
Pramod W. Ramteke
Department of Biological Sciences, School of Basic Sciences, Sam Higginbottom Institute of Agriculture, Technology and Sciences, Allahabad 211 007 Uttar Pradesh, India
Babu Joseph
College of Applied medical Sciences, P.B. No. 1383, Shaqra University, Shaqra11961, Kingdom of Saudi Arabia
Abin Mani
Directorate of Research, Sam Higginbottom Institute of Agriculture, Technology and Sciences, Allahabad 211 007 Uttar Pradesh, India
Sonia Chacko
Directorate of Research, Sam Higginbottom Institute of Agriculture, Technology and Sciences, Allahabad 211 007 Uttar Pradesh, India
International Journal of Soil Science
Year: 2012 | Volume: 7 | Issue: 1 | Page No.: 15-27







ABSTRACT
The rhizosphere or the zone of influence around roots harbour a multitude of microorganisms that are affected by both abiotic and biotic stresses. This study investigates the impact of sewage irrigation on culturable rhizobacterial communities of Pisum sativum L. The representative PGPR strains of Bacillus, Pseudomonas, Rhizobium and Azotobacter were tested for plant growth promoting activities and heavy metal tolerance pattern. Rhizobacterial strains isolated from sewage-irrigated soil were more tolerant to heavy metals. The test rhizobacterium Pseudomonas putida was both metal tolerant and efficient in producing PGP compounds like indole acetic acid, ammonia, siderophore and catalase, was subjected to seed germination test. Results confirmed that this bacterium had significant impact on stimulation of root and shoot growth. Our findings suggest that P. putida is a strong candidate for development as soil/seed inoculants to enhance crop yields where untreated sewage water is used in irrigation.
PDF Abstract XML References Citation
Received: October 30, 2011;
Accepted: November 14, 2011;
Published: January 09, 2012
How to cite this article
Pramod W. Ramteke, Babu Joseph, Abin Mani and Sonia Chacko, 2012. Pisum sativum and Associated Plant Growth Promoting Rhizobacteria: Effect of Normal and Sewage Irrigation. International Journal of Soil Science, 7: 15-27.
DOI: 10.3923/ijss.2012.15.27
URL: https://scialert.net/abstract/?doi=ijss.2012.15.27
DOI: 10.3923/ijss.2012.15.27
URL: https://scialert.net/abstract/?doi=ijss.2012.15.27
INTRODUCTION
In India, pea (Pisum sativum L.) is cultivated as winter and summer crop in the plains and hills, respectively. About 0.81 million ha of land is under P. sativum L. cultivation of which about 65% is in the State of Uttar Pradesh. In the year 2001-02 the total pea production in India was 2038,000 mt and the annual N2-fixation by P. sativum was estimated to be around 0.053 lakh tonnes. P. sativum is widely grown as vegetable, pulses, fodder, soil enricher and green manure. It is also considered as a potential candidate for alternate to cereals and usually included in crop intensification practices. Irrigation of agricultural fields with sewage water is increasing and sewage irrigated soil has high levels of heavy metal contamination. Pollution with heavy metals has received a great attention in the last few years. In agricultural soil, heavy metals influence microbial population and metabolic processes (Giller et al., 1998). Studies on long-term exposure of heavy metals showed decrease in microbial diversity and metabolic activities (Smit et al., 1997; Kozdroj and Van Elsas, 2001). Plant roots and soil microbes and their interaction can improve metal bioavailability in rhizosphere (Saravanan et al., 2007). The rhizosphere is a nutrient rich environment and richness of the rhizosphere microbial community is determined by several mechanisms, including plant excretion of specific organic compounds, competition for nutrients and solid attachment sites. Chemical compounds released from the rhizosphere of plants are generally species or cultivar specific and plants are thought to selectively enrich their rhizosphere for microorganisms that can utilize the specific organic compounds. Plant Growth-promoting Bacteria (PGPB) are associated with plants and are commonly present in the environment (Bashan and Holguin, 1998). The widely studied group of PGPB are Plant Growth-promoting Rhizobacteria (PGPR) colonizing the root surfaces and closely adhering soil interface, the rhizosphere (Kloepper et al., 1999). The endophytic colonization of host plant organs and tissues reflects the ability of bacteria to selectively adapt to these specific ecological niches (Gray and Smith, 2005). As a result, the intimate associations between bacteria and host plants are formed without harming the plant (Kloepper et al., 1999). PGPR influences plant growth by several mechanisms such as providing the N source for plant through the N2 fixation; exerting a biological control of soil-borne pathogens as well as producing the plant-stimulating growth substances (phytohormone) that improve growth development by the production of auxin, cytokinin and gibberellins. Moreover, ethylene is known as phytohormone and might inhibit elongation of root in some plants, therefore PGPR containing ACC-deaminase activity might promote plant growth as well as reduce the stress imposed by excessive ethylene to the plant (Piromyou et al., 2011). Although several studies are carried out to investigate the effect of heavy metal pollution on PGPR, very little information is available on rhizosphere of P. sativum irrigated with sewage water. Therefore, we studied the effect of sewage water on abundance, diversity and plant growth promoting traits of rhizobacteria associated with rhizosphere of P. sativum. We also monitored the presence of dominant bacterial population in the rhizosphere of P. sativum irrigated with contaminated water. This study helped us to increase our understanding of the distribution, diversity and composition of microbial communities associated with peas grown under sewage irrigated water.
MATERIALS AND METHODS
Sampling: The rhizosphere soil samples were collected from both normal and sewage irrigated fields growing P. sativum L. from west of Allahabad Agricultural Institute, India. The field is being irrigated with domestic sewage for last 5 years. Randomly located plants were uprooted carefully and the excess of soil was removed by gentle shaking and the soil adhering to roots formed composite samples. The collected samples were placed in plastic bags and kept in an icebox during field survey and sample collection and finally kept at 4°C in the laboratory until processed.
Isolation of rhizobacteria: Soil samples were serially diluted in sterile phosphate-buffered saline (Hi-Media, pH 7.2) and plated on the appropriate medium for isolating different rhizobacteria Vis: Nutrient Agar for Bacillus spp., Kings B medium for Pseudomonas spp. (Ahmad et al., 2008), Yeast Extract Mannitol Agar (YEMA) for Rhizobium spp. (Vincent, 1970) and Ashby’s agar for Azotobacter spp. (Norris and Chapman, 1968). For the isolation of Rhizobium from root nodules, the collected nodules were immersed in 0.1% acidified mercuric chloride, in 70% ethyl alcohol for five minutes for surface sterilization and then washed with sterile distilled water. Crushed root nodules were serially diluted and plated on YEMA medium. After incubation at 28-30°C for 2-3 days, bacterial colonies were counted and representative colonies were selected based on distinct types and observed according to the morphological characteristics such as pigments; colony form, elevation and margin; texture; and opacity (Simbert and Krieg, 1981).
Biochemical characterization of rhizobacteria: Selected isolates of Bacillus (19), Pseudomonas (23), Rhizobium (21), Azotobacter (17) and Rhizobium from nodule (13) were biochemically characterized by Gram’s reaction, carbohydrate fermentation, oxidase test, O-F test, H2S production, IMViC tests, NO2 reduction, starch and gelatin hydrolysis as per the standard methods (Cappuccino and Sherman, 1992).
Heavy metal and physicochemical analysis of soil: Soil samples were analyzed for physicochemical parameters like pH, conductivity, total organic carbon, organic matter, total nitrogen, etc. For determination of heavy metal content soil samples were digested with ternary acid consisting of Nitric acid, Hydrochloric acid and sulphuric acid (5:1:1). The metals Co, Zn, Hg, Cu Cr, Cd, Ni and Pb were estimated by using a Direct Current Plasma (DCP) spectrophotometer.
Heavy metal tolerance: These isolated bacterial strains were tested for their resistance to heavy metals by agar dilution method (Washington and Sutter, 1980). Freshly prepared agar plates were amended with various soluble heavy metal salts namely Co, Zn, Hg, Cu, Cr, Cd, Ni, As and Pb at various concentrations ranging from 25 to 200 μg mL-1 were inoculated with overnight grown cultures. Heavy metal tolerance was determined by the appearance of bacterial growth after incubating the plates at room temperature for 24-48 h.
Characterization of rhizobacteria for PGP traits: Rhizobacterial isolates were characterized for plant growth promoting characteristics based on the standard procedures.
Indole acetic acid: The bacterial strains were cultured in test tubes for 24 h containing LB medium amended with 50 mg mL-1 of tryptophan. After incubation 2 mL of cell suspension was centrifuged at 1000 rpm for 10 min and 2-3 drops of orthophosphoric acid were added to the supernatant along with 4 mL of Solawaski’s reagent. The tubes were kept at room temperature for 20 min. Indole Acetic Acid (IAA) production was indicated by the development of pink colour. Optical density was read at 530 nm and level of IAA production was estimated by standard IAA graph (Bric et al., 1991).
Ammonia production: The isolates were grown in peptone water at 30°C for 4 days and 1 mL of Nessler’s reagent was added. Production of ammonia was represented by development of faint yellow colour (Bakker and Schippers, 1987).
Siderophore production: Siderophore production was detected by the universal method of (Schwynn and Neilands, 1987) using blue agar plates containing the dye chrom azurol S (CAS). Orange halos around the colonies on blue were indicative for siderophore production.
Catalase production: Bacterial cultures were grown in a nutrient agar medium for 18-24 h. The cultures were mixed with appropriate amount of H2O2 on a glass slide to observe the evolution of oxygen.
Phosphate solubilization: The isolates were tested for phosphate solubilizing ability on Pikovskaya medium (Nautiyal, 1999) incorporated with tricalcium phosphate (Ca3(PO4)5). Phosphate solubilization was indicated by the formation of a clear halo zone around the bacterial growth after three days of incubation.
HCN production: The isolates were streaked on King’s B medium amended with 4.4 g L-1 of glycine. The plates were covered with sterile filter paper impregnated with 0.5% picric acid in 2% sodium carbonate, sealed with parafilm and incubated for 4 days (Bakker and Schippers, 1987). Development of yellow colour on the filter paper indicated positive result.
Seed germination test: Surface sterilized and uniform size maize (Zea mays L.) seeds dipped in gum acacia were inoculated with test bacterium Pseudomonas putida and placed in pertridishes. Treated and untreated seeds were kept for germination in dark for two days in an incubator at 27±2°C. Germination of treated and untreated seeds was recorded and the germinated seeds were transferred into nitrogen free minimal agar medium containing heavy metals (100 μ mL-1). After seventh day of incubation, the seedlings were taken out for various studies like root and shoot length and the data were recorded. Seeds without coating P. putida were maintained as control.
Plasmid curing: Representative strains were subjected to acridine orange mediated plasmid elimination (Marcelo et al., 1993). Overnight logarithmically growing culture was inoculated in nutrient broth containing acridine orange at various concentrations ranging from 10-100 μg mL-1. After incubation for 48 h, the tube showing turbidity and containing higher concentration of curing agent was serially diluted and plated in nutrient agar plates. Individual colonies were screened for plasmid-encoded traits.
RESULTS AND DISCUSSION
The contamination of the environment with toxic metals has become a worldwide problem, affecting crop yields, soil biomass and fertility contributing to bioaccumulation in the food chain. Although, organic content in sewage increases soil fertility, it also contributes to soil contamination with heavy metals (Giller et al., 1984; Muller et al., 2001). In the present study we noticed that microbial biomass is unaffected by sewage irrigation (Table 1). The numbers of Bacillus in normal and sewage-irrigated soil were around 4.0x106 and 1.0x106, respectively. There was no much difference in the bacterial count of both hetrotrophs and nitrogen fixers in both soil types. Several studies have documented that heavy metal contamination results in the reduction of bacterial diversity, biomass and metabolic activity (Kandeler et al., 1996; Ellis et al., 2003). It is known that heavy metal pollution causes selection and/or development of tolerant microorganisms (Baath, 1989). However, contradictory results are also available on effect of wastewater or sewage on soil microbial biomass. Microbial biomass is either unaffected by wastewater application or increased after amendment with sewage sludge (Martin-Laurent et al., 2004). In the present investigation, we also observed negligible depletion of rhizobacterial population was noticed as compared to sewage irrigated rhizosphere of P. sativum and rhizosphere irrigated with normal water.
| Table 1: | Microbiological analysis of soil samples collected from rhizosphere of P. sativum |
 | |
Rhizobacterial isolates of Bacillus (19), Pseudomonas (23), Rhizobium (21) and Azotobacter (17) and Rhizobium from nodule (13), from both normal and sewage-irrigated rhizosphere were isolated and selected for the present study. To maintain plant health, it is important to control the optimum constitution of rhizobacterial species in the rhizosphere. For this reason, many efforts have been made to characterize the structure and function of rhizobacterial communities. Most important is their contribution to the overall N economy of soil by sequestering atmospheric N through symbiotic N2-fixation and through subsoil N retrieval (Gattumbi et al., 2002). The effect of legume rhizodeposition on bacterial communities, showed a distinct plant-dependant rhizosphere effect on the distribution of different bacterial groups present in legume rhizosphere (Sharma et al., 2005). Rhizobia are gram-negative soil bacteria of particular interest due to their symbiotic nitrogen fixing ability with members of leguminaceae (Saleena et al., 2001). Many abiotic factors affect the growth and survival of Rhizobia in soil. Therefore, for the good growth of legumes in semiarid and arid regions, it is necessary that the plant is being nodulated by an effective and stress tolerant strain of Rhizobium. Rhizobium spp. with genetic potential for increased tolerance to heavy metals could enhance production of food and forage legumes in semiarid and arid regions of the world. In the developing countries, fast-growing legumes are required for both fodder and sustainable fuel wood production. We observed that the levels of total organic carbon, organic matter and heavy metals in sewage-irrigated rhizosphere increased three fold or more as compared to normal soil (Table 2). Organic carbon content of the sewage irrigated soil is 67.65% greater than the normal soil. This may be due to the exudates, sloughing of root tissues during root death and senescence (Graham and Haynes, 2006). The presence of high organic content in the rhizosphere promotes microbial proliferation.
Quantification of heavy metal content revealed a strong contamination of the soil in proportion to amount of the waste water applied. As expected, tolerance to heavy metals was found more predominant among rhizobacteria from sewage-irrigated soil as compared to normal except for Nickel. Among all the rhizobacteria studied, tolerance to heavy metals was observed less frequently in Azotobacter spp. (Fig. 1). None of the isolates of Azotobacter spp. were tolerant to Cu and Pb. Azotobacter spp. isolated from normal soil were fully susceptible to Cr, As, Cd and Zn. Surprisingly, none of the isolates of Azotobacter spp. from sewage irrigated rhizosphere were tolerant to Nickel. Whereas 50% isolates of Azotobacter spp. from normal soil were tolerant to Ni.
| Table 2: | Heavy metals content and physicochemical analysis of soil samples |
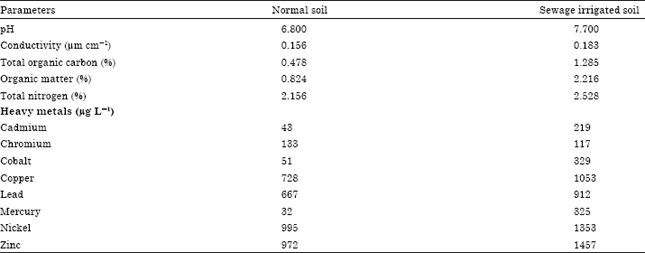 | |
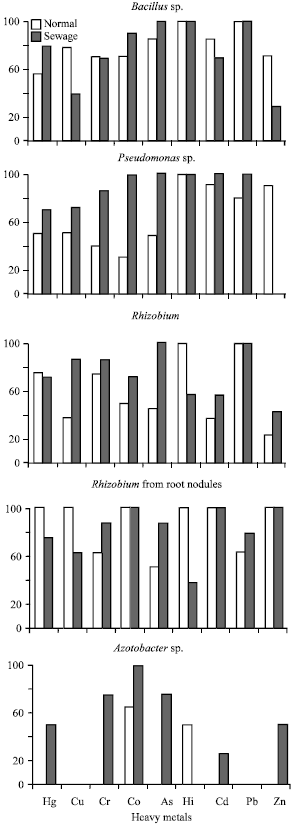 | |
| Fig. 1: | Heavy metal tolerance shown by the isolates of rhizobacteria isolated from normal and sewage irrigated P. sativum (Heavy metals μg mL-1 Hg 50; Cu 100; Cr 100; Co 100; As 100; Ni 100; Cd 100; Pb 200; Zn 100) |
The dominance of rhizobacteria in the normal soil was Bacillus spp. whereas, sewage irrigated soil was represented by Pseudomonas spp. However, some isolates obtained from the normal rhizosphere were tolerant towards heavy metals such as of Bacillus spp. (Cu and Zn), Pseudomonas spp. (Zn) and Rhizobium (Cu) as compared to the isolates from sewage irrigated soil (Table 3). Heavy metal tolerances among different rhizobial strains were reported earlier (Giller et al., 1984; El-Aziz et al., 1991). Several workers reported adverse effect of heavy metal pollution on PGP characteristics. Heavy metals, at higher concentration, are toxic to cells and may cause cell death by interacting with nucleic acids and enzymes active site (Hazel and Williams, 1990; Cervantes and Gutierrez-Corana, 1994). Significant reductions in rhizobial numbers occurred even at metal concentrations well below the permissible limit (Chaudri et al., 1993).
Rhizobacteria tolerant to multiple heavy metals exhibited a couple of PGP activities (Table 4). PGPR increase plant growth by decreasing heavy metal toxicity (Burd et al., 1998). Rhizobacteria which establish positive interactions with plant roots, PGPR play a key role in agricultural environments and are promising for their potential use in sustainable agriculture (De Fago et al., 1994). It was also apparent that more cultures of PGPR isolated from sewage-irrigated rhizosphere were tolerant to elevated levels heavy metals. However, data of this investigation seem to indicate that application of wastewater which exposes soil micro biota to heavy metal, did not alter PGP abilities. Microbial resistance to metals is generally associated with plasmids. The tolerance to heavy metals was found not curable when acridine orange was used as curing agent suggesting chromosomal traits (data not shown).
It is clear that sewage irrigation did not have profound inhibitory influence on PGP characteristics of rhizobacteria. On the other hand isolates from sewage-irrigated soil exhibited plant growth promoting activities as compared to their counterparts isolated from soil irrigated with normal water. All the rhizobacteria isolated from both normal and sewage irrigated rhizosphere of P. sativum were found to produce plant growth hormone IAA and catalase. Bacterial IAA plays a major role in promotion of root elongation when a bacterium is associated with its host plant. IAA secreted by a bacterium may promote root growth directly by stimulating plant cell elongation or cell division or indirectly by influencing bacterial ACC deaminase activity. Catalase activity in the bacterial strains may potentially be very advantageous.
| Table 3: | Heavy metal tolerance of selected rhizhobacterial isolate |
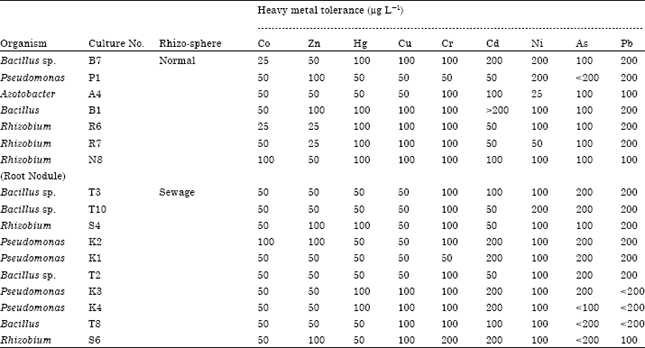 | |
Bacterial strains showing catalase activity must be highly resistant to environmental, mechanical and chemical stress. Production of ammonia was detected in all Bacillus spp., in >80% Psuedomonas spp. and Rhizobium spp. (root nodules) isolated from normal rhizosphere (Fig. 2).
| Table 4: | Plant growth promoting characteristics |
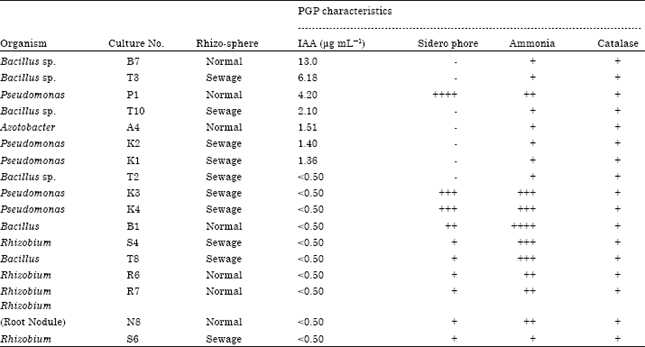 | |
| +: Poor; ++: Fair; +++: Good;+ +++: Excellent | |
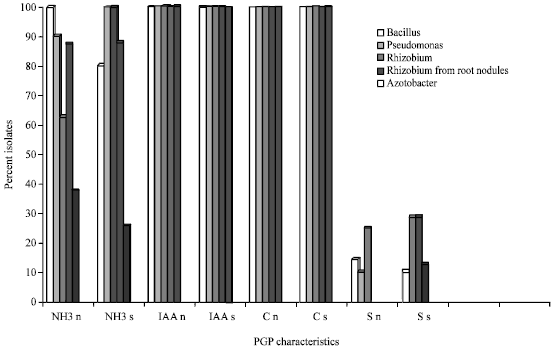 | |
| Fig. 2: | Rhizobacterial isolates showing plant growth promoting characteristics NH3-Ammoniaproduction; IAA-Indole acetic acid production; C-Catalase positive; S-Siderophore production; n-normal soil; s-sewage irrigated soil |
Production of ammonia was found less frequent in Azotobacter spp. as compared to other rhizobacteria. Higher number of isolates of Rhizobium spp. and Psuedomonas spp. from sewage irrigated rhizosphere of P. sativum showed significant production of ammonia as compared to their counterparts from normal soil. Production of siderophore was detected less frequently than other PGP characteristics. Production of siderophore was detected significantly higher among isolates of Psuedomonas spp. Three isolates of Pseudomonas spp. were strong siderophore producers and out of them, two were isolated from sewage rhizosphere. Siderophore may indirectly stimulate the biosynthesis of other antimicrobial compounds by increasing the availability of these minerals to the bacteria. Siderophore may further function as stress factor or signals inducing local and systematic host resistance (Leeman et al., 1996). PGPR can suppress the growth of pathogenic organisms directly by producing either antibiotic compound (Kloepper, 1993) or siderophores that allowed effective iron capture from the rhizosphere, depriving pathogenic organisms of this necessary element (Loper and Buyer, 1991). None of the isolates of Azotobacter spp. produced siderophore. Production of HCN and Phosphate solubilizing activity were not detected in any of the isolates.
In this study, one of the potential bacterial isolate (P1)which was identified as P. putida, tolerant to multiple metals (As, Pb and Ni) and produced IAA, siderophore and ammonia was subjected to elucidate its role in stimulation of root and shoot growth in presence of heavy metals. Promotion of root growth is one of the major markers by which the beneficial effect of plant growth promoting bacteria is measured (Glick et al., 1995). Rapid establishment of roots, whether by elongation of primary roots or by proliferation of lateral roots, is advantageous for young seedlings as it increases their ability to anchor themselves to the soil and to obtain water and nutrients from their environment, thus enhancing their chances for survival. Stimulation in formation of lateral roots is attributed to high level of bacterial IAA while low levels of IAA stimulate root elongation (Xie et al., 1996; Patten and Glick, 2002). From the results obtained from seed germination test, it is apparent that the bacterium had significant impact on stimulation of root and shoot growth (Table 5). Roots from seeds treated with bacterial culture were an average of 42% longer than the roots from untreated control seeds after seven days. Influence of PGP activity on shoot length was evaluated in presence of metals (Table 6). Maize seedlings treated with bacterium exhibited enhanced shoot length in all experiments. Heavy metal resistant microorganisms which grow not only under contaminated environment but also possess growth promoting properties, are of particular importance for the degraded and/or polluted land-use practices.
| Table 5: | Effect of P. putida on shoot and root growth in maize |
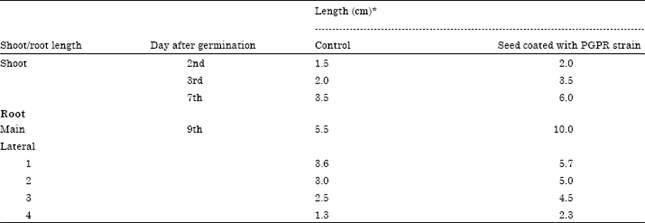 | |
| *Average of triplicate | |
| Table 6: | Effect of P. putida on root growth in maize in presence of heavy metals |
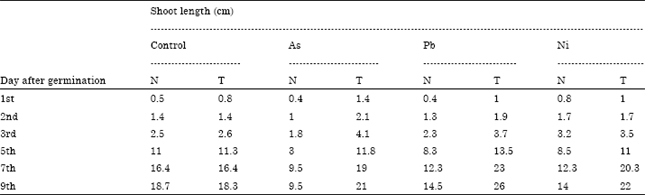 | |
| N: Normal seeds; T: Treated seeds; Heavy metals 100 μg mL-1 | |
Chromium-resistant pseudomonads, isolated from paint industry effluents, were able to stimulate seed germination and growth of Triticum aestivus in the presence of potassium dichromate (Hasnain and Sabri, 1996). From the present study the rhizobacterium P. putida may be a strong candidate for development as soil inoculant to enhance crop yield. Bacterial inoculants are able to increase plant growth and germination rate, improve seedling emergence, responses to external stress factors and protect plants from disease (Lugtenberg et al., 2002).
CONCLUSION
The present study clearly indicates that the rhizobacterial population was not affected by the sewage irrigation. All the isolated strains possessed one or more PGPR traits. These isolates were tolerant to various levels of heavy metals and the representative strain of P. putida was outstanding for PGP potential. The knowledge of plant associated bacteria of rhizosphere is not only for understanding their ecological role and the interaction with the plant but also for future biotechnological applications. The mutual influence between rhizosphere environment and genetic diversity patterns of local microbial populations should be studied in detail.
REFERENCES
- Ahmad, F., I. Ahmad and M.S. Khan, 2008. Screening of free-living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol. Res., 168: 173-181.
CrossRefDirect Link - Baath, E., 1989. Effects of heavy metals in soil on microbial processes and populations: A review. Water Air Soil Pollut., 47: 335-379.
CrossRefDirect Link - Bakker, A.W. and B. Schippers, 1987. Microbial cyanide production in the rhizosphere in relation to potato yield reduction and Pseudomonas SPP-mediated plant growth-stimulation. Soil Biol. Biochem., 19: 451-457.
CrossRefDirect Link - Bashan, Y. and G. Holguin, 1998. Proposal for the division of plant growth-promoting rhizobacteria into two classifications: Biocontrol-PGPB (plant growth-promoting bacteria) and PGPB. Soil Biol. Biochem., 30: 1225-1228.
CrossRefDirect Link - Bric, J.M., R.M. Bostock and S.E. Silverstone, 1991. Rapid in situ assay for indoleacetic acid production by bacteria immobilized on a nitrocellulose membrane. Applied Environ. Microbiol., 57: 535-538.
PubMedDirect Link - Burd, G.I., D.G. Dixon and B.R. Glick, 1998. A plant growth promoting bacterium that decreases nickel toxicity in seedlings. Applied Environ. Microbiol., 64: 3663-3668.
Direct Link - Cervantes, C. and F.Gutierrez-Corana, 1994. Copper resistance mechanism in bacteria and fungi. FEMS Microbiol. Rev., 14: 121-137.
PubMedDirect Link - Chaudri, A.M., S.P. McGrath, K.E. Giller, E. Rietz and D. Sauerbeck, 1993. Enumeration of indigenous Rhizobium leguminosarum biovar trifolii in soils previously treated with metal-contaminated sewage sludge. Soil Biol. Biochem., 25: 301-309.
CrossRefDirect Link - Patten, C.L. and B.R. Glick, 2002. Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Applied Environ. Microbiol., 68: 3795-3801.
CrossRefPubMedDirect Link - El-Aziz, R., J.S. Angle and R.L. Chaney, 1991. Metal tolerance of Rhizobium meliloti isolated from heavy metal contaminated soils. Soil Biol. Biochem., 23: 795-798.
CrossRefDirect Link - Ellis, R.J., P. Morgan, A.J. Weightman and J.C. Fry, 2003. Cultivation-dependent and independent approaches for determing bacterial diversity in heavy metal contaminated soil. Applied Environ. Microbiol., 69: 3223-3230.
PubMedDirect Link - Gattumbi, S.M., K.E. Giller and G. Cadisch, 2002. Do mixed species improved follows increase above and below ground resources capture? J. Agron., 94: 518-526.
Direct Link - Giller, K.E., S.P. McGrath and P.R. Hirsch, 1984. Absence of nitrogen fixation in clover grown on soil subject to long-term contamination with heavy metals is due to survival of only ineffective Rhizobium. Soil Biol. Biochem., 21: 841-848.
CrossRefDirect Link - Giller, K.E., E. Witter and S.P. Mcgrath, 1998. Toxicity of heavy metals to microorganisms and microbial processes in agricultural soils: A review. Soil Biol. Biochem., 30: 1389-1414.
CrossRefDirect Link - Glick, B.R., D.M. Karaturovic and P.C. Newell, 1995. A novel procedure for rapid isolation of plant growth promoting pseudomonads. Can. J. Microbiol., 41: 533-536.
CrossRefDirect Link - Graham, M.H. and R.J. Haynes, 2006. Organic matter status and the size, activity and metabolic diversity of the soil microbial community in the row and inter-row of sugarcane under burning and trash retension. Soil Biol. Biochem., 38: 21-31.
Direct Link - Gray, E.J. and D.L. Smith, 2005. Intracellular and extracellular PGPR: Commonalities and distinctions in the plant-bacterium signaling processes. Soil Biol. Biochem., 37: 395-412.
CrossRefDirect Link - Kandeler, E., C. Kampichler and O. Horak, 1996. Influence of heavy metals on the functional diversity of soil microbial communities. Biol. Fertil. Soils, 23: 299-306.
CrossRefDirect Link - Kloepper, J.W., R. Rodriguez-Kabana, J.W. Zehnder, J. Murphy, E. Sikora and C. Fernandes, 1999. Plant root-bacterial interactions in biological control of soil borne diseases and potential extension to systemic and foliar diseases. Australas. Plant Pathol., 28: 21-26.
Direct Link - Kozdroj, J. and J.D. van Elsas, 2001. Structural diversity of microorganims in chemically perturbed soil assessed by molecular and cytochemical approaches. J. Microbiol. Methods, 43: 197-212.
PubMed - Lugtenberg, B.J.J., T.F.C. Chin-A-Woeng and G.V. Bloemberg, 2002. Microbe-plant interactions: Principles and mechanisms. Antonie van Leeuwenhoek, 81: 373-383.
CrossRefDirect Link - Martin-Laurent F., L. Cornet, L. Ranjard, J. Lopez-Gutierrez and L. Philippot et al., 2004. Estimation of atrizine-degrading genetic potential and activity in three French agricultural soils. FEMS Microbiol. Ecol., 48: 425-435.
PubMed - Nautiyal, C.S., 1999. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett., 170: 265-270.
CrossRefPubMedDirect Link - Piromyou, P., B. Buranabanyat, P. Tantasawat, P. Tittabutr, N. Boonkerd and N. Teaumroong, 2011. Effect of plant growth promoting rhizobacteria (PGPR) inoculation on microbial community structure in rhizosphere of forage corn cultivated in Thailand. Eur. J. Soil Biol., 47: 44-54.
CrossRefDirect Link - Saleena, L.M., P. Loganathan, S. Rangarajan and S. Nair, 2001. Genetic diversity of Bradyrhizobium strains isolated from Arachis hypogaea. Can. J. Microbiol., 47: 118-122.
Direct Link - Saravanan, V.S., M. Madhaiyan and M. Thangaraju, 2007. Solubilization of zinc compounds by the diazotrophic, plant growth promoting bacterium Gluconacetobacter diazotrophicus. Chemosphere, 66: 1794-1798.
CrossRef - Schwynn, B. and J.B. Neilands, 1987. Universal chemical assay for detection and determination of siderophores. Anal. Biochem., 169: 47-56.
PubMedDirect Link - Sharma, S., M. Aneja, J. Mayer, M. Schloter and J.C. Munch, 2005. Characterization of bacterial community structure in rhizosphere soil of grain legumes. Microbiol. Ecol., 49: 407-415.
PubMed - Smit, E., P. Leeflag and K. Wernars, 1997. Detection of shifts in microbial community structure and diversity in soil caused by copper contamination using amplified ribosomal DNA restriction analysis FEMS Microbiol. Ecology, 23: 249-261.
CrossRefDirect Link - Xie, H., J.J. Pasternak and B.R. Glick, 1996. Isolation and characterization of mutants of the plant growth-promoting rhizobacterium Pseudomonas putida GR12-2 that overproduce indoleacetic acid. Curr. Microbiol., 32: 67-71.
CrossRefDirect Link - Hazel, J.R. and E.E.Williams, 1990. The role of alterations in membrane lipid composition in enabling physiological adaptation of organisms to their physical environment. Prog. Lipid Res., 29: 167-227.
PubMed