.net/abstract/?doi=ijss.2017.97.103
INTRODUCTION
Soil microbial biomass is amongst the most labile fractions of Soil Organic Matter (SOM), represents about 2-3% of total SOM1 and drives the cycling of organic matter and nutrients in soil including P2-5. Soil microbial biomass is affected by the type and amount of soil inputs that farmers apply, whether in the form of synthetic fertilizer, organic residues, or a combination of the above two. Earlier studies showed that the absence or excess of P fertilization reduced the microbial biomass pool in soils from the South-Western Quebec6, while P concentration of soil solution reinforces P-limiting conditions7. To evaluate P turnover in different land use systems and management, the labelled 33P was commonly used8-10.
Terminal Restriction Fragment Length Polymorphism (T-RFLP) analysis is a sensitive and automated fingerprinting method which uses fluorescently digestion and analysis of terminal fragments with a DNA sequencer11.T-RFLP is used to study complex microbial communities based on the 16S-rRAN gene12. M-TRFLP is recommended when information for more than one taxon is required11. Soil microorganism population might be affected by organic and inorganic fertilizers in long-term experiments13,14, while bacterial community shifts might not occur in the short-term experiments15. Previous studies of the two Kenyan studied soils have shown an increase in P microbial biomass, resin-P and Po-OH Hedley fractions of the high P fixing soil as a result of addition of organic P (FYM) combined with KH2PO416. This study, evaluated how further addition of manure and inorganic fertilizer may affect P turnover and availability and microbial communities with the hypothesis that: 1) the increased biomass P in the high P fixing will also be noticed by an increase in33P turnover using 33P under laboratory conditions and will affect P availability; 2) this increased biomass P in the high P fixing soil will be related to lower DNA concentrations relative to the low P fixing soil and changes in soil microbial communities and will be linked to change in soil microbial community structure.
MATERIALS AND METHODS
The high P fixing soil 4, was sampled in Malava, located in the upper midland zone at an altitude of 1595 m above mean sea level (Kakamega district, Kenya) approximately 450 km West of Nairobi. Its highest fixation capacity was defined based on its low SOM content and acidic soil e.g., 0.73% C, 0.06% N, pH = 5.3; pH = 5.3; 13.80 g kg–1 Fe and 23.85 g kg–1 Al16. Annual precipitations of about 950-1950 mm and annual mean temperature is about 18-21°C (Malava, Dispensary Meteorological Station).The soils are mainly developed on granites. Malava soils were never fertilized due to the relatively low socio-economic conditions of the population4.
The low P-fixing soil was from "Mau Summit". As for the previous site, the lowest fixation capacity was also defined based on its relatively high SOM content and acidic soil e.g., 2.47% C, 0.21% N, pH = 5.3, 55.23 g kg–1 Fe and 36.31 g kg–1 Al16. It was sampled in an upper highland zone at an elevation of 2337 m above mean sea level (Nakuru district, Kenya) about 250 km West of Nairobi. The mean annual precipitation is less of about 1100 mm (1997). The temperature ranges from 18 -23°C (maximum) and 7-10°C (minimum) (Mau Summit, Meteorological station). The soils of the area have developed on ashes and other pyroclastic rocks from recently active volcanoes17. Occasional application of moderate amounts of Diammonium Phosphate (DAP) was applied at about 50 kg DAP per acre4.
Manure and soil sampling and analyses: The cow manure and soil samples used in this study were obtained from the same fields in Malava and Mau Summit where Ayaga et al.4 conducted their experiments. Details on manure and soil sampling are given in the publication by Koutika et al.16. The microbial biomass P turnover was determined according to Kouno et al.10. Soils were adjusted to 40% of Water Holding Capacity (WHC) and incubated for 7 days at 25°C in the dark. The 4 treatments were: 1) control, 2) + FYM (10g kg–1 soil), 3) KH2PO4 (g kg kg–1 soil) and 33P and 4) FYM (10g kg–1 soil) + KH2PO4 (g kg–1 soil) and 33P. The \pard fs18 33P solution was prepared by adding 20 MBq33P to 14.5 mL of 31P solution (10 mgP L–1). The 33P solution (3 mL of the solution equivalent to 4.138 MBq) was sprayed uniformly on 200 g of soil surface of each treatment. Therefore, there was 20.69 KBq33PO4 g–1 soil and 0.15 μg 31PO4g–1soil in the third and fourth treatments. 3 mL of water was added to the first and second treatments to obtain the same amount of WHC. The soils were then covered for 24 h to allow the solutions to equilibrate and were gently thoroughly mixed. Thereafter, the soils were incubated for a 7 days. After the first seven day incubation, FYM, containing 22.5% C and 1.42% N and milled to <1mm, was added to treatments 2 and 4 (a rate equivalent to 5000 μg C g–1 soil). Then all soils were incubated for another 7 days and the first 33P extraction and measurement were made (Day 0). The extraction was made at Day 0, 21, 42 and 63 after the initial 14 day equilibration incubations were completed. Biomass P was determined following using the same approach3. Molybdate reactive P was measured according to Murphy and Riley18 with a Lambda 25 UV/VIS spectrometer. The 33P in the soil extracts was measured by liquid scintillation counting using separate aliquots of soil extract (3 mL) mixed with ultima Gold scintillation cocktail (15 mL) and counted for 10 min on a Perkin Elmer TRI-CARB 2500 scintillation counter. The 33P measurements were corrected for decay back to the date of addition of 33P to soil. The 33P concentrations in biomass P were calculated using relative Specific Activity rel. SA = (r/R)/Qp; where r is the 33P measured in biomass P (kBq kg–1soil), R is the total 33P applied (kBq kg–1soil) and Qp is the total P concentration (mg P kg –1soil) according to Fardeau method19.
The Multi-TRFLP analysis was made on the high and low P fixing soils moistened at 40% and incubated for 16 weeks at 25°C in 3 replicates per treatment following the method described by Singh et al.11. Based on the results of SMB-P and P fractionation16, only (i) Nil or control and (ii) FYM (10 g per kg) + KH2PO4 (2.5 g kg–1) were considered in this study and the Multi-TRFLP was only made at week 1 and 16. (1) DNA extraction was made from 0.25 g using an ultra clean soil DNA isolation kit according to the manufactures’ instructions (Mo Bio laboratories, Carlsbad, CA). (ii) PCR optimization: DNA samples were amplified with PCR primers specific for bacteria, fungi and archaea individually. For each DNA sample, a multiplex PCR was performed with the reaction mixture containing all the three primer pairs for bacteria, fungi and archaea. The PCR master mixture contained 2 μL of NH4 reaction buffer, 0.6 μL MgCl2 and 0.4 μL m MdNTPs, 2 μL Muyzer forward primer (100 PM μL–1) 16s, 2 μL Muyzer reverse primer 16s, 0.1 μL Tag 9.9 μL H2O and 3 μL of template DNA. The samples were performed using the program consisted of an initial step of 5 min at 95°C followed by cycles of denaturation at 95°C for 30s, annealing at 55°C for 30s and elongation at 72°C for 1 min. The last cycle was followed by extension at 72°C for 10 min. The PCR amplicons were made with ethidium bromide and visualized on a 1.5% agarose gel using UV radiation. TRFLP analysis PCR products were purified a GenElute PCR clean-up kit (sigma-Aldrich, Dorset UK) according to manufacturer’s instructions. Prior to digestion, purified PCR product concentrations were determined with a spectrophotometer (UV photometer, Eppendorf, Germany).
All multiplex PCR were digested with the restriction enzymes in 20 μL reaction mixtures containing 500. 10–9 (ng) of PCR products, 1×buffer, 0.1 μg μL–1 of acetylated bovine serum albumin and 20 U of restriction enzyme. Samples were incubated for 3 h at 37°C and then deactivated at 95°C for 15 min. After digestion, 2 μL of each sample was mixed with 0.3 μL of LIZ-labelle GS500 (-250) internal standard of 12 μL of formamide. Prior to fragment analysis, samples were denatured at 95°C for 5 min and then chilled on ice for 5 min (Norwich, UK).
Statistical analysis: Mean and standard error of the mean were calculated. One-way analysis of variance (ANOVA) followed by the Tukey’s HSD was used to estimate the effects of the combined manure and inorganic fertilizers on P availability and microbial communities. Differences were taken as significant when p<0.05. Pearson correlation coefficients (r) between these measured variables were calculated and taken as significant when p<0.05. All statistical analyses were made with the R software20.
RESULTS
The 33P specific activity was lower than 0.10 μg P g soil–1 at Day 0 in the KH2PO4 and FYM+ KH2PO4 treatments of both soils (Fig. 1a). Then the specific activity of the high P fixing soil significantly p<0.05 increased at Day 21 (Fig. 1c), while no significant increase was measured in the low P fixing soil (Fig. 1b, d).
At Days 42 and 63, the specific activities in the two treatments were lower than 0.10 μg P g soil–1. However, the specific activity of the high P fixing soil was statistically higher than that of the low P fixing soil at Day 42 (Fig. 1g, h), except for the FYM at Day 42. The effects of soil type on 33P specific activity were found at Day 21, 42 and 63, while significant and beneficial effects of FYM addition were only found at Day 42 in the high P fixing soil (Malava, Fig. 1e).
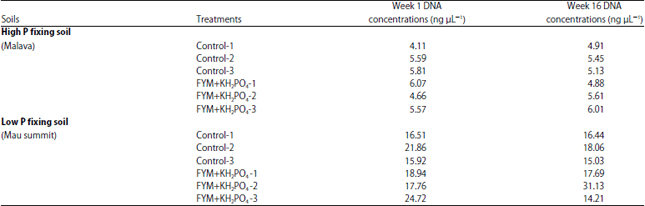
The DNA concentrations of the low P fixing soil were much higher (>14 ng μL–1) than those of the high P fixing soils (<10 ng μL–1) (Table 1). However, the Multi TRFLP samples of the studied two soil at week 1 and 16 and showed difference in the microbial communities of two soils (Fig. 2). This difference in microbial communities are pronounced in the FYM+KH2PO4 treatment of the high P fixing soil at week 1 and 16 than in the low P fixing soil counterpart (Fig. 2).
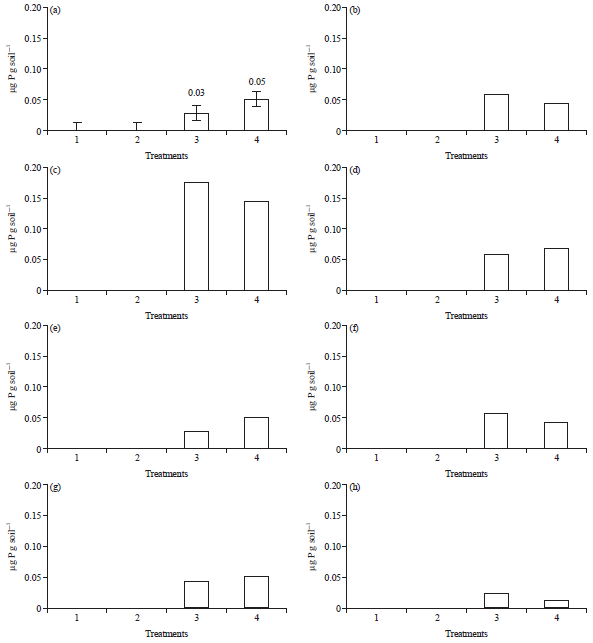 | |
| Fig. 1(a-h): | 33P Specific Activity in the high (Malava i.e., a), c), e) and g) and low (Mau Summit i.e., b), d), f) and h)) P fixing soils incubated at 25°C at Day 0, 21, 42 and 63 (mean values withstandard errors) |
| Table 1: | DNA concentrations in the high and low P fixing soils moistened at 40% of water holding capacity (WHC) at week 1 and 16 |
 | |
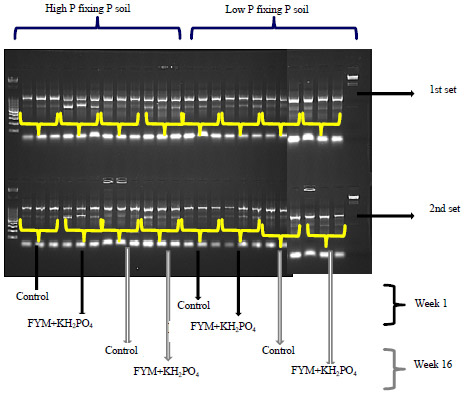 | |
| Fig. 2: | Multi T-RLFP procedure in the high and low P fixing soils moistened at 40% of water holding capacity at week 1 and 16 of incubation at 25°C (Control and FYM+KH2PO4 ) |
DISCUSSION
33P specific turnover: The 33P specific activity of the high P fixing soil was much higher than that of the low P fixing soil and confirmed the first hypothesis of this study i.e., the increased biomass P in the high P fixing observed in previous study16 induced an increase in 33P turnover of high P fixing soil. Even though in the P limiting condition, P microbial immobilization may be enhanced7, this result is in agreement with the findings by Buchler et al.8, revealing that inorganic P is much exchangeable and P dynamics is enhanced in the soil with the low inorganic P reserves and low C content e.g., the high P fixing soil. In the planted mixed species forest established on poor Congolese Arenosols (C% <1.2, N% <0.06 and P< 10 mg kg–1 in the 0.05 m), P dynamics were enhanced due to both high plant residues and higher demand for soil P to maintain the stoichiometry of leaves of N fixing trees at the end of the 7-year first rotation21, especially of those in the pure nitrogen fixing species stands at year 2 of the second rotation22.
The current increase in33P turnover and P availability of the high P fixing confirms our first hypothesis. In the studied high fixing P soil with a low inorganic reserve, an enhanced P dynamics has been already found through an increase in P biomass and P availability e.g., resin P and Po-OH Hedley fractions relative to the low P fixing soil4,16. The difference in P dynamics of the two soils may be due to difference in soil properties i.e., 0.73% C, 0.06% N, pH = 5.3, 13.80 g kg–1 Fe and 23.85 g kg–1 Al for the high fixing soils vs 2.47% C, 0.21%N, 55.23 g kg–1 Fe and 36.31 g kg–1 Al for the low P fixing soil16. Achat et al.23 highlighted the dominant role of organic carbon in controlling the dynamics of phosphate ions in acidic and non-acidic soils besides aluminium and iron oxides. This high 33P specific activity found in the Kenyan high P fixing soil seems to indicate an enhanced P turnover probably due to low organic content and high P fixing ability. Even conducted in laboratory conditions and without plant growth, the increased biomass P of high fixing soil16 might prevent P from fixation and increases P availability as shown by its high 33Pspecific activity mainly at Day 42 (Fig. 1e). This is in line with Koutika et al.16, who argued that in the low input systems, the biomass P turnover is a large potential as source of P for plants. Therefore, this study is confirming previous studies which have shown the importance of combining organic residues with inorganic fertilizer to both increase soil fertility through an increase in available P and improve crop growth and yield4,14. This result is relevant for weathered soils in many countries where P is fixed and occluded in the oxides iron and aluminium.
DNA concentrations: Even though, the DNA concentrations were higher in the low P fixing soil than in the high fixing soil, a probable change in microbial groups seems to be more pronounced in the high P fixing soil e.g., the FYM+KH2PO4 treatment at week 1 and 16 (Fig. 2a). This result confirms the second hypothesis and shows that the increased P biomass in the high P fixing soil may have enhanced both P turnover and change in DNA concentrations due to change in the microbial community structure, following addition of C components from manure (Fig. 2). In a long-term experiment, Marschner et al.13 argued that the addition of organic amendments is of fundamental importance for soil microorganisms, while the release of carbon compounds commonly increases microbial biomass and activity in the rhizosphere with probable impact on community structure and function24. Similarly, Brant et al.25 demonstrated that microbial utilization of C compounds to change as result of altering plant inputs to soil. They also argued that many changes in microbial community are due to alteration of their size and composition. The low DNA concentrations in the high P fixing soil is probably due to its low nutrients reserve compared to the low P fixing soil, but may also be due to lower sensitivity to C components derived from manure.
CONCLUSION
Even though conducted under laboratory conditions within short time frame (63 days), the results of this study reveal an enhanced P turnover with low DNA concentrations in the high P fixing soil relative to the low P fixing soil after the addition of farmyard manure and superphosphate. The enhanced P turnover and low DNA concentrations of the high P fixing soil were probably due to its poor-nutrient status (0.73% C, 0.06 % N) compared to the low P fixing soil, much richer (2.47% C, 0.21% N) and a lower sensitivity to added C components through manure. Both the applied methods have confirmed the hypotheses of the study: the high 33P specific activity is more pronounced in the high P fixing soil, poor in P reserve resources, however more able to enhance the P turnover, even though a low DNA concentrations was noticed. Therefore, this practice is be useful in cropping systems in weathered soils, where P is occluded by iron and Fe oxides, to reduce it fixation and increase crop growth and yields.
SIGNIFICANCE STATEMENTS
Through the use of labelled phosphorus 33P, this study highlighted an enhanced phosphorus turnover in the highly phosphorus-fixing soils in Kenya relative to the low fixing soils, after the combined addition of manure and inorganic fertilizer. The contrary tendency was found for the DNA concentrations. These findings might help researchers and farmers to propose and apply agricultural practices that enhance the turnover and availability of phosphorus which is usually mostly occluded by Fe and Al in weathered soils of sub Saharan Africa. This will help to sustain crop production in these areas, where phosphorus is one of the most limiting nutrients due to its low natural availability and the high costs of fertilizers.
ACKNOWLEDGMENTS
The authors thank to Dr. J. Andralojc, Ian Clark and R. White (Rothamsted Research) for their assistance in the setting of experiments and for the statistical analysis. LS. Koutika is very grateful to Rothamsted International which allowed this research.
REFERENCES
- Brookes, P.C., D.S. Powlson and D.S. Jenkinson, 1984. Phosphorus in the soil microbial biomass. Soil Biol. Biochem., 16: 169-175.
CrossRefDirect Link - Brookes, P.C., D.S. Powlson and D.S. Jenkinson, 1982. Measurement of microbial biomass phosphorus in soil. Soil Biol. Biochem., 14: 319-329.
CrossRefDirect Link - Ayaga, G., A. Todd and P.C. Brookes, 2006. Enhanced biological cycling of phosphorus increases its availability to crops in low-input sub-Saharan farming systems. Soil Biol. Biochem., 38: 81-90.
CrossRefDirect Link - Heuck, C., A. Weig and M. Spohn, 2015. Soil microbial biomass C: N: P stoichiometry and microbial use of organic phosphorus. Soil Biol. Biochem., 85: 119-129.
CrossRefDirect Link - Liu, A., C. Hamel, T. Spedding, T.Q. Zhang, R. Mongeau, G.R. Lamarre and G. Tremblay, 2008. Soil microbial carbon and phosphorus as influenced by phosphorus fertilization and tillage in a maize-soybean rotation in South-Western Quebec. Can. J. Soil Sci., 88: 21-30.
Direct Link - Bunemann, E.K., A. Oberson, F. Liebisch, F. Keller, K.E. Annaheim, O. Huguenin-Elie and E. Frossard, 2012. Rapid microbial phosphorus immobilization dominates gross phosphorus fluxes in a grassland soil with low inorganic phosphorus availability. Soil Biol. Biochem., 51: 84-95.
CrossRefDirect Link - Buehler, S., A. Oberson, I.M. Rao, D.K. Friesen and E. Frossard, 2002. Sequential phosphorus extraction of a 33P-labeled oxisol under contrasting agricultural systems. Soil Sci. Soc. Am. J., 66: 868-877.
CrossRefDirect Link - Oberson, A., D.K. Friesen, I.M. Rao, S. Buhler and E. Frossard, 2001. Phosphorus transformations in an oxisol under contrasting land-use systems: The role of the soil microbial biomass. Plant Soil, 237: 197-210.
CrossRefDirect Link - Kouno, K., J. Wu and P.C. Brookes, 2002. Turnover of biomass C and P in soil following incorporation of glucose or ryegrass. Soil Biol. Biochem., 34: 617-622.
CrossRefDirect Link - Singh, B.K., L. Nazaries, S. Munro, I.C. Anderson and C.D. Campbell, 2006. Use of multiplex terminal restriction fragment length polymorphism for rapid and simultaneous analysis of different components of the soil microbial community. Applied Environ. Microbiol., 72: 7278-7285.
CrossRefDirect Link - Osborn, A.M., E.R.B. Moore and K.N. Timmis, 2000. An evaluation of Terminal-Restriction Fragment Length Polymorphism (T-RFLP) analysis for the study of microbial community structure and dynamics. Environ. Microbiol., 2: 39-50.
CrossRefDirect Link - Marschner, P., E. Kandeler and B. Marschner, 2003. Structure and function of the soil microbial community in a long-term fertilizer experiment. Soil Biol. Biochem., 35: 453-461.
CrossRefDirect Link - Chu, H., X. Lin, T. Fujii, S. Morimoto, K. Yagi, J. Hu and J. Zhang, 2007. Soil microbial biomass, dehydrogenase activity, bacterial community structure in response to long-term fertilizer management. Soil Biol. Biochem., 39: 2971-2976.
CrossRefDirect Link - Marschner, P., C.H. Yang, R. Lieberei and D.E. Crowley, 2001. Soil and plant specific effects on bacterial community composition in the rhizosphere. Soil Biol. Biochem., 33: 1437-1445.
CrossRefDirect Link - Koutika, L.S., T.E. Crews, G. Ayaga and P.C. Brookes, 2013. Microbial biomass P dynamics and sequential P fractionation in high and low P fixing Kenyan soils. Eur. J. Soil Biol., 59: 54-59.
CrossRefDirect Link - Murphy, J. and J.P. Riley, 1962. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta, 27: 31-36.
CrossRefDirect Link - Fardeau, J.C., 1993. Le phosphore assimilable des sols: Sa representation par un modele fonctionnel a plusieurs compartiments. Agronomie, 13: 317-331.
Direct Link - Koutika, L.S., D. Epron, J.P. Bouillet and L. Mareschal, 2014. Changes in N and C concentrations, soil acidity and P availability in tropical mixed acacia and eucalypt plantations on a nutrient-poor sandy soil. Plant Soil, 379: 205-216.
CrossRefDirect Link - Koutika, L.S., L. Mareschal and D. Epron, 2016. Soil P availability under eucalypt and acacia on Ferralic Arenosols, republic of the Congo. Geoderma Regional, 7: 153-158.
CrossRefDirect Link - Achat, D.L., N. Pousse, M. Nicolas, F. Bredoire and L. Augusto, 2016. Soil properties controlling inorganic phosphorus availability: General results from a national forest network and a global compilation of the literature. Biogeochemistry, 127: 255-272.
Direct Link - Morgan, J.A.W., G.D. Bending and P.J. White, 2005. Biological costs and benefits to plant-microbe interactions in the rhizosphere. J. Exp. Bot., 56: 1729-1739.
CrossRefDirect Link - Brant, J.B., E.W. Sulzman and D.D. Myrold, 2006. Microbial community utilization of added carbon substrates in response to long-term carbon input manipulation. Soil Biol. Biochem., 38: 2219-2232.
CrossRefDirect Link