
Hashem M. Mohamed
Department of Soil and Water, Faculty of Agriculture, University of Assiut, Assiut, 71526, Egypt
Ebaid M.A. Ibrahim
Department of Genetics, Faculty of Agriculture, University of Assiut, Assiut, 71526, Egypt
International Journal of Soil Science
Year: 2011 | Volume: 6 | Issue: 3 | Page No.: 176-187







ABSTRACT
This investigation aimed at modifying the ability of phosphate-solubilizing bacteria, Bacillus polymyxa var. phosphaticum to improve plant growth of tomato plants. Bacillus polymyxa mutants selected after N-methyl-N′-nitro-N-nitrosoguanidin (NTG) treatments were evaluated for their efficiency in improving growth, P-uptake and Fe-uptake by tomato in greenhouse. Only five mutants out 1000 colonies were found to solubilize phosphates more than wild type strain on the basis of the solubilization index on Pikovskyaya's solid medium. Phosphate solubilization index of the five mutants M1, M2, M3, M4, M5 were 2.56, 2.42, 2.28, 2.49 and 2.31, respectively. The highest phosphate solubilization index was recorded by mutant strain M1 compared with the wild type strain (2.13). The results show that all tested strains produced; in their cultures; organic substances that chelated iron in the calcareous soil. In greenhouse experiment the results showed that inoculation of tomato cultivar Super marmand with wild strain or mutant M1 with or without the addition of rock phosphate had a significant effect on shoot and root dry weights, p-uptake and Fe-uptake in shoot compared with uninoculated treatment (control) or amended with rock phosphate alone. Similar results were recorded with Burchard cultivar but in general, the cultivar Super marmand was more responsive to wild type and mutant M1 inoculation than the cultivar Burchard. Application of both mutant and rock phosphate resulted in the highest availability of P and Fe in the soil, resulted in increases about 26% in P and 36% in Fe as compared to control.
PDF Abstract XML References Citation
Received: December 25, 2010;
Accepted: February 23, 2011;
Published: March 25, 2011
How to cite this article
Hashem M. Mohamed and Ebaid M.A. Ibrahim, 2011. Effect of Inoculation with Bacillus polymyxa Mutants on Growth, Phosphorous and Iron Uptake by Tomato (Lycopersicon esculentum L.) in Calcareous Soils. International Journal of Soil Science, 6: 176-187.
DOI: 10.3923/ijss.2011.176.187
URL: https://scialert.net/abstract/?doi=ijss.2011.176.187
DOI: 10.3923/ijss.2011.176.187
URL: https://scialert.net/abstract/?doi=ijss.2011.176.187
INTRODUCTION
Phosphorus and iron are essential elements for plants but the concentration of total P in soils ranges from 100 and 3000 mg P kg"1 soil (Frossard et al., 2000), the variation being largely due to differences in weathering intensity and parent material composition (Raji, 2006; Bilgili et al., 2008). To increase the availability of phosphorus for plants, large amounts of fertilizers are used on a regular basis but after application, a large proportion of fertilizer phosphorus is quickly transferred to insoluble form (Omar, 1998). Arpana et al. (2002) reported that a great proportion of phosphorus in chemical fertilizer becomes unavailable to the plants after its application to the soil. They referred this to formation of strong bonds of phosphorus with calcium and magnesium in alkaline pH and the same bonds with iron and aluminum in acidic soils. In calcareous soils phosphorous fertilizers are fixed by calcium carbonate through adsorption and precipitation, so their efficiency is not more than 20% (Tisdale et al., 1993). Also, the total Fe content of soils is much higher than plants require, its bioavailability is limited in most soils, particularly in calcareous soils (Guerinot and Yi, 1994; Lopez-Bucio et al., 2000). As a result, Fe deficiency chlorosis is evident in about 30% of crops worldwide (Imsande, 1998).
The ability of soil microorganisms to solubilize phosphates is considered to be one of the most important traits associated with plant P nutrition (Richardson, 2001; Mishra et al., 2010). Phosphorus biofertilizers in the form of microorganisms especially phosphate- solubilizing bacteria in rhizosphere can help in increasing the availability of accumulated phosphates for plant growth by solubilization (Nautiyal et al., 2000). Phosphate Solubilizing Microorganisms (PSMs) play an important role in supplementing phosphorus to the plants, allowing a sustainable use of phosphate fertilizers. Microorganisms are involved in a range of process that affect the transformation of soil Phosphorus (P) and thus are integral component of the soil ‘P’ cycle. Application of PSMs to the field has been reported to increase crop yield. Several mechanisms like lowering of soil pH by acid production, ion chelation and exchange reactions in the growth environment have been reported to play a role in phosphate solubilization by PSMs (Whitelaw, 2000; Gyaneshwar et al., 2002; Pradhan and Sukla, 2006; Mostafa and Abo-Baker, 2010).
Also, many microorganisms are crucial partner of plants to access Fe in the rhizosphere, they can improve plant growth through a continuous and efficient Fe supply for growth and chlorophyll synthesis (Masalha et al., 2000; Rroco et al., 2003). Different mechanisms have been suggested by which microbial species enhance Fe availability to plants. For instance, some bacterial species produce and secrete bacterial siderophores that mobilize Fe from soil to roots (Neilands, 1995). In addition, certain bacterial species isolated from the rhizosphere of bean and maize plants contribute significantly to Fe uptake by plants likely through increased Fe (III) reduction in the rhizosphere (Valencia-Cantero et al., 2007).
The aim of this study was to modify of the ability of phosphate-solubilizing bacteria, Bacillus polymyxa var. phosphaticum to improve plant growth by classical genetic methods of mutants with higher phosphate-solubilization activity. The efficiency of the wild and selected mutants to promote plant growth, P-uptake and Fe-uptake was determined by using different cultivars of tomato (Lycopersicon esculentum) as test crop in a pot experiment.
MATERIALS AND METHODS
Bacterial strain: Bacillus polymyxa strain was obtained from Desert Research Center, Egypt which used in large-scale production of biofertilizer called Phosphorien. The strain was maintained on Pikovskyaya's agar slants medium modified by Rao and Sinha (1963) having following composition: Glucose, 10 g; Ca3 (PO4)2 , 5 g; (NH4)2SO4, 0.5 g; KCl, 0.2 g; MgSO4.7H2O, 0.1 g; MnSO4 trace; FeSO4 trace; 20 g agar, at 4°C in refrigerator.
Development of mutants: The wild type strain Bacillus polymyxa mutated with N-methyl-N’-nitro-N-nitrosoguanidin (NTG) as described by Mishra and Goel (1999). In order to estimate the optimal NTG concentration that would lead to a 50% killing rate, a killing curve was performed by exposure of Bacillus polymyxa cell suspension for different time intervals to different concentrations of NTG ranging from 25-300 μg mL-1 (dissolved in phosphate buffer, pH 7.0). Exposure of a 24 h old cell suspension of Bacillus polymyxa to 50 μg mL-1 of NTG for 90 min at 28°C in shacked water bath resulted in a 50% killing rate. This concentration was used to induce mutations. After exposure, the cells were centrifuged, washed 2 times with phosphate buffer and suspended in 2 mL sterile saline solution (0.9 M NaCl). Overnight-grown culture of Bacillus polymyxa was inoculated in Pikovskyaya's liquid medium and was subjected to NTG treatment for mutagenesis. After NTG treatment, the cell suspension (0.1 mL) was spread onto solid Pikovskyaya's (PVK) medium (pH 7.0) and incubated at 28°C along with untreated control for 48 h. Afterwards, the colonies were observed on the media and were collected for further analysis. All of the possible mutants were screened for solubilizing index (Edi-Premono et al., 1996) (compared to parent strains) after 48 h growth in solid PVK medium (pH 7.0) at 28°C. Mutants showing maximum solubilizing index were selected for further studies.
Determination of solubilization index: The ability of the wild type and selected mutants to dissolve insoluble phosphate was described by the solubilization index: the ratio of the total diameter (colony+halo zone) and the colony diameter (Edi-Premono et al., 1996). Halo zone formation around strains growth was measured after inoculated on Pikovskyaya's agar medium for 5 days at 28°C.
Determination of pH and soluble phosphates: The ability of wild type and selected mutants to dissolve insoluble tricalcium phosphate was measured in 100 mL aliquots of modified Pikovskyaya's liquid medium. The wild and mutants strains were grown in 100 mL aliquots of the liquid medium for 5 days at 28°C, then the cultures were filtered and centrifuged at 10000 g for 10 min. Soluble phosphates were assayed by chlorostannus reduced molybdo-phosphoric acid blue method (Allen, 1974) and pH of the supernatant was measured by a pH meter.
Production of organic chelating agents: The ability of wild type and selected mutants strains grown on PVK broth medium to produce organic substances and chelating agents capable of complexing and extracting polyvalent mineral ions from soil was tested. Each strain was grown, for 5 days at 28°C, in 100 mL aliquots of the medium in a 200 mL Erlenmeyer flask. The cultures were then filtered and adjusted to pH 7.0 before use for Fe extraction. Suspensions of a calcareous soil (CaCO3 161.8 g kg-1) with 0.005M EDTA (ethylene di-amine tetra aceticacid), 0.005M DTPA (di-ethylene tri-amine penta-aceticacid) and each of the wild type and mutants strains culture (109 CFU mL-1) (1:2) were prepared and shacked for 30 min, then filtered. Amounts of available Fe extracted from the soil by each of the culture filtrates in comparison with those extracted by EDTA and DTPA extractants were then determined using an atomic adsorption spectrometer model (GBC 109 AA).
Greenhouse experiment: The trial was carried out in the Greenhouse of Soil and Water Department, Faculty of Agriculture Assiut University. A pot experiment was conducted in 2008 season to study the effect of wild strain and selected mutant M1 on plant growth, P uptake and Fe-uptake by tomato plants in calcareous soil collected from El-Gorahib Experimental Farm of Assiut University. The physical and chemical properties of soil are presented in Table 1. The experiment was established with 6 treatments: without rock P material or bacterial inoculation (control), rock P material 0.3%, inoculation with wild strain, inoculation with selected mutant M1, rock P+wild strain, rock P+selected mutant M1. Tomato seed varieties Burchard and Super marmand obtained from the Ministry of Agriculture, Egypt were sown in trays (30x50 cm, 10 cm deep) containing sieved clay soil mixed with 3% peat moss and watered twice a week. After 45 days, similar healthy seedlings (15 cm in length) were uprooted, inoculated or un-inoculated with separate culture of the strain used before transplanting in black pots, 30 cm in diameter containing 5 kg sieved clay soil collected from Assiut Experimental Farm.
| Table 1: | Soil physical and chemical characteristics |
 | |
The wild type and mutants strains were grown each on 100 mL aliquot of nutrient broth (NB) medium and incubated at 28°C for 5 days. The cultures contained about 109 CFU mL-1 determined on plates of (NB) agar medium. Dipping the root of seedlings in broth culture wild strain or mutants for one hour made inoculation. Two seedlings were transplanted in each pot and 5 replicates were planted for each particular treatment. Plants were irrigated when needed and fertilized as usual (N-fertilizer, in urea form, at a rate of 215 kg N ha-1). After 40 days from transplanting, plants were uprooted, washed and used for determination of dry weight of shoot and roots. Dried shoots were ground and submitted to the acid-digestion using a 2:1 HNO3: HClO4 acid mixture for the determination of P and Fe. To analyze mineral elements, soils were sampled for chemical analyses before the experiment and after harvesting the plants. At harvest, following root removal, soils in the whole pot were mixed dried and sieved (2 mm screen) and analyzed for available P and Fe.
Statistical analysis: Data obtained were subjected to statistical analysis by using the statistical computer program (StatSoft, 1995). Differences among treatments were tested by ANOVA and mean values among treatments were compared by Duncan's Multiple Range test at p = 0.05.
RESULTS AND DISCUSSION
After subjecting B. polymyxa to mutagensis with NTG, only five mutants from out 1000 colonies were found to solubilize phosphates more than wild type strain on the biases of the solubilization index on Pikovskyaya's solid medium (Fig. 1). The highest phosphates solubilization index was recorded by mutant strain M1 (2.56) compared with the wild type strain (2.13). The strains were further studied for their ability to solubilize insoluble phosphates in Pikovskyaya's liquid medium containing tricalcium phosphate. Amounts of phosphorus solubilized from tricalcium phosphate by the wild type and mutant strains are shown in Fig. 2. The selected mutant strains of Bacillus polymyxa significantly solubilized more phosphates than the wild type strain.
The maximum amount of P solubilized (2.68 μg mL-1) was recorded for mutant M1 which also recorded the highest phosphate solubilization index, 2.56. The result also show that the lowest amount of phosphates solubilized was recorded by mutant M3 (2.41 μg mL-1). Figure 3 shows the pH values of the strain cultures after 5-days growth on the Pikovskyaya's liquid medium (adjusted to pH 7.00).
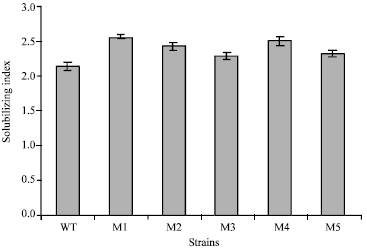 | |
| Fig. 1: | Solubilization index of the wild type (WT) strain and mutants (M1, M2, M3, M4 and M5) grown on Pikovskyaya's solid medium for 5 days |
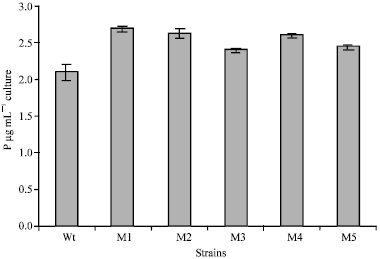 | |
| Fig. 2: | Released-P by the wild type (WT) strain and mutants (M1, M2, M3, M4 and M5) during 5 days of incubation on Pikovskyaya's broth medium |
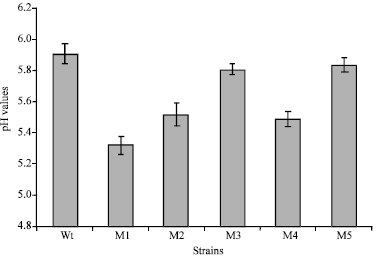 | |
| Fig. 3: | Final pH values of the wild type (WT) strain and mutants (M1, M2, M3, M4 and M5) cultures after 5 days incubation on Pikovskyaya's medium adjusted to pH 7.0 |
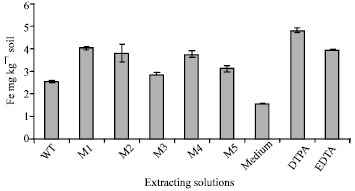 | |
| Fig. 4: | Amounts of extracted Fe by the wild type (WT) strain and mutants (M1, M2, M3, M4 and M5) cultures compared with DTPA, EDTA and Pikovskyaya's medium |
As indicated from the results obtained, the pH of the broth was found to decline with all strains and the highest acidity was produced by the mutant M1 (pH 5.32). The solubilized amounts of Phosphates and the determined phosphate solubilization index for the strains are compatible with the acidity produced in their cultures. This indicates that the organic acids produced during the metabolism of glucose are the main cause of solubilization (Hwangbo et al., 2003). Solubilization of insoluble phosphates by microorganisms is mainly by production of organic acids and chelation substances (Singh et al., 1980; Luo et al., 1993). The phosphate solubilization ability of wild and mutant strains of Pseudomonas fluorescens had been reported to be accompanied by decrease in pH of the medium (Katiyar and Goel, 2003). Two mutants of Pseudomonas corugata were found more efficient than wild type strain for phosphate solubilization activity across a range of temperature from psychotropic (4°C) to mesophilic (28°C) in aerated GRC medium containing insoluble rock phosphate (Trivedi and Sa, 2008). Asad-ur-Rehman et al. (2003) reported that the Aspergillus niger mutant UV-M9 was produced by giving UV dose to pre-grown mycelia for enhanced citric acid production.
The ability of wild type and mutant strains to produce organic substances in their cultures which can chelate iron (or other polyvalent elemental ions) from soil was tested in a laboratory experiment. Figure 4 shows the amount of Fe (ppm) extracted from calcareous soil by filtrate of the wild type and mutant strains compared with those extracted by nutrient broth medium, DTPA and EDTA. The results show that all tested strains produced in their cultures organic substances that chelated iron in the soil. The extracted amounts of iron by the mutant strains cultures were 4.08, 3.84, 2.92, 3.82, 3.15 and 2.63 mg kg-1 soil for the mutant M1, M2, M3, M4, M5 and wild type strain, as compared with 1.60, 4.90 and 4.03 mg kg-1 soil, respectively extracted by PVK broth medium, DTPA and EDTA.
These results show the difference in amounts of extracted Fe by mutant strain cultures, being highest by mutant M1 and lowest by mutant M3 culture. The data also show that the iron extracted by M1 culture (4.08 mg) was more than that extracted by EDTA (4.03 mg) but less than that extracted by DTPA (5.12 mg). It is also noticed that mutant M1 produced the highest acidity of all tested mutant (pH 5.19), as shown in Fig. 3.
Nevertheless, the extracted amounts of Fe by wild type and mutant cultures are probably produced by organic chelating substances other than acidity, since the pH of the cultures and PVK broth medium, as well as the extraction solutions EDTA and DTPA, was adjusted to pH 7.3 before extraction. It is well known that numerous organic substances that are formed in soils, such as folvic and humic acids, have complexing ability, largely due to content of oxygen-containing functional groups such as COOH, phenolic OH and C = O groups of various types. Amino and imino groups may also be involved (Stevenson, 1982). Moreover, evidences are presented by many investigators, about the formation of organic substances, in microbial cultures, having metal complex capability, like vitamin, nucleic acids, amino acids and hormones, in addition to the presence of organic acids (Stevenson, 1982).
Berrow et al. (1982) studies the trace element levels extractable by 2-Ketogluconate (2-KG), ammonium acetate, acetic acid, EDTA and DTPA in three different soils. They concluded that 2-Ketogluconate obtained from a batch culture of an Erwina species, was able to extract more cobalt, nickel, zinc, iron, titanium and vanadium than ammonium acetate and was equal to EDTA and DTPA in extracting copper, manganes, molybdenum, nickel and zinc. The capability of 2-Ketogluconate to extract more Co, Ni and Zn and considerably more Fe, Ti and V than ammonium acetate at the same pH indicates that it is chelating a portion of the non-exchangeable form of these metals in soil. Microbial isolates forming poly-basic organic acids as tartaric and citric, oxalic, formic and glycolic acids are found more effective in chelating Ca and Fe than other organic acids (Banik and Dey, 1982). Kucey (1988) showed that the addition of 0.05 M EDTA to solutions containing insoluble copper and zinc compounds had the same solubilizing effect as inoculation with Penicillium bilaji . But, reduction of the solution pH to 4.0 by the addition of 0.1 N HCl did not result in metal ion solubilization.
In greenhouse experiment an attempt was tried to study the effect of inoculation with wild type and mutant with or without rock phosphate (0.3%), on growth, phosphorous and iron uptake by tomato (cv. Super marmand and Burchard) and available P and Fe in the soil 40 days following the planting were presented in Table 2. Addition of rock phosphate to the soil non-significantly increased available P or Fe compared with control (without inoculum and without rock phosphate fertilizer). Inoculation with wild type (B. polymyxa) or mutant M1 resulted in significant increase in P and Fe availability in the soil, in which mutant M1 was more efficiently than wild type.
Application of both mutant and rock phosphate resulted in the highest availability of P and Fe in the soil, resulted in increases about 26% in P and 36% in Fe as compared to control. The data on Super marmand inoculation (Table 3) show that the addition of rock phosphate alone caused non-significant increases in shoot and root dry weights and Fe uptake while a significant effect on P uptake. Also data show that inoculation of wild strain or mutant to the soil with or without addition rock phosphate had a significant effect on shoot and root dry weights, P-uptake and Fe uptake in shoot compared with control (without inoculum and without rock phosphate fertilizer), or amended with rock phosphate only. The treatment inoculated with mutant M1 plus rock phosphate was stimulative; inducing the following shoot and root dry weights, P uptake and Fe uptake in shoot; 41.2, 57.5, 26.2 and 34.3% compared with control in percentage increases, respectively.
Data on Burchard cultivar inoculation (Table 4) show that in general, the cultivar Super marmand was more responsive to wild type and mutant inoculation than cultivar Burchard. Similar to Super marmand, inoculation with mutant M1 plus rock phosphate was the most stimulative; inducing the following % increases in shoot and root dry weights, P-uptake and Fe-uptake in shoot; 31.7, 41.1, 25.1 and 28.3% compared with control respectively. In accordance with the results obtained in the present pot experiment Narula et al. (2001) investigated the establishment of phosphate dissolving strains of Azotobacter chroococcum, including soil isolates (wild type) and their mutants, in the rhizosphere and their effect on the growth wheat cultivars (Triticum aestivum L.).
| Table 2: | Effects of inoculation with wild type and selected mutant on available P and Fe (mg kg-1) in soil planted with tomato cultivars |
 | |
| RP = Rock phosphate, WT = Wild type, M1 = Mutant strain No. 1.The values in column followed by the same letter(s) are not significantly different by Duncan's multiple range test at 5% level significant | |
| Table 3: | Effects of inoculation with wild type and selected mutant on dry weight and uptake of P and Fe of tomato cv. Super marmand |
 | |
| RP = Rock phosphate, WT = Wild type, M1 = Mutant strain No. 1. The values in column followed by the same letter(s) are not significantly different by Duncan's multiple range test at 5% level significant | |
| Table 4: | Effects of inoculation with wild type and selected mutant on dry weight and uptake of P and Fe of tomato cv. Burchard |
 | |
| RP = Rock phosphate, WT = Wild type, M1 = Mutant strain No. 1. The values in column followed by the same letter(s) are not significantly different by Duncan's multiple range test at 5% level significant | |
They found that the mutant strains performed better than the soil isolates for in vitro phosphate solubilization (11-14%) and growth hormone production (11.35%). Seed inoculation of wheat varieties with phosphate-solubilizing and phytohormone-producing A. chroococcum showed a better response over the control. Mutant strains of A. chroococcum showed a higher increase in grain (15.30%) and straw (15.10%) yield over the control and better survival (12-14%) in the rhizosphere as compared to their parent soil isolate (P4). Mutant strain M15 performed better in all three varieties in terms of increase in grain yield (20.8%) and root biomass (20.6%) over the control. Gupta et al. (2002) developed of Cd, Ni and Cr-resistant of phosphate solubilizing Pseudomonas sp. NBRF 4014. Mutants were stable under non-selective pressure. However, they were able to promote root and shoot elongation in soybean in the presence of metals unfamiliar to wild type.
Using sterilized soil amended with either soluble or rock phosphate, inoculation with mutants of Pseudomonas corrugate showed greater positive effect on all of the growth parameters of wheat plants compared to the wild strain in greenhouse trials (Trivedi and Sa, 2008). Genetically modified strains of Penicillium rugulosum were able to stimulate growth of maize plants by means of increased P uptake (Reyes et al., 2002). Improving availability of P in soils with bacterial inoculation or with combined inoculation and rock phosphate which may lead to increas P uptake and plant growth was reported by many researchers (Wahid and Mehana, 2000; Sahin et al., 2004; Han et al., 2006). Omar (1998) reported that application of rock phosphate to non-sterilized soil with bacterial inoculation increased P uptake and shoot and total dry mass of wheat plant. A plant growth promoting effect of phosphate solubilizing bacteria like Bacillus may be related to their ability to produce plant growth regulating substances, such as IAA (Sattar and Gaur, 1987; Leinhos and Bergmann, 1995; Sheng and Huang, 2001). Also, the production of other metabolites beneficial to plant, such as antibiotics and siderophores, has created confusion about the specific role of phosphate solubilization in plant growth stimulation (Kloepper et al., 1989). Several researches show that microbial inoculation enhances plant Fe uptake. Masalha et al. (2000) stated that plants (maize and sunflower) cultivated under non-sterile condition grew well, showed no Fe deficiency symptoms and had fairly high Fe concentration in the roots in contrast to plant grown in the sterile medium. It may be therefore assumed that microbial activity in particular their siderophores is of pivotal importance for plant Fe uptake. Soil microbial enhancement of Fe acquisition by red clover by producing siderophores which increase Fe bioavailability and auxins which increase Fe(III) reduction capability (Chong et al., 2006).
CONCLUSION
In general, this result indicates that it is possible to select efficient strain of Bacillus polymyxa through mutagenesis, which may improve availability of P and Fe in soils, plant growth and P and Fe uptake by plants. Inoculation of wild strain or mutant M1 to the soil with or without addition rock phosphate had a significant effect on shoot and root dry weights, p uptake and Fe uptake in shoot of tomato plants compared with control (without inoculum and without rock phosphate fertilizer). Also, the treatment inoculated with mutant M1 plus rock phosphate was the most promotive in all tested growth parameters. In future we should investigate the stability and performance of the phosphate solubilization trait once the bacteria have been inoculated in soil, in both wild type and mutagenesis strains.
ACKNOWLEDGMENT
The authors would like to thank Faculty of Agriculture, Assiut University, Egypt for supporting his study.
REFERENCES
- Banik, S. and B.K. Dey, 1982. Available phosphate content of an alluvial soil as influenced by inoculation of some isolated phosphate-solubilizing micro-organisms. Plant Soil, 69: 353-364.
CrossRefDirect Link - Berrow, M.L., M.S. Davidson and J.C. Burridge, 1982. Trace elements extractable by 2-ketogluconic acid from soil and their relationship to plant contents. Plant Soil, 66: 161-171.
CrossRefDirect Link - Chong, W.J., Y.F. He, C.X. Tang, P. Wu and S.J. Zheng, 2006. Mechanisms of microbially enhanced Fe acquisition in red clover (Trifolium pretense L.). Plant Cell Environ., 29: 888-897.
CrossRefDirect Link - Frossard, E., L.M. Candron, A. Oberson, S. Sinaj and J.C. Fardeau, 2000. Processes governing phosphorus availability in temperate soils. J. Environ. Qaul., 29: 15-23.
Direct Link - Guerinot, M.L. and Y. Yi, 1994. Iron: Nutritious, noxious and not readily available. Plant Physiol., 104: 815-820.
Direct Link - Gupta, A., J.M. Meyer and R. Goel, 2002. Development of heavy metal-resistant mutants of phosphate solubilizing Pseudomonas sp. NBRF 4014 and their characterization. Curr. Microbiol., 45: 323-327.
CrossRefDirect Link - Gyaneshwar, P., G.N. Kumar, L.J. Parekh and P.S. Poole, 2002. Role of soil microorganisms in improving P nutrition of plants. Plant Soil, 245: 83-93.
CrossRefDirect Link - Han, H.S., Supanjani and K.D. Lee, 2006. Effect of co-inoculation with phosphate and potassium solubilizing bacteria on mineral uptáme and growth of pepper and cucumber. Plant Soil Environ., 52: 130-137.
CrossRefDirect Link - Hwangbo, H., R.D. Park, Y.W. Kim, Y.S. Rim and K.H. Park et al., 2003. 2-Ketogluconic acid production and phosphate solubilization by Enterobacter intermedium. Curr. Microbiol., 47: 87-92.
CrossRef - Imsande, J., 1998. Iron, sulfur and chlorophyll deficiencies: A need for an integrative approach in plant physiology. Physiol. Plantarum, 103: 139-144.
CrossRefDirect Link - Katiyar, V. and R. Goel, 2003. Solubilization of inorganic phosphate and plant growth promotion by cold tolerant mutants of Pseudomonas fluorescens. Microbiol. Res., 158: 163-168.
CrossRefDirect Link - Kloepper, J.W., R. Lifshitz and R.M. Zablotowicz, 1989. Free-living bacterial inocula for enhancing crop productivity. Trends Biotechnol., 7: 39-44.
CrossRefDirect Link - Leinhos, V. and H. Bergmann, 1995. Influence of auxin producing rhizobacteria on root morphology and nutrient accumulation of crops, pt. 2: Root growth promotion and nutrient accumulation of maize(Zea mays L.) by inoculation with indol-3-acetic acid (IAA) producing Pseudomonas strains and by exogenously applied IAA under different water supply conditions. Angew. Bot., 69: 37-42.
Direct Link - Lopez-Bucio, J., M.F. Nieto-Jacobo, V. Ramirez-Rodriguez and L. Herrea-Estrella, 2000. Organic acid metabolism in plants: From adaptive physiology to transgenic varieties for cultivation in extreme soils. Plant Sci., 160: 1-13.
CrossRefPubMedDirect Link - Luo, A.C., X. Sun and Y.S. Zhang, 1993. Species of inorganic phosphate solubilizing bacteria in red soil and the mechanism of solubilization. Pedospphere, 3: 285-288.
Direct Link - Masalha, J., H. Kosegarten, O. Elmaci and K. Mengel, 2000. The central role of microbial activity for iron acquisition in maize and sunflower. Biol. Fertil. Soils, 30: 433-439.
CrossRef - Narula, N., V. Kumar and R.K. Behl, 2001. Effect of phosphate-solubilizing strains of Azotobacter chroococcum on yield traits and their survival in the rhizosphere of wheat genotypes under field conditions. Acta Agronimica Hungarica, 49: 141-149.
CrossRefDirect Link - Nautiyal, C.S., S. Bhadauria, P. Kumar, H. Lal, R. Mondal and D. Verma, 2000. Stress induced phosphate solubilization in bacteria isolated from alkaline soils. FEMS Microbiol. Lett., 182: 291-296.
CrossRefPubMedDirect Link - Neilands, J.D., 1995. Siderophores: Structure and function of microbial iron transport compounds. J. Biol. Chem., 270: 26723-26726.
CrossRefDirect Link - Pradhan, N. and L.B. Sukla, 2006. Solubilization of inorganic phosphate by fungi isolated from agriculture soil. Afr. J. Biotechnol., 5: 850-854.
Direct Link - Richardson, A.E., 2001. Prospects for using soil microorganisms to improve the acquisition of phosphorus by plants. Aust. J. Plant Physiol., 28: 897-906.
CrossRefDirect Link - Rroco, E., H. Kosegarten, F. Harizaj, J. Imani and K. Mengel, 2003. The importance of soil microbial activity for the supply of iron to sorghum and rape. Eur. J. Agron., 19: 487-493.
CrossRef - Sahin, F., R. Cakmakci and F. Kantar, 2004. Sugar beet and barley yields in relation to inoculation with N2-fixing and phosphate solubilizing bacteria. Plant Soil, 265: 123-129.
CrossRefDirect Link - Sattar, M.A. and A.C. Gaur, 1987. Production of auxins and gibberellins by phosphate dissolving microorganisms. Zentral. Microbiol., 142: 393-395.
Direct Link - Singh, C.P., M.M. Mishra and K.S. Yadav, 1980. Solubilization of insoluble phosphate by thermophilic fungi. Ann. Microbiol., 131: 289-296.
PubMedDirect Link - Trivedi, P. and T. Sa, 2008. Pseudomonas corrugate (NRRL B-30409) mutants increased phosphate solubilization, organic acid production and plant growth at lower temperatures. Curr. Microbiol., 56: 140-144.
PubMedDirect Link - Valencia-Cantero, E., E. Hernandez-Calderon, C. Velazquez-Becerra, J.E. Lopez-Meza, R. Alfaro-Cuevas and J. Lopez-Bucio, 2007. Role of dissimilatory fermentative iron-reducing bacteria in Fe uptake by common bean (Phaseolus vulgaris L.) plants grown in alkaline soil. Plant Soil, 291: 263-273.
CrossRef - Wahid, O.A. and T.A. Mehana, 2000. Impact of phosphate-solubilizing fungi on the yield and phosphorus uptake by wheat and faba bean plants. Microbiol. Res., 155: 221-227.
PubMed - Asad-ur-Rehman, A. Sikander and Ikram-ul-Haq, 2003. Phosphate limitation for enhanced citric acid fermentation using Aspergillus niger mutant UV-M9 on semi-pilot scale. Pak. J. Biol. Sci., 6: 1247-1249.
CrossRefDirect Link - Bilgili, A.V., V. Uygur, S. Karaca, O. Dengiz and S. Aydemir, 2008. Phosphorous sorption in some great soil groups of semi-arid region of Turkey. Int. J. Soil Sci., 3: 75-82.
CrossRefDirect Link - Mishra, A., K. Prasad and G. Rai, 2010. Effect of bio-fertilizer inoculations on growth and yield of dwarf field pea (Pisum sativum L.) in conjunction with different doses of chemical fertilizers. J. Agron., 9: 163-168.
CrossRefDirect Link - Mostafa, G.G. and A.A. Abo-Baker, 2010. Effect of bio- and chemical fertilization on growth of sunflower (Helianthus annuus L.) at South valley area. Asian J. Crop Sci., 2: 137-146.
CrossRefDirect Link - Raji, B.A., 2006. Phosphorus fractions of a dune in the semi-arid savanna of Nigeria. Int. J. Soil Sci., 1: 64-74.
Direct Link - Omar, S.A., 1997. The role of rock-phosphate-solubilizing fungi and Vesicular-Arbusular-Mycorrhiza (VAM) in growth of wheat plants fertilized with rock phosphate. World J. Microbiol. Biotechnol., 14: 211-218.
CrossRefDirect Link