
Girma Abera
Department of Plant and Environmental Sciences, Norwegian University of Life Sciences, P.O. Box 5003, N-1432 Aas, Norwa
Endalkachew Wolde-meskel
School of Plant and Horticultural Sciences, Hawassa University, P.O. Box 05, Hawassa, Ethiopia
Sheleme Beyene
School of Plant and Horticultural Sciences, Hawassa University, P.O. Box 05, Hawassa, Ethiopia
Lars R. Bakken
Department of Plant and Environmental Sciences, Norwegian University of Life Sciences, P.O. Box 5003, N-1432 Aas, Norwa
International Journal of Soil Science
Year: 2012 | Volume: 7 | Issue: 4 | Page No.: 132-145







ABSTRACT
This study was conducted to assess nitrogen (N) mineralization from Soil Organic Matter (SOM) in situ and in laboratory incubation under different soil moisture regimes. Andosol soils were sampled from coffee (CF) and crop (CP) land uses of southern Ethiopia. Extractable NO3¯-N and mineral N were strongly increased, while NH4+-N declined in response to soil moisture increase (from air dry to 100% FC, field capacity) during laboratory incubation. In situ extractable NO3¯-N and mineral N were strikingly low in December (dry) with both CF and CP. However, they were consistently greater during March to August due to more rain flushes, suggesting better N release during cropping season. Increasing soil moisture from air dry to 100% FC enhanced extractable NO3¯-N and mineral N of CF by 95.7 and 81.7%, respectively, while that of CP was increased by 58.7 and 46.4% respectively. By contrast, extractable NH4+-N declined by 82.9% with CF and by 87.0% with CP. The assessment of N mineralization in relation with soil moisture showed consistent trends both under in situ and laboratory methods. Indeed, the N mineralization of CF was about double to that of CP at 75-100% FC and during July-August (wet season). In contrast, average in situ N mineralization of CP was greater than CF, attributed to longer history of annual inputs of N-fertilizer and might be because of absence of actively competing plants with soil microorganisms. Generally, the knowledge of annual patterns of N mineralization in relation with soil moisture is necessary to synchronize crop N demands with plant-available N in the soil.
PDF Abstract XML References Citation
Received: April 10, 2012;
Accepted: June 19, 2012;
Published: August 13, 2012
How to cite this article
Girma Abera, Endalkachew Wolde-meskel, Sheleme Beyene and Lars R. Bakken, 2012. Nitrogen Mineralization Dynamics under Different Moisture Regimes in Tropical Soils. International Journal of Soil Science, 7: 132-145.
DOI: 10.3923/ijss.2012.132.145
URL: https://scialert.net/abstract/?doi=ijss.2012.132.145
DOI: 10.3923/ijss.2012.132.145
URL: https://scialert.net/abstract/?doi=ijss.2012.132.145
INTRODUCTION
Soil Organic Matter (SOM) is a major factor to determine whether soils are sink or source of C in the global cycling (Post and Kwon, 2000). Land use, soil type, climate and vegetation are the drivers of SOM dynamics (Feller and Beare, 1997). The microbial activity that recycles the nutrients on which plant productivity depends is highly sensitive to water availability.
Soil organic matter decomposition and N mineralization depend on the quantity and quality of organic matter, soil conditions, temperature and moisture (Goncalves and Carlyle, 1994). Soil climatic conditions vary in different ecosystems and seasons, this in turn affect the soil microbial activity and biomass (Singh et al., 1989; Michelsen et al., 2004; Sugihara et al., 2010). Consequently, the overall N cycle is affected in terrestrial ecosystem (Paul and Clarke, 1996). The major factors that limit N mineralization and nitrification are environmental parameters such as temperature, aeration, soil moisture, pH and soil type (Owena et al., 2003). Soil moisture is the most important factor that regulates N mineralization and nitrification by modifying substrate availability and affecting osmotic potential. Consequently, water potential alters the rate of SOM mineralization (Stark and Firestone, 1995; De Neve et al., 2003). Fluctuations in environmental conditions (temperature, moisture and aeration) affect N mineralization and nitrification by altering microbial population size (Jha et al., 1996). Vegetation also affects N mineralization and nitrification through both litter quality and quantity. It is obvious that temperature is not a limiting factor in tropical climates; rather soil moisture is the most critical factor that limits microbial activities during the dry season of the tropics (Abera et al., 2012).
The rate of supply of available N generated by N mineralization involves the microbial conversion of more complex organic N into simpler available mineral-N forms (NH4+-N+NO3¯-N). This implies that soil microorganisms exert significant influence on ecosystem functions by regulating litter decomposition and nutrient cycling (Jenkinson and Ladd, 1981). The rates of soil N mineralization and nitrification not only govern the availability of mineral N for plant growth but also indicate the ability of soil to retain N (Haynes, 1986). Perhaps efficient N recycling in these dry agroecosystems depends on a synchrony between the production of mineral N and plant N uptake. For example accumulated NO3 during the dry season, could easily leach and/or denitrified if large rain events follow, complicating the ability to meet goals of agricultural management or this could reduce synchrony of production of mineral N and plant N uptake. In general, exploring the effect of drought stress on soil N mineralization and N release help to understand the rate of decomposition in relation with N requirement of the crops. Furthermore, understanding the rate of N mineralization potential of soils could help to identify and develop appropriate management interventions required to maximize the N use efficiency and thereby reduce the N fertilizer cost and loss to environment.
Most of previous studies evaluated seasonal N mineralization dynamics in the dry tropics of forest and savanna land uses, whereas little information has been acquired for crop and coffee land uses (Kushwaha et al., 2000; Singh and Kashyap, 2007; Sugihara et al., 2010). However, coffee and crop land uses have contrasting vegetation cover and acquire different management practices that could affect soil organic C stocks, aggregation, bulk density, water retention, aeration, soil structure development and in turn microbial biomass and activity. In general, microbial activity during the dry season in tropical climate is presumed to be marginal due to drought stress, however, evidences are meager. Therefore, the objective of the present study was to explore the effect of drought stress on N mineralization dynamics of coffee and crop land soils under in situ and laboratory incubation.
MATERIALS AND METHODS
Site description: Soil samples were collected from coffee (CF) and crop (CP) land use systems of southern Ethiopia, which were different in terms of land use, vegetation cover and management practices. The agroecosystem covers about 1225 km2, situated at 1708-1950 m above sea level and located 7°3' to 7°6'N and 38°7' to 38°28'E. Basically, the soils of both land uses represent tropical Andosol (An1 refers for CP and An2 refers for CF) and they were loam in texture (Abera et al., 2012). The Andosol was formed from the parental materials of volcanic origin during tertiary periods of geologic time (FAO, 1998). The CF receives mean annual rain fall of 1105 mm and average monthly temperature of 19.4°C, whereas CP receives 952 mm and 19.4°C of average monthly temperature. The study sites are seasonally dry, mainly during November to February and then receive small amount of rains (short rainy season) during March to May, followed by long rainy season (large amount) during June to September (Fig. 1).
The CP land soil was in fallow condition at the time of sampling, while CF (Coffea arabica L.) soil was with perennial mixed agroforestry trees as a shade to coffee. The agronomic management practices applied to coffee (perennial) was completely different from crop land (annual crops). For instance, there is no frequent tillage in coffee plots, rather there is only once hoeing or manual cultivation per season. In addition, coffee production is largely based on organic fertilizer sources such as litter fall, farmyard manure and compost, while annual crop farming was based on chemical fertilizers supply of 92 kg ha-1 N and 20 kg ha-1 P.
Soil sampling and pretreatment of samples for laboratory incubation: Surface layer (0-15 cm) soil samples were collected from multiple positions and then bulked into three separate replicates per each site. The sampled soils were transported to Hawassa University and air dried under ambient temperature. Later, soil samples were transported to Norwegian University of Life Sciences (UMB), Norway and used for laboratory incubation experiment (see details in laboratory incubation section). The soils were kept open aerated under ambient temperature in a laboratory until required physical and chemical measurements were carried out.
Soil water regulation for laboratory incubation: The soil moisture for laboratory incubation included wide moisture regimes that expected to encompass the seasonal soil moisture contents of the land uses. The soil moisture was first set at pF 2 (FC, Field capacity) using a pressure plate apparatus and then the required soil moisture levels (air dry, 25% FC, 50% FC, 75% FC and 100% FC) were set by proportionally mixing wet and air dry soils. Air content of soil samples were measured by air pycnometer at field capacity. The bulk density was determined using a disturbed soil samples in core sampler by oven drying at 105°C for 24 h.
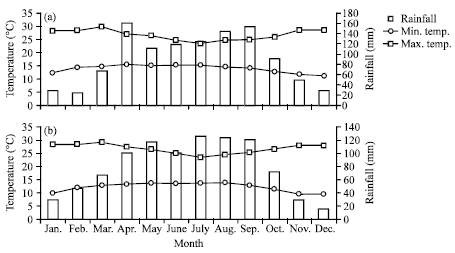 | |
| Fig. 1(a-b): | Long term monthly rainfall and monthly minimum and maximum temperature (°C) of (a) Coffee; CF (An2) and (b) Crop; CP (An1) land uses in southern Ethiopia |
Selected soil properties: The soil organic C content was determined through dry combustion by Allison method (Nelson and Sommers, 1982), whereas, total nitrogen was determined by Dumas method (Bremner and Mulvaney, 1982). Extractable P was determined by Bray’s method (Bray and Kurtz, 1945). Soil pH was determined with a pH electrode at soil water ratio of 1:2.5 (pH H2O and pH CaCl2).
In situ soil sampling and N mineralization: Three fixed experimental plots of 5x5 m were set at each site in three replications from where unconfined soil samples were collected for NH4+-N and NO3¯-N determination during December 2009 to August 2010. In situ N mineralization was periodically measured from two different land uses (CF and CP) as described above. Five soil samples were collected using augur soil sampler from 0-15 cm depth in 30 days interval at each plot. A total of 15 samples per each site were collected in plastic bags and immediately sealed to avoid moisture loss and transported to the laboratory during every sampling period. The samples were either immediately extracted or kept in refrigerator at 4°C and then extracted the next day. Ammonium and nitrate were measured from five replicate soil samples, by extracting ~10 g dry soil with 25 mL of 2 M KCl. Soil slurries were shaken in 120 mL flask for 30 min (reciprocal shaker; 180 strokes per min), settled for 10 min and filtered through Whatman No. 42 filter paper and then filtrate was collected in 15 mm tube. The filter paper was pre-washed with 0.2 M KCl to avoid any mineral N contamination. The NH4+-N and NO3¯-N content were measured by colorimetric method using spectrophotometer.
Laboratory incubation and N mineralization: The N mineralization (NH4+-N and NO3¯-N) under laboratory incubation was determined using soils sampled from CF and CP land uses of southern Ethiopia. The five moisture regimes (air dry, 25% FC, 50% FC, 75% FC and 100% FC) were set by proportional mixing air dry and wet soils and incubated by adding into 50 mL tubes. The tubes were immediately capped to avoid moisture loss and were incubated at 15°C for 45 days. The soils were factorially arranged in completely randomized design of 2x5x3, two land uses, five soil moisture regimes and three replications, respectively.
The soil solution was made by adding ~10 g dry soil into 25 mL of 2 M KCl. Then the soil samples were manually shaken for few seconds followed by gentle shaking of flasks on horizontal shaker for 30 min at 180 rpm. The soil particles were allowed to settle for awhile and filtered through Whatman No. 42 filter paper and the filtrate was collected in 15 mm tube. The NH4+-N and NO3¯-N contents were determined using spectrophotometer (NSF, 1975) and flow injection analyzer method (EPA, 1979), respectively.
Soil moisture determination in the field condition: The surface (0-15 cm) layer soil moisture contents were measured in the field using time domain reflectometry, TDR (HH2 meter with SM200 soil moisture sensor, Delta-T Devices Ltd., UK) in five replicates during December to August in 30 days interval. The soil moisture measurement was randomly sampled from fixed 5x5 m plot area. Concurrently, from the same plot soil samples were randomly collected from 5x5 m plot for NH4+-N and NO3¯-N determination at 30 days interval.
Experimental design and treatments: For in situ sampling, the treatments were factorially arranged into 2x9x5, two land use, nine sampling months and five replications, respectively at the fixed plots of 5x5 m. The sampling months partly included dry and wet season tropical climates. Whereas, soils for laboratory incubation was organized into 2x5x3, two land use, five soil moisture regimes and three replications, respectively. Both experiments were set in Completely Randomized Design (CRD).
Statistical analyses: The effects of drought stress on N mineralization and nitrification were analyzed based on one and two way analyses of variance using GLM Model in MINITAB Statistical Software for Windows Release 14 (Minitab, State College, Pa.). The relationship between drought stress effect and N mineralization and nitrification were estimated by linear regression.
RESULTS
Selected soil properties: Significant differences (p<0.05) were observed in Soil Organic Carbon (SOC) and total N concentrations between land use systems that varied from 14 g kg-1 with CP to 32 g kg-1 with CF. Similarly, total N concentration was in the range of 1.5 to 2.7 g kg-1. The textural class of the studied soils was loamy (Table 1). The initial soil pH (H2O) ranged from 5.9 to 6.8. There were significant variations in soil pH among sampling date (p<0.05), between land uses (p<0.05) and sampling date x land use interaction (p<0.05). The soil pH tended to decline from December to August sampling period (Fig. 2). The soils revealed low bulk density that ranged from 0.93-1.10 g cm-3. The soil samples also showed substantial variation in water retention and aeration capacity. Both CF and CP exhibited ~41 vol% soil moisture at field capacity (pF 2) but CP held greatly higher water at wilting point (pF 4.2) than CF. Similarly, Water Field Pore Space (WFPS) i.e., water held pore space relative to total pore volume of the soil of CF was lower than CP. On the other hand, air content at pF 2 tended to be lower with CP (14.2%) than CF (23.3%) (Table 1).
In situ soil moisture content: Soil moisture and rainfall varied over sampling period in similar patterns with both land uses (Fig. 3). The analyses of variance showed significant differences in soil moisture contents among sampling dates (p<0.05), between land uses (p<0.05) and sampling datexland use interaction (p<0.05). Soil moisture in December/January was very low (~10%) in both land uses but subsequently increased as precipitation increased to a maximum in June-August (Fig. 3).
| Table 1: | Selected characteristics of coffee (CF) and crop land (CP) soils |
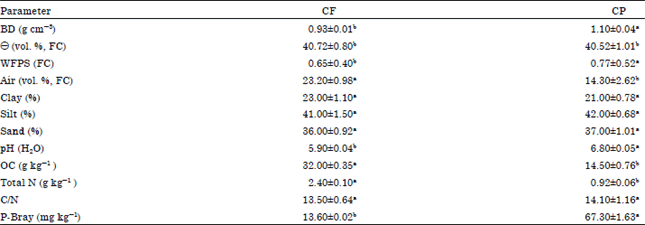 | |
| BD: Bulk density, FC: Field capacity, | |
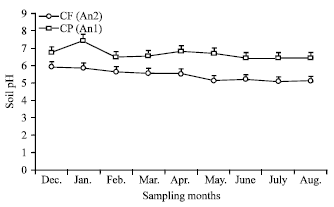 | |
| Fig. 2: | Soil pH determined during in situ nitrogen mineralization from coffee (CF) and crop (CP) land uses of southern Ethiopia |
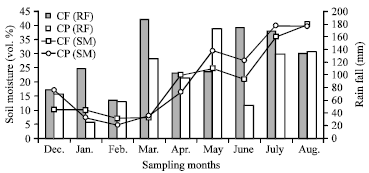 | |
| Fig. 3: | Monthly rain fall; RF (mm) and soil moisture; SM (vol. %) of coffee (CF) and crop (CP) land agroecosystems in southern Ethiopia. The rainfall was determined during study year and soil moisture was taken from 0-15 cm soil depth |
Significant differences were also recorded in in situ soil moisture contents among sampling dates (months) within the same land use. The moisture contents recorded at the field condition with both CF and CP were lower than that recorded in the laboratory at field capacity, pF 2 (Table 1, Fig. 3).
Extractable NH4+-N, NO3¯-N and mineral N content: Extractable NH4+-N was greater at drier condition than wetter condition and in coffee (CF) than in crop (CP) land use during laboratory incubation. There was also difference in extractable NH4+-N between CF and CP during in situ sampling (Table 2). Extractable NO3¯-N differed over time (p<0.05), between land uses (p<0.05) and sampling datexland use interaction (p<0.05) during in situ sampling. Likewise, mineral N content was differed over time (p<0.05), between land uses (p<0.05) and sampling datesxland use interaction (p<0.05). The mineral N content tended to be low when soil moisture content was low and vice versa, both under laboratory incubation and in situ conditions but the degree of response varied between land uses and incubation methods (Table 2). Overall, the mineral N content tended to be greater under in situ condition than with laboratory incubation. The average in situ mineral N content of CP was superior over CF (Table 2). The NO3¯-N concentration was the predominant form of the mineral N with both soils. The NH4+-N content was significantly increased as drought stress progressed in contrary to NO3¯-N mainly under laboratory incubation. In general, the drought stress effect on mineral N content appeared to be greater under laboratory incubation as compared to in situ condition may be because of the soil moisture levels as treatment can be effectively regulated (controlled) in laboratory (Table 2).
Nitrogen mineralization and nitrification rates: The response of N mineralization and nitrification rates to soil moisture contents were estimated by linear regression for both in situ sampling and laboratory incubations. The rates of N mineralization and nitrification were linearly and significantly increased in response to in situ soil moisture with CF. These rates were 9.0 mg kg-1 (R2 = 0.64) for N mineralization and 5.5 mg kg-1 (R2 = 0.53) for nitrification with CF soil during in situ sampling. However, lower rates of N mineralization, 3.8 mg kg-1 (R2 = 0.38) and nitrification 2.4 mg kg-1 (R2 = 0.33) were recorded with CP in response to in situ soil moisture (Fig. 4).
Likewise, under laboratory condition, the rates of N mineralization and nitrification significantly and linearly increased in response to soil moisture increase from air dry to 100% FC (Fig. 5). The degree of responses, however, varied between CF and CP land uses. The rate of N mineralization was 17.3 mg kg-1 with CF (R2 = 0.96) and 4.6 mg kg-1 with CP (R2 = 0.89) in response to soil moisture increase over 45 days incubation.
| Table 2: | Extracted NH4+-N, NO3¯-N and estimated mineral N under in situ versus laboratory condition with two tropical Andosol from southern Ethiopia |
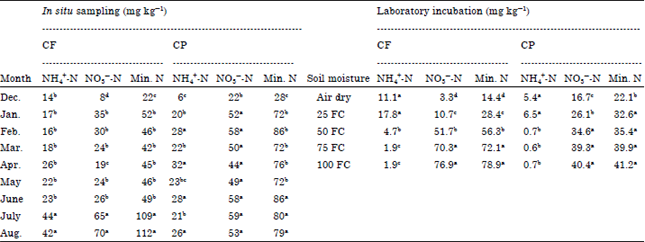 | |
| FC: Field capacity, Min. N: Mineral N. Mean values followed by different letters in a column show statistically significant (otherwise, non-significant difference) | |
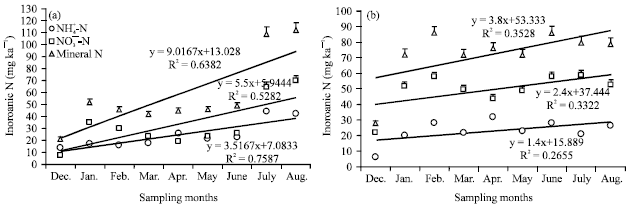 | |
| Fig. 4(a-b): | Relationships between soil moisture and in situ determined NH4+-N and NO3¯-N and calculated mineral N concentration of (a) Coffee; CF (An2) and (b) Crop; CP (An1) land use soils of southern Ethiopia at 0-15 cm soil depth. The soil moisture was determined in 30 days interval, concurrently in situ NH4+-N and NO3¯-N concentrations were determined in 30 days interval with unconfined soil samples |
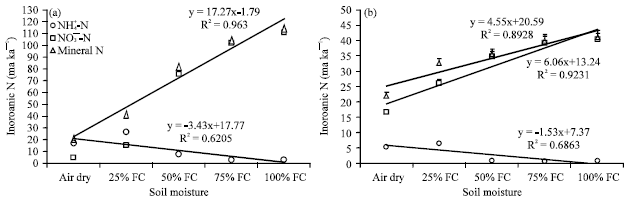 | |
| Fig. 5(a-b): | Relationships between soil moisture and measured NH4+-N and NO3¯-N and estimated mineral N concentration of (a) Coffee; CF (An2) and (b) Crop; CP (An1) land use soils from southern Ethiopia based on laboratory incubation. Soil moisture was initially set at pF 2 (100% FC) and then other moisture levels (treatments; air dry, 25% FC, 50% FC, 75% FC and 100% FC) were developed by proportionally mixing wet and air dry soils, FC: Field capacity |
The rate of nitrification also showed nearly similar trends (Fig. 5). There was remarkably stronger N mineralization in response to soil moisture increase in laboratory incubation than in situ condition.
DISCUSSION
Soil characteristics: The result depicted that coffee (CF) land use, a site with 34 years of coffee plantation history had more SOC than crop (CP) land use of 50 years. The SOC revealed an inverse relationship with bulk density (Table 1). Management practices and vegetation cover differences of land use attributes directly and indirectly affect the level of crop/plant productivity and decomposition of soil organic matter (Ogle et al., 2005). Further more, though we did not exactly know the types of mineral with CF and CP soils, Andosol with rich allophone mineral, aluminum and iron oxide contents were reported to contain greater organic carbon (Jonathan, 2006).
In accordance to our findings, several reports indicated that Andosol and forest soil characterized by high SOC and low bulk density but depended on land use and management practices adhered (Mulugeta, 2004; Ambachew, 2009). Apparently greater SOC and total N content of CF soil could be ascribed to the litter fall and root exudates from permanent vegetations (coffee and shade trees). Similar studies showed that land use had a significant effect on SOC content of soils and its reduction was noted when forest soil was converted to farmlands (Mulugeta, 2004; Lemma and Olsson, 2006; Shrestha et al., 2007). Similar to SOC, the CF land use had significantly (p<0.05) greater total N concentration than CP (Table 2). The most likely explanation for varied SOC and total N concentration between the land uses could be attributed to the variation in intensity of cultivation and amount of residue return to soils. The pH of the studied soils invariably declined from December to August sampling period (Fig. 2) due to organic acid increase as SOM decomposed. The other reasons for pH decrease during SOM decomposition could be strong nitrification, causing release of H+ (Haynes and Mokolobate, 2001).
Nitrification in response to soil moisture: Variation in ammonia oxidation was observed between CF and CP soils both under in situ and laboratory conditions that can be due to the differences in soil characteristics and also due to nitrifiers biomass and activity differences between land uses (Vervaet et al., 2002). Nitrification linearly increased as soil moisture tends to increase with CF and CP during in situ sampling (Fig. 3). Similarly, during laboratory incubation nitrification was linearly increased as soil moisture increased with both CF and CP (Fig. 4). The magnitude of NO3¯-N concentration with varying soil moisture was greater during laboratory incubation. We have calculated a critical value for both nitrification and N mineralization as affected by soil moisture. Then a nitrification rate below 50% of the maximum rate (relative to 100% FC) was considered as critical value (data not shown). Accordingly, the critical value as a response to soil moisture appeared to vary between the land uses. The critical value for CF was 25% FC, i.e., if the soil moisture become lower than 25% FC its nitrification rate would be reduced greatly (by more than 50%). In congruent to our findings, reports indicated that soil moisture appeared to have a regulator effect on heterotrophic processes, once it is lower than a critical value (Grunzweig et al., 2003). It is instructive to note that as soils dry ammonia oxidation rates declines due to constraints on diffusive transport of ammonium, ultimately reducing the activity due to physiological limitations imposed by water stress (Stark and Firestone, 1995; Wang et al., 2006).
The average of both in situ and laboratory incubation extractable NO3¯-N content varied from 64 to 85% of the total inorganic N pool with the studied soils, indicating that NO3¯-N was the predominant form of mineral N pool. This suggested the presence of active and large biomass of nitrifying communities and good soil environment for rapid ammonia oxidation with soils of most near neutral tropical Andosol. However, higher NO3¯-N availability in soils may not be good unless it is synchronized with plant uptake, since it can easily be leached or denitrified. Several other findings have also reported that NO3¯-N is generally the dominant form of plant-available soil N pool that is released during decomposition in most of terrestrial ecosystems, except in wetland and boreal forests which are dominated by NH4+-N (Liang and MacKenzie, 1994; Kaboneka et al., 1997). Similar to our findings an increased nitrification rate, proportional to the N mineralization rate increase was reported by Verchot et al. (2001). The extractable NO3¯-N recorded during in situ sampling of CP could be partly a function of fertilizer N carryover. Besides, the soils release a certain amount of N through the mineralization of soil organic matter and crop residues. This could also be another plausible reason for relatively greater NO3¯-N in CP than in CF during in situ. However, environmental conditions may play substantial role in determining the amounts of N to be mineralized each year.
Nitrogen mineralization rate as affected by soil moisture: Nitrogen mineralization rate was linearly increased as soil moisture tended to increase with CF and CP during in situ sampling (Fig. 4). Similarly during laboratory incubation, rate of N mineralization was linearly increased as soil moisture increased (Fig. 5). The results on N mineralization based on laboratory and in situ methods showed consistent trends in response to soil moisture, with both CF and CP (Fig. 4, 5). The N mineralization of CP tended to be greater under in situ than laboratory incubation, due to the possibly presence of conducive microclimate such as temperature, soil moisture and substrates. By contrast, the in situ N mineralization of CF soil was remarkably lower than laboratory incubation, suggesting an intense competition between roots of perennial plants (coffee and shade trees) and soil microorganisms for nutrients. Likewise, low mineral N during the wet season due to heavy nutrient demand of vigorously growing plants (Singh et al., 1989) and lower decomposition of litter in the presence of trees or under shaded conditions was reported attributed to competition for nutrients between plant roots and decomposer communities (Moorhead et al., 1998). In addition, differences in soil microclimatic conditions among agroecosystems, could affect the microbial biomass and activity, thereby the rates of N mineralization (Jha et al., 1996), consequently, the overall N cycle is affected (Paul and Clarke, 1996). The lower soil P test of coffee than crop land use (Table 2) might also be another contributing factor for low in situ N mineralization and nitrification rates observed. Similar to the present findings substantial variations in the rates of N mineralization and nitrification between pasture and forest soils was reported (Neill et al., 1995).
During laboratory incubation, the NO3¯-N content was almost equivalent with N mineralization with both CF and CP, suggesting that NH4+-N substrate limitation and/or the nitrifiers could be in more active state. Similarly, during in situ condition the large differences between NO3¯-N and inorganic N (NH4+-N + NO3¯-N) was noted with CF due to low nitrifiers activity as the soil was slightly acidic in pH (Abera et al., 2012). The relatively low NO3¯-N contents of CF might be beneficial from an environmental perspective in that there would be low leaching and denitrification losses. The present laboratory incubation results demonstrated that nitrification was much more affected by water stress and diffusion limitation than ammonification. The study confirms the differential sensitivity to water availability of the microorganisms responsible for the two soil N transformation processes, ammonification and nitrification (Yahdjian and Sala, 2008; Abera et al., 2012).
Methods of N mineralization assessment and its implication: Many reports indicated that in situ measurement of N mineralization by sequential incubation of soil cores has many advantages compared with laboratory measurements (Raison et al., 1987; Adams et al., 1989). The argument was that soil remains relatively undisturbed and the mineralization process occurs under ambient temperatures and, to some degree with ambient moisture conditions (fewer disturbances to natural processes than others). The moisture content in the cores remains close to the level prevailing at the start of the incubation (Raison et al., 1987). This is a limitation of the method, because in the unconfined incubation the moisture content generally fluctuates than in the confined cores. In the present study, however, we used unconfined in situ sampling and measurement of N mineralization by periodic sampling of soils and immediately measuring NH4+-N and NO3¯-N contents in laboratory condition. Great care was also taken to minimize moisture loss during sampling, transportation and determination in the laboratory condition. Hence it may be even more efficient than in situ incubation of core soils as the moisture and microbial movements were not restricted as in the core incubation technique. However, the method we use may have some limitation since the measurement of NH4+-N and NO3¯-N was made after transporting the soils to laboratory which we suppose might have created some disturbance with regards to soil moisture and microbial activity. Furthermore, in situ mineralized N could be prone to leaching and denitrification loss and could also be immediately taken up by growing plants/crops. These factors could under estimate the potential N dynamics. Nevertheless, the in situ N mineralization determination has significant ecological and agronomic implication than laboratory and sequential core incubation methods.
In general, in situ N mineralization showed a substantial release of N across seasons in both CF and CP, so that this knowledge can help in estimating the amount of soil organic N pool that can be mineralized over time to sustain crop productivity in low input tropical agriculture. In nutshell, the results of both methods enhanced our understanding of the extent of N mineralization from SOM in order to decide the rational amount of N to be supplied through fertilizer sources. This knowledge could help in optimizing the N use efficiency; thereby reduce the N fertilizer cost and N losses that may lead to environmental hazards. Such types of unconfined in situ sampling studies on N mineralization and nitrification are scarce under tropical conditions but it has potential links to ecosystem N losses, either through gaseous emissions or nitrate leaching.
CONCLUSION
Land use management practices and soil moisture significantly affected decomposition of SOM, total N and soil fertility. Coffee (CF) land use exhibited higher SOC and total N than crop (CP) land use. As a result, CF soil displayed greater N mineralization at optimum soil moisture conditions than CP. The results of laboratory incubation revealed that extractable NH4+-N declined, while NO3¯-N and mineral N were increased in response to soil moisture increase implying that the ammonifiers and nitrifiers communities varied in sensitivity to soil moisture stress.
The remarkable N mineralization and nitrification reduction of both CF and CP soils due to drought stress under laboratory incubation indicates the seasonality of N mineralization in tropical soils. Therefore, the knowledge of the effect of soil moisture stress on N mineralization dynamics of tropical soils could be important from economic and environmental perspectives with regards to deciding optimum N fertilizer supply and for model based prediction of the rates of N mineralization. The present laboratory and in situ studies suggested that the use of different N mineralization assessment methods could be important to provide reliable results. For instance, the N mineralization dynamics linearly responded to soil moisture in laboratory incubation, suggesting the striking effect of soil moisture when other factors kept controlled. However, under in situ sampling, albeit there was good positive relationship between N mineralization dynamics and soil moisture but was not strongly linear like in laboratory incubation. It is essential to note that the in situ N mineralization determination would have more ecological and agronomic implications as it indicated that N mineralization is affected not only by soil moisture but also by many other factors such as changes in soil microbial communities, temperature, litter input and SOM.
ACKNOWLEDGMENTS
We gratefully acknowledge the Norwegian Programme for Development, Research and Higher Education (NUFU) for funding both the field and laboratory research work that allowed us to present the results here as scientific article. Help rendered by different groups of people during the process of data generation is duly acknowledged. We also acknowledge the anonymous reviewers for their constructive suggestions and comments on the early draft of this manuscript.
REFERENCES
- Abera, G., E. Wolde-meskel and L.R. Bakken, 2012. Carbon and nitrogen mineralization dynamics in different soils of the tropics amended with legume residues and contrasting soil moisture contents. Biol. Fertil. Soils, 48: 51-66.
CrossRef - Adams, M.A., P.J. Polglase, P.M. Attiwill and C.J. Weston, 1989. In situ studies of nitrogen mineralization and uptake in forest soils, some comments on methodology. Soil Biol. Biochem., 21: 423-429.
CrossRef - Ambachew, D., 2009. Effects of conversion of Natural forest to plantations, traditional agroforestry and cultivated lands on carbon sequestration and maintenance of soil quality in Gambo district, southern Ethiopia. Ph.D. Thesis, Department of Plant and Environmental Sciences, Norwegian University of Life Sciences, As, Norway.
- De Neve, S., R. Harmann and G. Hofman, 2003. Temperature effects on N-mineralization: Changes in soil solution composition and determination of temperature coefficients by TDR. Eur. J. Soil Sci., 54: 49-61.
CrossRef - Feller, C. and M.H. Beare, 1997. Physical control of soil organic matter dynamics in the tropics. Geoderma, 79: 69-116.
CrossRefDirect Link - Goncalves, J.L. and J.C. Carlyle, 1994. Modelling the influence of moisture and temperature on net nitrogen mineralization in a forested sandy soil. Soil Biol. Biochem., 26: 1557-1564.
CrossRefDirect Link - Grunzweig, J.M., S.D. Stephen and F.S. Chapin, 2003. Impact of forest conversion to agriculture on carbon and nitrogen mineralization in subarctic Alaska. Biogeochemistry, 64: 271-296.
Direct Link - Jha, P.B., A.K. Kashyap and J.S. Singh, 1996. Effect of fertilizer and organic matter inputs on nitrifier populations and N-mineralization rates in dry tropical region, India. Applied Soil Ecol., 4: 231-241.
Direct Link - Kaboneka, S., W.E. Sabbe and A. Mauromoustakos, 1997. Carbon decomposition kinetics and nitrogen mineralization from corn, soybean and wheat residues. Commun. Soil Sci. Plant Anal., 28: 1359-1373.
CrossRefDirect Link - Kushwaha, C.P., S.K. Tripathi and K.P. Singh, 2000. Variations in soil microbial biomass and N availability due to residue and tillage management in a dry land rice agroecosystems. Soil Till. Res., 56: 153-166.
CrossRef - Michelsen, A., M. Anderson, M. Jensen, A. Kjoller and M. Gashew, 2004. Carbon stocks, soil respiration and microbial biomass in fire-prone tropical grassland, woodland and forest ecosystems. Soil Biol. Biochem., 36: 1707-1717.
CrossRefDirect Link - Moorhead, D.L, M.M. Westerfield and J.C. Zak, 1998. Plants retard litter decay in a nutrient-limited soil: A case of exploitative competition?. Oecologia, 113: 530-536.
CrossRef - Neill, C., M.C. Piccolo, P.A. Steudler, J.M. Melillo, B.J. Feigl and C.C. Cerri, 1995. Nitrogen dynamics in soils of forests and active pastures in the western Brazilian Amazon Basin. Soil Biol. Biochem., 27: 1167-1175.
Direct Link - Nelson, D.W. and L.E. Sommers, 1983. Total Carbon, Organic Carbon and Organic Matter. In: Methods of Soil Analysis, Part 2: Chemical and Microbiological Properties Page, A.L., R.H. Miller and D.R. Keeney (Eds.)., 2nd Edn., ASA and SSSA, Madison, WI., USA, pp: 539-579.
CrossRefDirect Link - Owena, J.S., M.K. Wangb, C.H. Wanga, H.B. Kingc, H.L. Sunb, 2003. Net N mineralization and nitrification rates in a forest ecosystem in northeastern Taiwan. For. Ecol. Manage., 176: 519-530.
CrossRef - Post, W.M. and K.C. Kwon, 2000. Soil carbon sequestration and land-use change: Processes and potential. Glob. Change Biol., 6: 317-327.
Direct Link - Raison, R.J., M.J. Connell and P.K. Khanna, 1987. Methodology for studying fluxes of soil mineral-N in situ. Soil Biol. Biochem., 19: 521-530.
CrossRef - Shrestha, B.M., B.R. Singh, B.K. Sitaula, R. Lal and R.M. Bajracharya, 2007. Soil aggregate-and particle-associated organic carbon under different land uses in Nepal. Soil Sci. Soc. Am. J., 71: 1194-1203.
CrossRefDirect Link - Singh, J.S., A.S. Raghubanshi, R.S. Singh and S.C. Srivastava, 1989. Microbial biomass acts as a source of plant nutrients in dry tropical forest and savanna. Nature, 338: 449-500.
CrossRefDirect Link - Stark, J.M. and M.K. Firestone, 1995. Mechanisms for soil moisture effects on activity of nitrifying bacteria. Applied Environ. Microbiol., 61: 218-221.
Direct Link - Sugihara, S., S. Funakawa, M. Kilasara and T. Kosaki, 2010. Effect of land management and soil texture on seasonal variations in soil microbial biomass in dry tropical agroecosystems in Tanzania. Applied Soil Ecol., 44: 80-88.
CrossRef - Verchot, L.V., Z. Holmes, L. Mulon, P.M. Groffman and G.M. Lovett, 2001. Gross vs net rates of N mineralization and nitrification as indicators of functional differences between forest types. Soil Biol. Biochem., 33: 1889-1901.
CrossRef - Vervaet, H., P. Boeckx, V. Unamuno, O. Van Cleemput and G. Hofman, 2002. Can 15N profiles in forest soils predict NO3- loss and net N mineralization rates?. Biol. Fertil. Soils, 36: 143-150.
CrossRef - Yahdjian, L. and O.E. Sala, 2008. Do litter decomposition and nitrogen mineralization show the same trend in the response to dry and wet years in the Patagonian Steppe?. J. Arid Environ., 72: 687-695.
CrossRefDirect Link - Wang, C., S. Wan, X. Xing, L. Zhangc and X. Han, 2006. Temperature and soil moisture interactively affected soil net N mineralization in temperate grassland in Northern China. Soil Biol. Biochem., 38: 1101-1110.
CrossRef - Bray, R.H. and L.T. Kurtz, 1945. Determination of total, organic and available forms of phosphorus in soils. Soil Sci., 59: 39-46.
CrossRefDirect Link - Haynes, R.J. and M.S. Mokolobate, 2001. Amelioration of aluminium toxicity and P deficiency in acid soils by addition of organic residue: A critical review of the phenomenon and the mechanism involved. Nutrient Cycling Agroecosyst., 59: 47-63.
CrossRef