
R.W. Mukhongo
Tropical Soil Biology and Fertility Institute of the International Centre for Tropical Agriculture (TSBF-CIAT), ICIPE Duduville Campus, Nairobi, Kenya
Department of Soil Science, University of Eldoret, Eldoret, Kenya
M.A. Kavoo-Mwangi
Tropical Soil Biology and Fertility Institute of the International Centre for Tropical Agriculture (TSBF-CIAT), ICIPE Duduville Campus, Nairobi, Kenya
Department of Horticulture, Jomo Kenyatta University of Agriculture and Technology, Nairobi, Kenya
School of Agriculture and Biotechnology, Karatina University, Karatina, Kenya
M.E. Kahangi
Department of Horticulture, Jomo Kenyatta University of Agriculture and Technology, Nairobi, Kenya
E.M. Ateka
Department of Horticulture, Jomo Kenyatta University of Agriculture and Technology, Nairobi, Kenya
A.B. Were
Department of Soil Science, University of Eldoret, Eldoret, Kenya
J.R. Okalebo
Department of Soil Science, University of Eldoret, Eldoret, Kenya
M.E. Mutegi
Tropical Soil Biology and Fertility Institute of the International Centre for Tropical Agriculture (TSBF-CIAT), ICIPE Duduville Campus, Nairobi, Kenya
K.E. Mwangi
Tropical Soil Biology and Fertility Institute of the International Centre for Tropical Agriculture (TSBF-CIAT), ICIPE Duduville Campus, Nairobi, Kenya
T.T. Tepeni
University of Nairobi, Nairobi, Kenya
K.S. Njuguini
Department of Mycology, National Museums of Kenya, Nairobi, Kenya
J.M. Onguso
Department of Horticulture, Jomo Kenyatta University of Agriculture and Technology, Nairobi, Kenya
S.A. Okoth
University of Nairobi, Nairobi, Kenya
J.M. Jefwa
Tropical Soil Biology and Fertility Institute of the International Centre for Tropical Agriculture (TSBF-CIAT), ICIPE Duduville Campus, Nairobi, Kenya
International Journal of Soil Science
Year: 2015 | Volume: 10 | Issue: 2 | Page No.: 45-62







ABSTRACT
The impact of microbiological commercial products (PHC Biopak, Rhizatech and ECO-T) on the occurrence of mycorrhizae and Fusarium in the rhizosphere of tissue culture banana (Gros Mitchel cv.) was assessed. Tissue cultured banana plantlets were inoculated with PHC Biopak (Bacillus), Rhizatech (mycorrhiza) and ECO-T (T. harzianum) under greenhouse conditions using a completely randomized design in a Vertisol, Rhodic Ferralsol and Humic Nitisol sampled from the major banana growing regions in Kenya. Potted plants were later established under field conditions in the three agro ecological zones. Roots and soils sampled at end of potting and at flowering were assessed for AM fungi colonization and Fusarium populations. The effect of product inoculation on AM fungi colonization varied and only significant (p<0.05) in Rhodic Ferralsol with Rhizatech increasing intensity of colonization by 31.9% and PHC Biopak increasing the frequency of colonization by 38.6% compared to the non-inoculated control (12.9%). F. oxysporum, fsp. cubense, F. proliferatum and F. incarnatum were recovered from the experimental soils. Foc was the most abundant in the three soils (prior to inoculation) accounting for 60.6% of all Fusarium colony forming units. After inoculation, at the end of potting stage and at flowering, F. proliferatum was mostly isolated from the three zones accounting for 35.2% of the total fungal population. Foc was isolated from Humic Nitisol and Vertisol accounting for 11.5% of the total fungal population. PHC Biopak, ECO-T and Rhizatech suppressed Foc colony forming units per gram of soil by 47, 68 and 55%, respectively in the Humic Nitisol. ECO-T reduced Fusarium colony forming units per gram of soil by 6% in Rhodic Ferralsol and PHC Biopak by 50% in Vertisol compared to the non-inoculated soils. There is potential in use of commercial microbiological products to suppress Foc and the efficacy of the products depends on soil physico-chemical properties.
PDF Abstract XML References Citation
Received: March 08, 2015;
Accepted: May 26, 2015;
Published: June 26, 2015
How to cite this article
R.W. Mukhongo, M.A. Kavoo-Mwangi, M.E. Kahangi, E.M. Ateka, A.B. Were, J.R. Okalebo, M.E. Mutegi, K.E. Mwangi, T.T. Tepeni, K.S. Njuguini, J.M. Onguso, S.A. Okoth and J.M. Jefwa, 2015. Occurrence of Arbuscular Mycorrhizal Fungi and Fusarium in TC Banana Rhizosphere Inoculated with Microbiological Products in Different Soils in Kenya. International Journal of Soil Science, 10: 45-62.
DOI: 10.3923/ijss.2015.45.62
URL: https://scialert.net/abstract/?doi=ijss.2015.45.62
DOI: 10.3923/ijss.2015.45.62
URL: https://scialert.net/abstract/?doi=ijss.2015.45.62
INTRODUCTION
Microorganisms living in the rhizosphere can have a neutral, pathogenic or beneficial interaction with their host plant (Whipps, 2001; Raaijmakers et al., 2009). Banana (Musa acuminate Colla AAA), is a monocotyledonous herbaceous species, that shows a great ability to establish mycorrhizal symbiosis (Jaizme-Vega et al., 2002). Mycorrhiza are regarded as tripartite symbioses since, they commonly interact with bacteria and other soil organisms producing beneficial effects on plant nutrition and health as well as on soil structure and stability (Frey-Klett et al., 2007). Although little detailed information is available on the direct impact and interaction of bacteria on mycorrhizal fungi, it has been shown that the germination of mycorrhizal spores can be affected by the presence of some bacteria (Xavier and Germida, 2003). Some of the bacteria associated with Arbuscular Mycorrhiza (AM) fungi, can improve the mycorrhizal colonization (Hildebrandt et al., 2002), improve root branching (Gamalero et al., 2002), or present antifungal properties (Budi et al., 1999). Since, they share common microhabitats, AM fungi and Plant Growth Promoting Bacteria (PGPBs) must interact during the colonization process and/or as rhizosphere microorganisms (Gamalero et al., 2004).
It has been proposed that plants must be mycorrhizal to thrive in degraded nutrient-poor and arid soils (Barea, 2000) and that mycorrhizal effect can be improved by co-inoculation with mycorrhiza-helper bacteria, which can play an important role in stressed areas (Requena et al., 1997; Vazquez et al., 2000). In vitro experiments in which saprotrophic fungi were paired with spores of Glomus mosseae or Gigaspora rosea showed a direct effect of Trichoderma pseudokoningii on the germination of spores of both AM fungi. This suggests a direct interaction between the mycorrhizal fungus and the saprotrophic fungi in the pre-symbiotic phase of the former. Though many studies have been conducted on banana, there are not many references concerning the effect of commercially prepared microbiological inoculants on the occurrence of AM fungi and the pathogenic Fusarium oxysporum (Perez and Jaizme-Vega, 1997). Furthermore, most studies on artificial microbiological inoculation of banana have been done under greenhouse conditions but have rarely considered the persistence of the inoculants under field conditions or their effect on other beneficial or pathogenic conditions (Kavoo-Mwangi et al., 2014).
The use of tissue cultured plants as planting materials leads to a reduction in the spread of Fusarium oxysporum, f.sp. cubense (Foc), but at the same time, results in enhanced susceptibility to Foc under field conditions due to the loss of native endophytes during tissue culturing, including beneficial plant growth promoting rhizobacteria and fungi (Vuylsteke, 2000). Therefore, biotization of tissue culture plantlets with native effective non-pathogenic endophytic microbes including mycorrhizal fungi at the acclimatization stage enhances plant resistance to tissue cultured plants against Fusarium wilt (Nowak, 1998). Several reports have previously demonstrated the successful use of different species of Trichoderma, Pseudomonas, Streptomyces, mycorrhiza and non-pathogenic Fusarium of both rhizospheric and endophytic in nature against Fusariumwilt disease under both greenhouse and field conditions (Thangavelu et al., 2002; Thangavelu and Mustaffa, 2012). The potential of Bacillus, Trichoderma and mycorrhiza (AM fungi) in suppressing Fusarium oxysporum f.sp. cubense (Foc) could be harnessed by biological acclimatization. These microorganisms proliferate in the rhizosphere and may mitigate the challenge posed by soil borne Foc. The use of non-sterile field soils for establishment of TC banana in the nursery has not been explored in Kenya despite the opportunity it offers for decentralization of commercial TC banana nurseries to increasingly meet the demand for plantlets. This study aimed at evaluating the potential of mycorrhizal, Bacillus and Trichoderma based commercial products on growth of AM fungi and suppression of Fusarium spp. populations in the rhizosphere of tissue cultured banana grown in greenhouse and field soils.
MATERIALS AND METHODS
Source of soil and tissue culture plants: Tissue cultured banana plantlets cv. Gros Michel with 3 fully developed leaves were obtained in nutrient agar (Murashige and Skoog, 1962) from Jomo Kenyatta University of Agriculture and Technology (JKUAT) Biotechnology laboratory. Three soil types: Vertisol, Rhodic Ferralsol and Humic Nitisol were sampled from three agro ecological zones in Kenya where bananas are grown i.e., Western Kenya in Nyanza (Vertisol), coastal Kenya in Kilifi (Rhodic Ferralsol) and central Kenya in Meru south (Humic Nitisol) at a depth of 0-20 cm and used for hardening and potting of tissue culture plantlets. The 0-20 cm soil depth was chosen for mycological considerations since it contains the majority of soil microbiota (Skujins, 1984).
Experimental design and inoculation process of tissue culture banana plants: A three by three factorial experiment consisting of three soil types (Vertisol, Humic Nitisol and Rhodic Ferralsol) and three commercial microbe-based products including ECO-T (Trichoderma), PHC Biopak (Bacillus) and Rhizatech (mycorrhizal) were used. The source of products, composition, rate and mode of application is described in Table 1. The products are normally recommended for plant growth but due to the composition of the products, this attempt was made to evaluate whether they also have capacity to reduce soil pathogens. Each treatment was replicated three times and the experimental units subjected to a completely randomized design in a greenhouse.
The inoculation of plantlets with commercial products was initially done at the deflasking stage during the hardening process and subsequently at the beginning of the potting stage, eight weeks after deflasking. Soils used for hardening of tissue culture plants were sterilized before being used for hardening by autoclaving twice for 30 min at 80°C following the mycorrhizal training manual prepared from the Centre for Ecology and Hydrology, Peniciuk, UK (Ingleby, 2007). Non-sterile soils were used for potting of TC plants eight weeks after deflasking.
Initial soil physico-chemical characterization: Soil chemical and physical characterization was done before inoculation with commercial products for nutrient composition (nitrogen, phosphorous, potassium, carbon, magnesium, calcium and sodium), Cation Exchange Capacity (CEC), pH and soil texture composition (clay (%), sand (%) and silt (%)) (Table 2). This was done according to procedures of Anderson and Ingram (1998) and Okalebo et al. (2002).
| Table 1: | Description of microorganism based commercial products used in experiment |
 | |
| Table 2: | Initial soil characteristics (0-20 cm) of vertisol, rhodic ferralsol and humic nitisol |
 | |
Kavoo-Mwangi et al. (2013). Olsen P, K, Ca, Mg, Na, ECEC measured in cmol kg–1, N, C, sand, clay and silt measured in percentage | |
Soil mycological characterization
Morphological characterization of isolated cultures: Soil samples were thoroughly mixed aseptically and air dried over night before Fusarium isolation. Fusarium spp were isolated from initial soil samples using serial dilution plating method (Burrges et al., 1988) with 0.1% Tap Water Agar (TWA) (Brayford, 1993). Air dried soil samples (10 g) were suspended into dilution blanks (90 mL of sterile TWA) to make a ten-fold dilution (10–1) of a microbial suspension. The dilution was shaken to obtain a uniform suspension of microorganisms and 10 mL of resulting suspension were pipetted into a flask containing 90 mL of sterile distilled water. This procedure was repeated up to the third ten-fold dilution. One millilitre aliquots from second and third dilutions, in three replicates, were aseptically pipetted on to petri dishes containing Fusarium-selective PCNB-Peptone Agar (PPA) media and spread evenly across the agar surface using a sterile glass applicator. The petri dishes were kept at room temperature (25°C) and observations were made from the third day onwards for developing colonies. For each of the colonies growing on PPA plates, a well-defined and shaped colony was chosen and a small piece at the edge of the colony was carefully and aseptically transferred onto a separate Synthetic Nutrient Agar (SNA) petri plate and incubated at 25°C for 5 days. Subsequently, in order to obtain monosporic cultures of each colony formed on SNA, from which identification was based, very dilute inocula, of 5-10 spores per drop of suspension (when viewed at low power magnification), were prepared and spread on 2% Tap Water Agar plates. These were then incubated for 15 h for germination. Germlings were then sub-cultured on different media i.e., SNA, Carnation-Leaf-Agar (CLA) and Potato-Dextrose-Agar (PDA) media plates, for growth and identification. Species of Fusarium readily formed sporodochia with robust, uniform macroconidia on the CLA that was used for identification. The PDA cultures were used to assess pigmentation and gross colony morphology. Cultures grown on SNA were evaluated for microconidia, which were more abundant and diverse on this medium and for chlamydospores, which were more common and produced rapidly on this medium. All the pure isolates sub-cultured on PDA, CLA and SNA were incubated for ten to twenty days at 25°C under fluorescent lamps to enhance sporulation. Fusarium was identified to the species level where morphological characters were used as the basis of identification (Nelson et al., 1983; Burrges et al., 1988; Brayford, 1993). After identification, the single spore cultures were stored in agar slants of SNA in screw cap bottles at 4°C and in sterilized soil in screw cap bottles.
Molecular characterization of morphologically identified cultures: In order to obtain DNA from each of the identified Fusarium species, single-spore Fusarium species were grown for five days at 25°C in Potato Dextrose Agar (PDA) (Difco). Mycelium (~0.1-0.2 g) was collected using sterile scalpel from PDA media and placed in eppendorf tubes. Culture cells were opened by adding 500 μL of CTAB extraction buffer (100 mM Tris HCl (pH 8), 2% (wt/v) CTAB, 50 mM EDTA, 0.7 M NaCl, 0.17% (v/v) β-mecarptoethanol and 1% (w/v) PVP), pre-warmed to 65°C, two glass beads added and the mixture placed in miller at a frequency of 30 sec for 5 min. Samples were incubated at 65°C for 30 min in a water bath. Chloroform (500 μL)-isoamylcholoroform) (24:1 v/v) was added and the two phases were mixed several times by inverting tubes gently. The tubes were centrifuged at 14,000 rpm for 10 min at room temperature in a microfuge. The supernatant was removed and transferred into new 1.5 eppendorf tube. Pre-boiled 10 μL of 10 mg RNase A was added and mixed gently by inverting the tube five times. The samples were then incubated in a water bath at 37°C for 30 min. Centrifuging and addition of RNase A were repeated. An equal volume of cold isopropanol (pre-chilled in a -20°C freezer) was added mixed gently and incubated at -20°C in a freezer for 30 min. The samples were then centrifuged at 14000 rpm for 10 min at room temperature in a microfuge and supernatant removed by pouring into clean eppendorf tubes taking care not to loose the pellet. Five hundred micro liter of 70% ethanol (at room temperature) was added to the tube containing DNA, centrifuged at 14000 rpm for 5 min and the supernatant carefully poured off. The 70% ethanol wash was repeated once, the supernatant carefully poured off and the DNA pellet dried for 60 min by leaving the open tube on its side on the bench. Low salt TE buffer (100 μL) was added to the dried pellet. The pellet was dissolved by incubating at 37°C in a water bath for 30 min. One micro liter of DNA was ran on Agarose gel to determine the concentration and stored at -20°C.
DNA amplification and sequencing procedure: The Polymerase Chain Reaction (PCR) procedures were carried out as described by Khalil et al. (2003). Forward primer TEF 1 (5'-ATGGGTAAGGA(A/G)GACAAGAC-3') and the reverse primer TEF 2 (5'-GGA(G/A)GTACCAGT (G/C)ATCATGTT-3') (O’Donnell et al., 1998) were used to amplify the translation elongation factor 1-a (TEF) gene. The TEF region encodes an essential part of the protein translation machinery and has high phylogenetic utility because it is highly informative at the species level in Fusarium, non-orthologous copies of the gene have not been detected in the genus and universal primers have been designed that work across the phylogenetic breadth of the genus (Geiser et al., 2004). The amplification reactions were performed in 25 μL volumes in thin-walled PCR tubes after optimization in a PTC-100 (Programmable Thermal Controller), programmed for an initial cycle of 1 min at 95°C, 5 min at 95°C, annealing at 58°C and extension 1 min at 72°C, followed by 34 cycles of 5 min at 95°C, annealing at 58°C and extension 1 min at 72°C. There was a final extension step of 5 min at 72°C followed by a cooling to 4°C until samples were recovered. Amplified products were analyzed on a 1.5% agarose gel in 1X TAE buffer (40 mM Tris acetate and 1.0 mM EDTA) and documented using Bio-Rad documentation system.
The 700 bp PCR products were sequenced for the DNA region coding for the TEF gene using the BigDye terminator Cycle Sequencer (ABI, Foster City, CA). Seaview4 application (Gouy et al., 2010) was used for aligning sequences and building a phylogenetic tree derived from the partial sequences of the TEF gene by neighbor-joining method. Bootstrap values were set at 100 (100 replicates). The phylogenetic analysis was carried out to compare the degree of genetic relatedness of the TEF gene sequences of each isolate with those available in the GenBank database. Sequences obtained with each primer set were compared to GenBank nucleotide sequences by using nucleotide-nucleotide Basic Local Alignment Search Tool (BLAST) (www.ncbi.nlm.nih.gov/BLAST). Sequences were compared with closest matches in GenBank through BLAST.
Assessment of Fusarium populations at the end of nursery and potting phases: Soils were collected from the rhizosphere of the potted Tissue Cultured (TC) bananas, following a destructive harvest at the end of the nursery stage i.e., 22 weeks after deflasking of in vitro plants. The nursery phase lasted two and half weeks longer than the experimental nursery period followed by Rodriguez-Romero et al. (2005). Three soil replicates from each treatment were thoroughly mixed in sterile containers to constitute a composite sample from which 1 kg soil was taken and placed in sterile paper bags and labeled. Soils were stored at 4°C until they were processed. Isolation and identification of Fusarium spp. from the inoculated soils was carried out using the above mycological characterization procedures.
Assessment of Fusarium populations under field conditions: Soils samples were collected from the rhizosphere of the inoculated tissue cultured bananas in the field trials and assessed for Fusarium communities. Four sub-samples were thoroughly mixed in sterile containers to constitute a composite sample from which 1 kg soil was taken, placed in sterile paper bags and labeled. The auger was sterilized by dipping in 70% ethanol between sampling points to avoid cross contamination. The 0-20 cm soil depth was chosen for mycological considerations since it contains the majority of soil microbiota (Skujins, 1984). Samples were placed in a cool box during transportation and stored at 4°C in until they were processed. Isolation and identification of Fusarium spp. from the inoculated field soils was carried out using the above mycological characterization procedures.
Assessment of mycorrhizal colonization: Roots and soil samples collected from the rhizosphere of inoculated tissue cultured banana plants under green-house and field conditions were assessed for presence of AM fungi. Assessment was done by evaluating the percentage root colonization, spore abundance (field samples only). The roots were stained according to procedures of Koske and Gemma (1989). In to each bottle 2.5% potassium hydroxide (KOH) was added before heating in the oven at 70°C for 1 h. The KOH was poured off and the roots rinsed to remove KOH. Alkaline hydrogen peroxide was then added and roots left for 1 h to distain to remove the phenolic substances. Alkaline hydrogen peroxide was then poured off, the roots thoroughly rinsed with tap water and 1% hydrochloric acid (HCl) added and left for 1 h. After pouring off HCl 0.05% Trypan blue was added and the roots placed in the oven for 1 h. De-staining solution (500 mL glycerol, 450 mL of distilled water and 50 mL of 1% HCl) was added. Analysis of AM fungi colonization was done according to McGonigle et al. (1990). Slides were prepared with 30 pieces of roots each 1 cm long then examined under a compound microscope. The percentage of each piece covered by arbuscules, vesicles and hyphae was assessed to determine the intensity and frequency of AM fungi colonization.
Spores were extracted from soil using Jenkins (1964) procedure with modifications by Ingleby (2007). The soil (50 g) was pre-soaked in water and washed through 710 and 45 μm pore sieves with running water. The beaker content was decanted into 50 mL centrifuge tubes and centrifuged for 5 min at 1750 rpm. Water was carefully decanted from the tubes and floating debris discarded and 48% of sucrose (227 g dissolved in 500 mL of water) was added and centrifuging for 1 min at 1750 rpm. Immediately after centrifugation, sucrose solution was carefully decanted through a 45 μm sieve. The spores were rinsed thoroughly with water to wash out the sucrose. Spores were transferred from the 45 μm sieve into a small petri dish for examination under a dissecting microscope. Under a dissecting microscope, spores were counted and grouped according to their morphotypes. Microscope slides were prepared with very small drops of PVLG (polyvinyl lactoglycerin) and Melzer’s+PVLG reagents and characterized under a compound microscope based on appearance, morphology, sub-cellular features and reaction to Melzer’s.
Data analysis: Analysis of variance were performed on all measured variables using Proc MIXED in the SAS statistical software (SAS., 2006). The main effects of treatments, soils and their interactions were treated as fixed effects. Replicate and replicate×soil effects were considered random. Effects that were found to be significant at p≤0.05, their means were subsequently separated using the Studentized Tukey’s HSD test in GENSTAT. The data was subjected to Pearson’s correlation using SPSS analytical package. Percentage data for AM fungi colonization frequency and intensity was arcsine transformed while counts data was square root transformed before being subjected to analysis in order to stabilize variance in the data set and to bring about normal distribution. Results of analysis were interpreted without transforming.
RESULTS
Occurrence of Fusarium in soils used for greenhouse experiments: Three species of Fusarium, i.e., F. oxysporum, F. proliferatum and F. incarnatum were isolated from the experimental soils (before and after establishment of Tissue Culture (TC) banana under greenhouse conditions) as well as from the rhizosphere of TC banana plants under field conditions. The species were distinguished by pigmentation on PDA as forming white, white pink and white violet colonies, respectively (Fig. 1-3).
Conidia, colony morphology and pigmentation of Fusarium isolates from Vertisol, Rhodic Ferralsol and Humic Nitisol soils Vertisol, Rhodic Ferralsol and Humic Nitisol soils Vertisol and Humic Nitisol soils used for growing tissue cultured banana plantlets.
Molecular characterization by sequencing of the translation elongation factor 1-a (TEF) gene of the isolates using the NCBI database concurred with the morphological identification. The Polymerase Chain Reaction (PCR) product gel image is shown in Fig. 4.
DNA database comparison of the TEF-1α sequence showed 100% identity to sequences from the Foc (GenBank accession No. AF008486), F. proliferatum (GenBank accession No. FJ538244) and F. incarnatum (GenBank accession No. JF270209).
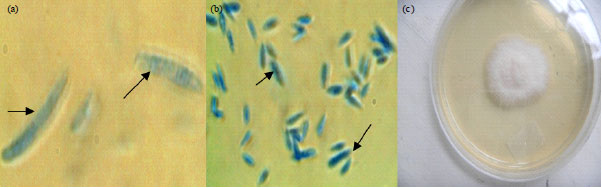 | |
| Fig. 1(a-c): | Fusarium proliferatum, (a) Macroconidia, (b) Microconidia and (c) Colony pigmentation on PDA, size of macroconidia and microconidia, 0.2 μm in width. X1000 magnification for macroconidia and X400 magnification for microconidia |
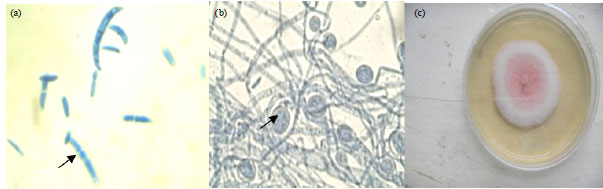 | |
| Fig. 2(a-c): | Fusarium oxysporum, (a) Microconidia and macroconidia, (b) Chlamydospores and (c) Colony pigmentation on PDA, size of macroconidia and microconidia, 0.4 μm in width. X400 magnification for both macroconidia and microconidia |
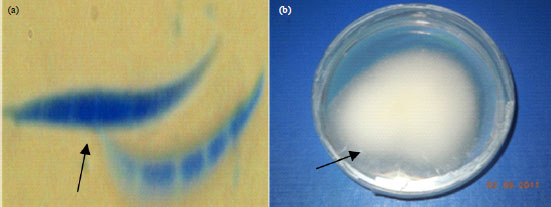 | |
| Fig. 3(a-b): | Fusarium incarnatum, (a) Macroconidia and (b) Colony pigmentation on PDA, size of spores, 0.2 μm. X1000 magnification for a and X400 magnification for b |
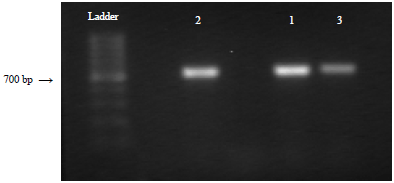 | |
| Fig. 4: | Gel image showing PCR product of Fusarium isolates, 1: Fusarium proliferatum, 2: Fusarium oxysporum and 3: Fusarium incarnatum |
The phylogenetic tree derived from the partial sequences of the translation elongation factor 1 and Foc, F. proliferatum and F. incarnatum standards obtained from the GenBank confirmed the genetic relatedness of the three isolates within the Fusarium genus as well as the correct identification of the species (Fig. 5).
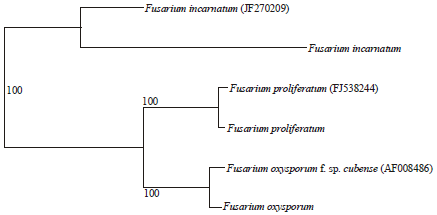 | |
| Fig. 5: | A phylogenetic tree of Fusarium isolates from soil derived from the partial sequences of the Translation Elongation Factor 1-α (TEF) gene by neighbor-joining method. Bootstrap values were set at 100 (100 replicates). Sequences with accession numbers are standard sequences from the NCBI GenBank |
DNA database comparison of the TEF-1α sequence showed 100% identity to sequences from the Foc (GenBank accession No. AF008486), F. proliferatum (GenBank accession No. FJ538244) and F. incarnatum (GenBank accession No. JF270209). The phylogenetic tree derived from the partial sequences of the translation elongation factor 1 and Foc, F. proliferatum and F. incarnatum standards obtained from the GenBank confirmed the genetic relatedness of the three isolates within the Fusarium genus as well as the correct identification of the species (Fig. 5).
Fusarium populations under greenhouse conditions: Preference for soil was observed in the occurrence of isolates with Foc prevalent in the central Kenya soil collected from Meru South (Humic Nitisol) and F. proliferatum prevalent in soils from western Kenya-Nyanza (Vertisol) and Coastal Kenya-Kilifi (Rhodic Ferralsol) and F. incarnatum occurring only in the Humic Nitisol and Vertisol. The morphological characteristics of the three isolates as observed in Potato Dextrose Agar (PDA) and Carnation Leaf Agar (CLA) are described in Plate 1. Foc was the most abundant in the three soils accounting for 60.6% of all Fusarium colony forming units (Fig. 6). Fusarium proliferatum was the second most isolated accounting for 32.9% of total number of isolates and mainly isolated in the Vertisol and Rhodic Ferralsol soils. Fusarium incarnatum was least isolated (6.4%) and was only found in the non-inoculated Humic Nitisol and Vertisol. Fusarium oxysporum was frequently isolated in all three soils especially in the non-inoculated Humic Nitisol (Fig. 6).
Effect of mycorrhiza, Trichoderma and Bacillus inoculation on Foc populations: We took special interest in the effect of microbiological inoculation on Foc since, it is the species associated with causing Fusarium wilt of banana. Statistically, the interaction between soil type and commercial products was highly significant at p<0.05 and Foc populations were influenced by soil type (Table 3). Non-inoculated soils from Humic Nitisol had the highest Foc (27×102 CFU g–1) compared to the non-inoculated Vertisol and Rhodic Ferralsol (2.67‾102 and 2.33×102 CFU g–1, respectively). Inoculation of Humic Nitisol soil with PHC Biopak, ECO-T and Rhizatech reduced Fusarium CFU g–1 by 47, 68 and 55%, respectively compared to the non-inoculated control soils.
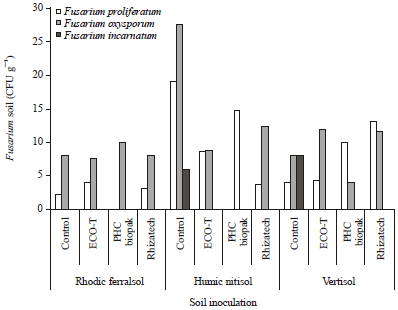 | |
| Fig. 6: | Population of Fusarium species in control and inoculated soils used for growing tissue cultured banana |
| Table 3: | Effect of Biopak, Rhizatech and ECOT on the occurrence of Fusarium and other fungi (CFU×102 g–1) in humic nitisol, ferralsol and vertisol |
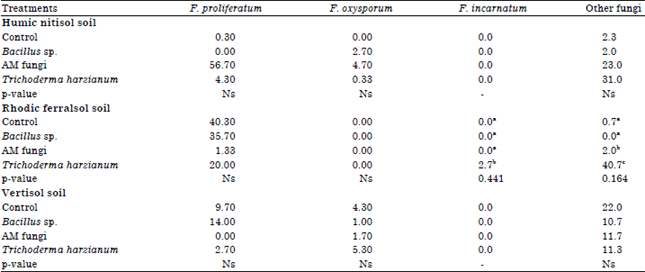 | |
Means within the same column with the same letter are not significantly different (Tukey test) at p≤0.05. Ns: not significantly different | |
Inoculation of the Vertisol with PHC Biopak reduced Foc population by 50% while inoculation with Rhizatech and ECO-T enhanced Foc populations by 50 and 44%, respectively compared to the non-inoculated soils. Inoculation of Rhodic Ferralsol with ECO-T reduced Foc population by 6% compared to the non-inoculated soils while PHC Biopak enhanced Foc populations by 25% while Rhizatech had no effect.
Other fungi were the most abundant in the three soils accounting for 50.8% of the fungal population. F. proliferatum accounted for 35.2% of the fungal population and was the most isolated Fusarium species from the three soils. F. oxysporum was isolated from Humic Nitisol and Vertisol accounting for 11.5% while F. incarnatum being the least isolated took only 1.4% of the fungal population.
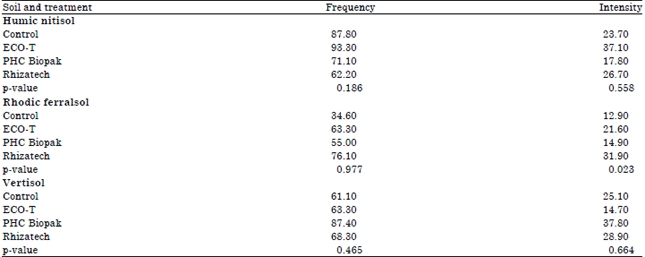
| Table 4: | Percent mycorrhizal colonization of tissue cultured banana plantlets inoculated with commercial microbiological products: 22 weeks after deflasking |
 | |
F. incarnatum was only isolated from the Rhodic Ferralsol. Other fungi are isolates that were not identified to be Fusarium. Most of these were of the genus Penicillium and they occurred in large numbers.
Effect of mycorrhiza, Trichoderma and Bacillus inoculation on mycorrhizal colonization under greenhouse conditions: Mycorrhizal colonization was only significantly affected (p<0.05) by product application in the Rhodic Ferralsol and this was only evident on the intensity of colonization (Table 4). Percent intensity of colonization was highest with Rhizatech inoculation (31.9%) and least with non-inoculated control (12.9%) in the Rhodic Ferralsol; highest with ECO-T inoculation (37.1%) in the Humic Nitisol; highest with PHC Biopak (37.8%) followed by Rhizatech (28.9%) and least with ECO-T inoculation (16.2%) in the Vertisol.
The effect of products on percent frequency of mycorrhizal colonization was variable and was highest with PHC Biopak (87.4) inoculation in the Vertisol. Rhizatech inoculation had the highest percent frequency of mycorrhizal colonization (76.1%) while, the control had the least (34.6%) in the Rhodic Ferralsol. Similar to the observations, made on percent intensity of colonization, inoculation of plants with ECO-T had the highest percent frequency of colonization (93.3%) in the Humic Nitisol.
Relationship between soil properties and Foc population: A strong positive correlation was observed between soil chemical and physical properties and Foc population (Table 5). Soil carbon (C) and nitrogen (N) had a positive effect on soil Foc r = 0.268 and 0.25, respectively. Soil physical properties had variable effects on Foc populations. Sandy soil was negatively correlated with Foc CFU g–1 showing a strong negative relationship with Foc CFU g–1 and a strong negative relationship between clay soil and Fusarium CFU g–1 (r = -0.25 and 0.25, respectively).
Effect of mycorrhiza, Trichoderma and Bacillus inoculation on mycorrhizal colonization and spore abundance in the field: The effect of the treatments on the intensity of root mycorrhizal colonization was not significant (p≤0.05) in the three soils (Table 6). Trichoderma harzianum increased the intensity of mycorrhizal colonization of banana roots by 4.1% when compared to the control in Humic Nitisol (Central soil).
| Table 5: | Pearson correlation coefficient (r) between Fusarium population and the soil chemical and physical properties in soils from central, coast and Western Kenya |
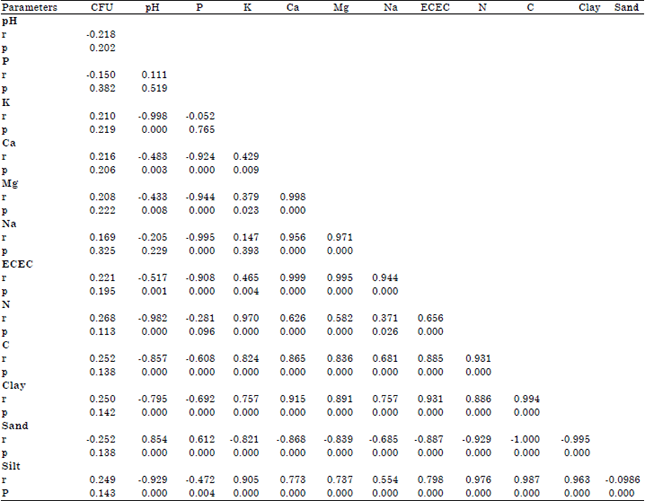 | |
| r, Pearson correlation coefficient; p, F probability at p ≤ 0.05 | |
| Table 6: | Effect of Bacillus, Trichoderma and AMF on mycorrhizal colonization and spore abundance in TC banana at flowering |
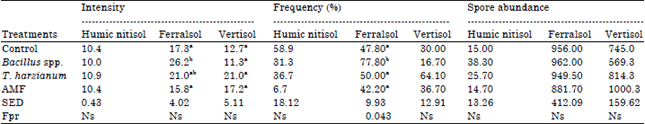 | |
Means within the same column with the same letter are not significantly different (Studentized Tukey HSD test) at p≤0.05 | |
In Rhodic Ferralsol (Coast soil), Bacillus spp. increased the intensity of mycorrhizal colonization of banana roots by 33.9% while in Vertisol (Nyanza soil), AM fungi increased the intensity of mycorrhizal colonization of banana roots by 39.7% when compared to the control. The effect of treatments on the frequency of root AM fungi colonization was only significant (p≤0.043) in Ferralsol, where Bacillus spp. increased the frequency of mycorrhizal colonization of banana roots by 38.6%. Bacillus spp. significantly performed (p≤0.05) better than the control, T. harzianum and AM fungi. In Vertisol, T. harzianum increased the frequency of colonization by 53.2% when compared to the control. The effect of the treatments on spore abundance was not significant (p≤0.05) in the three soils. In Humic Nitisol, Bacillus spp. increased AM fungi spore abundance by 60.8% while T. harzianum increased mycorrhiza spore abundance by 25.5% in Vertisol.
DISCUSSION
Mycorrhizal colonization and spore abundance: The variability in the effect of mycorrhiza, Bacillus and Trichoderma inoculation on mycorrhizal inoculation can be explained according to the high specificity involved in the rhizosphere microbial interactions (Raaijmakers et al., 2009). Negative effect of treatments was detected, since, the control had the most significant effect on the frequency of AM fungi colonization in Humic Nitisol and on the number of AM fungi infective propagules in Humic Nitisol and Rhodic Ferralsol. The situation of negative effect has been described (Germida and Walley, 1996). There was no significant effect of the treatments on spore abundance. This is attributed to the fact that mycorrhizal colonization commences after hyphal formation and subsequent infection of the root. This process may take longer depending on the state of propagules (spores, hyphae), which may take time to germinate and infect the plant (Kavoo-Mwangi et al., 2013).
Fusarium populations under green house and field conditions: The study showed three Fusarium species (Foc, F. proliferatum and F. incarnatum) with broad distribution across geographical regions and soil types in Kenya. The frequency of occurrence of the Fusarium species was variable with some species dominant in some regions more than others. The population of Foc, the causative agent of Fusarium wilt of banana was also variable. Foc was more widely distributed than F. proliferatum and F. incarnatum. Foc was found in the Humic Nitisol, Rhodic Ferralsol and Vertisol soils indicating a high adaptability to soil conditions. The distribution of Foc is determined by the presence of a suitable host and the frequency can be attributed to the intensity of cultivation of the host. The occurrence of Foc in the three Kenyan regions can therefore be attributed to cultivation of various types of banana and plantain varieties in the three regions. Fusarium wilt of banana was reported in susceptible cultivars Gros Michel (AAA) and sweet banana (AAA) in central Kenya, sweet banana in Western Kenya and Bluggoe (ABB) and sweet banana in coastal Kenya (Kung’u et al., 1998). Foc was most frequent in soils from central Kenya (Humic Nitisol) and least isolated from Coast (Rhodic Ferralsol) and Nyanza (Vertisol). This could be attributed to the intensity of cultivation of bananas which is highest in central Kenya, where it is driven by market demands from the capital city of Nairobi, compared to the other two regions (Kung’u et al., 1998; Njuguna et al., 2010). Similarly, the frequency of F. proliferatum followed trends explained by the most prevalent host, maize which is cultivated in all the three regions but at different intensities. It was highest in Vertisol soil where maize and other cereals such as sorghum and millet dominate the landscape (Manyong et al., 2008). F. incarnatum was more frequent in central Kenya and western Kenya soils. There are limited reports on this species with most associating it with cereals (Yli-Mattila, 2010).
Based on the observations on the occurrence, frequency and populations observed in these regions, we evaluated the efficacy of three commercial biological products on Foc and related species. In this study, commercial biological products suppressed the composition, abundance and frequency of Fusarium spp. Emphasis was placed on the suppression of Foc populations. Depending on soil type, Fusarium species responded differently to application of commercial products. All products suppressed Foc populations in the Humic Nitisol while not all products suppressed Foc in the Vertisol and Rhodic Ferrasol soils. The differential functioning of mycorrhizae, Trichoderma and Bacillus in the Humic Nitisol, Rhodic Ferralsol and Vertisol may be attributed to the ability of each soil to support growth and proliferation of the microorganisms around the rhizosphere and subsequent microbial survival and root colonization. Nutrient poor soils harboring soil-borne Fusarium may be difficult to suppress by addition of biological control agents as lack of sufficient soil nutrients or root exudates may limit their proliferation and suppressive abilities. For example, benefits of AM fungi are greatest in soils with >25 mg P kg–1 (Sastry et al., 2000) and they decrease as soil P levels increase beyond 50 mg P kg–1 (Schubert and Hayman, 1986). The Rhodic Ferralsol (7 mg P kg–1) and Vertisol (3 mg P kg–1) have low available P (FAO., 2006) and this may explain the high populations of Fusarium spp. observed even with the mycorrhiza inoculated soils. Humic Nitisol soil had the highest P (8.5 mg P kg–1) among the three field soils and the highest Foc population implying that the P levels in the soils are naturally too low to suppress Foc. There is no information on the threshold P levels required to suppress Foc. However, Rhizatech (containing mycorrhiza) was able to function under P level of 8.5 mg P kg–1 to suppress Foc to a magnitude of 68%. The P level of 7 mg P kg–1 in the Rhodic Ferrasol soil was not adequate for functioning of Rhizatech (though positive effects on mycorrhizal infectivity were evident) and worse still, lower P levels of 3 mg P kg–1 in the Vertisol soil caused Rhizatech to have a negative effect on Foc. Further studies should be undertaken to determine the optimal P levels that would naturally suppress Foc and the levels that would optimize the functioning of Rhizatech.
There were low populations of Foc in the Vertisol, however in our study, Bacillus functioned best in the Vertisol which is inherently clayey, with low P of 3 mg kg–1 and a pH of 5.87. Naturally, clay soil is known to suppress Foc (Sudarma and Suprapta, 2011). Beneficial effects of bacterial inoculation are optimal in nutrient deficient soils than in nutrient rich soils (Egamberdiyeva, 2007). Bacteria inoculation had a much better stimulatory effect on plant growth and nitrogen (N), phosphorus (P) and potassium (K) uptake of maize in nutrient deficient Calcisol soil than in relatively rich loamy sand soil, where bacterial inoculants stimulated only root growth and N, K uptake of roots (Egamberdiyeva, 2007). The Vertisols of western Kenya are rated lower in P levels and high in calcium compared to Humic Nitisol of central Kenya and Rhodic Ferralsol of coastal Kenya. While, there is evidence on conditions under which Bacillus functions best on nutrient uptake, there is limited information on the soil conditions for Foc suppression. Our study has shown that low available P of 3 mg kg–1, high total exchangeable cations (ECEC), (52.5 cmol kg–1) and calcium (25.99 cmol kg–1), magnesium (12.53 cmol kg–1) and sodium (0.19 cmol kg–1) conditions are conducive for suppression of Foc by Bacillus. The suppression of Foc by Bacillus inoculation in the Vertisol may also be explained by the high mycorrhizal colonization. Mycorrhizal symbiosis, through colonization of plant roots, protects plants from soil root pathogens (Dodd, 2000).
Trichoderma thrives best in organically amended soils (Okoth et al., 2007). It is therefore, expected that it would function best under similar conditions. In this study, the amount of carbon was highest in Vertisol (3.87%), Humic Nitisol (2.63%) and least in Rhodic Ferralsol (1%), yet the effect of Trichoderma was best in Humic Nitisol. There is no information on optimal carbon conditions suitable for Trichoderma although we observed Trichoderma to suppress Foc at carbon levels of 2.63%.
Pearson correlation revealed a strong relationship between soil physical and chemical properties and Fusarium populations. There was a negative correlation between sandy soil and Fusarium populations. Clay soil enhanced Fusarium CFUs. This may explain the high Fusarium populations observed in the Humic Nitisol and Vertisol which are clayey soils and low Fusarium populations observed in the Rhodic Ferralsol which is a sandy soil. The positive relationship between the Fusarium populations and soil carbon and nitrogen content may also explain the high populations of Fusarium recovered from the Vertisol and Humic Nitisol. Soil physical and chemical properties have been reported to affect the abundance of Fusarium species. The levels of F. solani f. sp. phaseoli were lower when soil pH decreased and when the levels of Ca, Mg, K and P reduced (Okoth and Siameto, 2011).
CONCLUSION
Treatments had variable effects on beneficial and pathogenic fungi in the rhizosphere of Tissue Culture banana and the effects were soil specific. Therefore, specific treatments should be screened for the different soil types and single strains of microorganisms should be used. The differences observed in the effectiveness of the treatments in the different soils resulted from the varying soil fertility.
Prevailing soil conditions greatly influence the efficacy of the products on Fusarium oxysporum fsp. cubense (Foc) as it is based on their mode of function. It is evident that Humic Nitisol soil provides the best conditions for suppression of Fusarium oxysporum f.sp cubense by the products irrespective of the composition (Mycorrhiza, Bacillus and Trichorderma). This implies that Humic Nitisol soil had conducive conditions for mycorrhiza, Bacillus and Trichorderma while Vertisol soil had conducive conditions for only Bacillus and Rhodic Ferrasol soil were not conducive for all products. This indicates that it is crucial to understand the soil conditions and mode of action of organisms in the product prior to application.
ACKNOWLEDGMENTS
We are grateful to Bill and Melinda Gates Foundation for funding the research, CIAT-TSBF team: Dr. Didier L, Dr. Pypers P and Dr. Sanginga N for technical and administrative support; our supervisors for academic supervision and Mr. Tepeni, Mr. Mwangi and Mr. Mutegi for technical support.
REFERENCES
- Budi, S.W., D. van Tuinen, G. Martinotti and S. Gianinazzi, 1999. Isolation from the Sorghum bicolor mycorrhizosphere of a bacterium compatible with arbuscular mycorrhiza development and antagonistic towards soilborne fungal pathogens. Applied Environ. Microbiol., 65: 5148-5150.
Direct Link - Egamberdiyeva, D., 2007. The effect of plant growth promoting bacteria on growth and nutrient uptake of maize in two different soils. Appl. Soil Ecol., 36: 184-189.
CrossRefDirect Link - Frey-Klett, P., J. Garbaye and M. Tarkka, 2007. The mycorrhiza helper bacteria revisited. New Phytol., 176: 22-36.
PubMed - Gamalero, E., M.G. Martinotti, A. Trotta, P. Lemanceau and G. Berta, 2002. Morphogenetic modifications induced by Pseudomonas fluorescens A6RI and Glomus mosseae BEG12 in the root system of tomato differ according to plant growth conditions. New Phytol., 155: 293-300.
CrossRefDirect Link - Gamalero, E., G. Lingua, F.G. Capri, A. Fusconi, G. Berta and P. Lemanceau, 2004. Colonization pattern of primary tomato roots by Pseudomonas fluorescens A6RI characterized by dilution plating, flow cytometry, fluorescence, confocal and scanning electron microscopy. FEMS Microbial. Ecol., 48: 79-87.
CrossRefDirect Link - Perez, J.G. and M.C. Jaizme-Vega, 1997. Influence of infection by mycorrhizal fungus Glomus intraradices on plant growth and root development of Grande Naine banana. Proceedings of the International Symposium on Banana in the Subtropics, November 10-14, 1997, Puerto de La Cruz, Tenerife, Canary Islands, Spain, pp: 155.
- Geiser, D.M., M. del Mar Jimenez-Gasco, S.C. Kang, I. Makalowska and N. Veeraraghavan et al., 2004. FUSARIUM-ID v. 1.0: A DNA sequence database for identifying Fusarium. Eur. J. Plant Pathol., 110: 473-479.
CrossRefDirect Link - Germida, J.J. and F.L. Walley, 1996. Plant growth-promoting rhizobacteria alter rooting patterns and arbuscular mycorrhizal fungi colonization of field-grown spring wheat. Biol. Fertil. Soils, 23: 113-120.
Direct Link - Hildebrandt, U., K. Janetta and H. Bothe, 2002. Towards growth of arbuscular mycorrhizal fungi independent of a plant host. Applied Environ. Microbiol., 68: 1919-1924.
CrossRefDirect Link - Kavoo-Mwangi, A.M., E.M. Kahangi, E. Ateka, J. Onguso and J.M. Jefwa, 2014. Integration of commercial microbiological products into soil fertility practices as a potential option for acclimatization and growth of TC banana in Kenya. Open J. Soil Sci., 4: 259-271.
CrossRefDirect Link - Kavoo-Mwangi, A.M., E.M. Kahangi, E. Ateka, J. Onguso, R.W. Mukhongo, E.K. Mwangi and J.M. Jefwa, 2013. Growth effects of microorganisms based commercial products inoculated to tissue cultured banana cultivated in three different soils in Kenya. Applied Soil Ecol., 64: 152-162.
CrossRefDirect Link - Khalil, H., A. Brunet, I. Saba, R. Terra, R.P. Sekaly and J. Thibodeau, 2003. The MHC class II β chain cytoplasmic tail overcomes the invariant chain p35‐encoded endoplasmic reticulum retention signal. Int. Immunol., 15: 1249-1263.
Direct Link - Koske, R.E. and J.N. Gemma, 1989. A modified procedure for staining roots to detect VA mycorrhizas. Mycol. Res., 92: 486-488.
CrossRefDirect Link - Manyong, V.M., A.D. Alene, A. Olanrewaju, B. Ayedun and V. Rweyendela et al., 2008. Baseline study of striga control using Imazapyr-Resistant (IR) Maize in Western Kenya. An agricultural Collaborative Study on Striga Control by the African Agricultural Technology Foundation and the International Institute of Tropical Agriculture, Nigeria, pp: 1-55.
- McGonigle, T.P., D.G. Evans and M.H. Miller, 1990. Effect of degree of soil disturbance on mycorrhizal colonization and phosphorus absorption by maize in growth chamber and field experiment. New. Phytol., 116: 629-636.
CrossRefDirect Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Nowak, J., 1998. Benefits of in vitro biotization of plant tissue cultures with microbial inoculants. In vitro Cell. Dev. Biol. Plant, 34: 122-130.
CrossRefDirect Link - O'Donnell, K., H.C. Kistler, E. Cigelnik and R.C. Ploetz, 1998. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA., 95: 2044-2049.
Direct Link - Okoth, S., H. Roimen, B. Mutsotso, E. Muya, J. Kahindi, J.O. Owino and P. Okoth, 2007. Land use systems and distribution of Trichoderma species in Embu region, Kenya. Trop. Subtrop. Agroecosyst., 7: 105-122.
Direct Link - Raaijmakers, J.M., T.C. Paulitz, C. Steinberg, C. Alabouvette and Y. Moenne-Loccoz, 2009. The rhizosphere: A playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant Soil, 321: 341-361.
CrossRefDirect Link - Requena, N., I. Jimenez, M. Toro and J.M. Barea, 1997. Interactions between plant-growth-promoting rhizobacteria (PGPR), arbuscular mycorrhizal fungi and Rhizobium spp. in the rhizosphere of Anthyllis cytisoides, a model legume for revegetation in Mediterranean semi-arid ecosystems. New Phytol., 136: 667-677.
CrossRefDirect Link - Rodriguez-Romero, A.S., M.S.P. Guerra and M.D.C. Jaizme-Vega, 2005. Effect of arbuscular mycorrhizal fungi and rhizobacteria on banana growth and nutrition. Agron. Sustainable Devel., 25: 395-399.
CrossRefDirect Link - Sastry, M.S.R., A.K. Sharma and B.N. Johri, 2000. Effect of an AM fungal consortium and Pseudomonas on the growth and nutrient uptake of Eucalyptus hybrid. Mycorrhiza, 10: 55-61.
CrossRefDirect Link - Schubert, A. and D.S. Hayman, 1986. Plant growth responses to vesicular-arbuscular mycorrhiza: XVI. Effectiveness of different endophytes at different levels of soil phosphate. New Phytol., 103: 79-90.
CrossRefDirect Link - Skujins, J., 1984. Microbial ecology of desert soils. Adv. Microbiol. Ecol., 7: 49-91.
CrossRefDirect Link - Sudarma, M.I. and D.N. Suprapta, 2011. Diversity of soil microorganisms in banana habitats with and without Fusarium wilt symptom. J. Int. Soc. Southeast Asian Agric. Sci., 17: 147-159.
Direct Link - Whipps, J.M., 2001. Microbial interactions and biocontrol in the rhizosphere. J. Exp. Bot., 52: 487-511.
PubMed - Xavier, L.J.C. and J.J. Germida, 2003. Bacteria associated with Glomus clarum spores influence mycorrhizal activity. Soil Biol. Biochem., 35: 471-478.
CrossRefDirect Link - Yli-Mattila, T., 2010. Ecology and evolution of toxigenic Fusarium species in cereals in northern Europe and Asia. J. Plant Pathol., 92: 7-18.
Direct Link - Vazquez, P., G. Holguin, M.E. Puente, A. Lopez-Cortes and Y. Bashan, 2000. Phosphate-solubilizing microorganisms associated with the rhizosphere of mangroves in a semiarid coastal lagoon. Biol. Fert. Soils, 30: 460-468.
CrossRefDirect Link