
Stephan Wirth
Leibniz Centre for Agricultural Landscape Research, Institute of Landscape Matter Dynamics
Axel Hohn
Institute of Soil Landscape Research, Eberswalder Str. 84, D-15374 Müncheberg, Germany
Lothar Muller
Institute of Soil Landscape Research, Eberswalder Str. 84, D-15374 Müncheberg, Germany
International Journal of Soil Science
Year: 2008 | Volume: 3 | Issue: 2 | Page No.: 52-61







ABSTRACT
Three different agricultural drainage systems located in the Northeast German lowland i.e., a free drainage of a shallow aquifer, a submerged deep drainage of a shallow aquifer and a free deep drainage were studied in order to detect evidence for translocation of enzyme activities and bacteria in percolating leachates. The activity of fluorescein diacetate (FDA) hydrolase as a measure of over-all microbial activity in the leachates was two up to three orders of magnitude lower as compared to the respective soil substrates. Enzyme activities in the leachates were almost constant throughout the year, except for a significant increase of FDA activity and cellulase activity due to a discharge by preferential flow, detected in the shallow free drainage system. FDA hydrolase was significantly but weakly correlated with chemical properties (NH4N, DOC). The population density of culturable bacteria was rather low (50 up to 300 colony-forming units mL-1 drain water) coinciding with a mostly marginal translocation of soil enzyme activity by leachates from arable drainage systems.
PDF Abstract XML References Citation
How to cite this article
Stephan Wirth, Axel Hohn and Lothar Muller, 2008. Translocation of Soil Enzyme Activity by Leachates from Different Agricultural Drainage Systems
. International Journal of Soil Science, 3: 52-61.
DOI: 10.3923/ijss.2008.52.61
URL: https://scialert.net/abstract/?doi=ijss.2008.52.61
DOI: 10.3923/ijss.2008.52.61
URL: https://scialert.net/abstract/?doi=ijss.2008.52.61
INTRODUCTION
Soil is a dynamic system of inorganic and organic components in varying proportions, which are the result of interactions between complex processes such as the weathering of parent materials, the decomposition and transformation of plant residues and the redistribution of organic and inorganic materials by water movement. The interactions between hydrological, organo-mineral and microbiological factors have crucial impact on geochemical processes and are critical to environmental quality and ecosystem health. Thus, the role of microorganisms inhabitating the complex, surface-rich soil environment has long been intensively studied (Foster, 1988; Voroney, 2007), but the complexity of processes and interactions of microorganisms with mineral surfaces remains hardly understood (Mills and Powelson, 1996; Nunan et al., 2006). Special emphasis was directed on the survival and the translocation, or transport of microorganisms in soils. Ever since molecular genetic approaches became available, microorganisms could be traced in terrestrial environments at the species level (Thies, 2007), which is especially important in context with risk assessments of genetically modified microbial strains (Amarger, 2002; Staley and Brauer, 2006), or the survival and spread of pathogenic microorganisms in soils (Unc and Goss, 2004).
It is well described since many decades, that microbial activities are concentrated in the top soil layers with a s harp decrease of microbial occurrence and activities with increasing soil depth, as determined by vertical distribution patterns of soil organic matter contents or root biomass (Wirth and Wolf, 1992; Salinas-Garcia et al., 1997; Cook and Kelliher, 2006). Evidence has already accumulated that microbial persistence and transport in soil is closely associated with physico-chemical properties, such as the nature of the substratum, the soil solute chemistry and accessibility of particle surfaces (Lawrence and Hendry, 1996; Fletcher and Murphey, 2001), while passive migration of bacteria or spores in the soil profile with the water flow seems to be the dominant (passive) transport mechanism (Huysman and Verstraete, 1993). However, soil microbiology has mostly focused on the plant root zone and has largely ignored the entirety of the vadose zone, the unsaturated zone extending from the soil surface to the groundwater table (Holden and Fierer, 2005). In recent years, however, attention was drawn on the relevance of protein components in soil leachates (Schulze, 2005), but hardly any studies are presently available especially on the translocation of soil organic matter decomposing enzymes with subsurface drain water.
Different land use systems, or land reclamation processes such as drainage, hold the potential to severely impact soil hydraulic properties and thus soil biochemical processes. Most of the hydromorphic soils in the North-East German lowland were drained in the last century, providing different water flow rates and different types of drainage systems i.e., field model systems with ditches and pipe drainage. The water balance as well as water and solute transport of selected sites have been previously studied in lysimeter experiments (Behrendt et al., 2001; Müller et al., 2001; Schindler et al., 2001). The aim of our study was to detect evidence for translocation of microbial activity in soils by leachates in different draining systems on sandy to loamy arable soils. Special emphasis was laid on the analysis of a temporal sequence of draining events, on the effect of different drainage systems and on the analysis of correlations with chemical properties of leachates.
MATERIALS AND METHODS
Study Sites and Soil Properties
The study sites are located in the Pleistocene and Holocene agricultural landscapes of Brandenburg, north-east Germany i.e., in Seelow (Landkreis Märkisch-Oderland), Paulinenaue (Landkreis Havelland) and Dedelow (Landkreis Uckermark). The climate is semi-continental with a mean annual temperature of 8.5°C and an annual precipitation of 480-550 mm. Typical soil types of the Pleistocene area are Luvisols and Stagnosols, formed on sandy to loamy materials. In depressions, Gleysols and Histosols are frequent, whereas in the lowland of the river Oder, Gleysols on clayey parent materials are dominant. Prevailing crops on arable sites are cereals (winter wheat, spring barley), maize and canola, whereas Histosols are mostly under extensive grassland. For this study, lysimeters as model drain sites and moreover, several pipe drains in arable field sites were analysed from February until June 2001, in spring 2002 and again in spring 2006. Thus, different water flow rates were provided by three types of drainage systems. In Figure 1 drain type A (groundwater lysimeter at Seelow) represents sites with shallow water table and free drainage, which are typical for most parts of intensively drained lowland areas, with both matrix flow and very rapid flow. Figure 1 drain type B (groundwater lysimeter at Paulinenaue) shows similar conditions, but with submerged drain systems which are typical for fields with drainage subirrigation systems such as pipe drains, ditches and weirs. At these less intensively drained sites, discharging rain water and solutes pass an anaerobic soil zone. Figure 1 drain type C (moraine field sites at Dedelow) represents typical, most common pipe drainages in the heterogeneous Pleistocene landscape. Drain outflow is free but the discharge contains a mix of water from different sources and ages. Soils of the lysimeters reflect the typical conditions of the surrounding landscapes. Lysimeters of drain type A contain different soils ranging from shallow peat soils and sand gleys to clay gleys (Müller et al., 2001) under arable use. Soils in lysimeters of type B contain peat soils and humic sand gley soils (Behrendt et al., 2001) under grassland. Soils of the drain type C are mainly Luvisols of loamy till rich in calcium (Schindler et al., 2001).
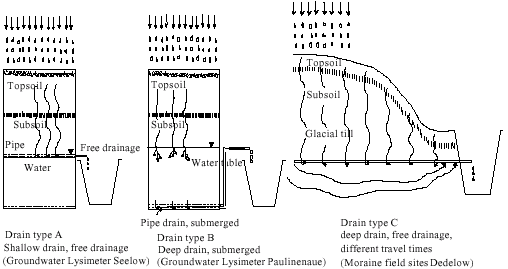 | |
| Fig. 1: | Schematic draft of the drainage systems at the arable sites under study. (A) shallow drain, free drainage, (B) deep drain, submerged drainage and (C) deep drain, free drainage, different travel times |
Microbiological Analyses of Soils and Leachates
Drain water samples were collected in plastic bottles, stored at 4°C and analysed within 48 h. Soil samples were sieved (2 mm) and stored at 4°C up to 5 days. Soil enzymes were extracted by suspending soil (1 g) with 25 mL HEPES Puffer (0.5 M, pH 7) and stirring for 60 min at 20°C (300 rpm). Extracts were finally centrifuged (10 min, 10000 g, 4°C) and stored on ice. Fluorescein diacetate (FDA) hydrolase as a non-specific measure of total microbial activity (Schnürer and Rosswall, 1982) was assayed using a micro-plate approach as described by Wirth (1992). In detail, 50 μL buffer (HEPES, 0.2 M, pH 7), 50 μL substrate solution (FDA, 100 μg mL-1) and 100 μL sterile filtered drain water or soil extract sample were incubated in triplicate at 40°C up to 4 h. After the addition of ice water (50 μL) to terminate the reaction, plates were directly assayed spectrophotometrically at 492 nm. One unit of FDA hydrolase activity was calculated as absorbancex1000xmin-1. Endo-acting cellulase activity of leachates was assayed colourimetrically via the detection of reducing sugars with the 3,5-dinitrosalicilic acid reagent (DNSA, Miller et al., 1960), adapted to 1.5 mL reaction tubes (Wirth, 1991). In detail, 100 μL HEPES buffer (0.2 M, pH 7), 100 μL carboxymethyl-cellulose substrate solution (CMC, Serva, Germany; 10 mg mL-1) and 200 μL sterile filtered leachate samples were incubated at 40°C up to 24 h. After the addition of 0.6 mL DNSA reagent and heating of reaction mixtures in a water bath (15 min, 100°C), samples were assayed spectrophotometrically at 550 nm. Based on glucose standards (5 up to 500 μmol glucose monohydrate), one unit of endo-cellulase activity was defined as equivalent to the release of 1.0 μmol glucose h-1. The FDA hydrolase and cellulase assay conditions were optimised in previous experiments with respect to pH, incubation temperature and substrate concentration.
The total numbers of culturable bacteria in drain waters was determined as colony forming units (cfu) by a dilution pour-plate method (Ulrich and Wirth, 1999), using tryptic soy broth, pH 7 (Difco Laboratories, Detroit, USA; 0.2 g L-1) and bacto-Agar (Difco, 20 g L-1). Undiluted water samples (1 mL) were inoculated into five replica Petri dishes and swirled with agar medium at 50°C. Solidified agar plates were incubated for up to 5 day at 20°C in the dark and counted for cfu against diffuse light.
Chemical Analyses of Leachates
All drain water samples were collected in plastic bottles, which had been previously acid washed and soaked in deionized water for 24 h. Samples were stored at 4°C prior to analysis, which occurred within a few weeks. All solution analyses, except pH, were measured after filtration (0.45 μm). The pH was measured with a glass electrode using the WTW pH Ionmeter pMX 2000. Redoxpotential was determined with a Schott pH-Meter CG 837. The electrical conductivity was measured with the WTW Ionactivitymeter LF 95. Ammonium, nitrate and phosphorus were detected colorimetrically using the Eppendorf EPOS 5060 autoanalyser. Sulphate and chloride were analysed by Ion chromatography Dionex Dx 320. Calcium was determined by Flame Atomic Absorption Spectrophotometry Unicam Solar 939.
RESULTS AND DISCUSSION
Chemical Properties of Leachates
The pH values of leachates were generally in the neutral to slightly alkaline range (pH 7.32 -7.63) and were uniformly distributed throughout the different drain types (Table 1). The redoxpotential was low in drain types A and B, which points to anaerobic or microaerophilic conditions in these soils, whereas the redoxpotential in drain C leachates was more than twice as high compared to A and B leachates. Samples from drain type B show the highest electrical conductivity values (2.53 mS cm-1), caused by high concentrations of chloride and calcium ions of the corresponding drain waters. The ion concentration of drain leachates from type A and C were similar and between 2.5 and 3.6 times lower than drain type B waters. With an average content of 53 mg L-1 Dissolved Organic Carbon (DOC), the drain type B water samples contained 2.8 to 5.2 times higher concentrations than drain waters from A and C types which is linked to the organic soil type at the lysimeter site. The ammonium concentrations of the drain waters had a wide concentration range between 0.01 mg L-1 (type C) and 2.1 mg L-1 (type A) with intermediate contents of 0.98 mg L-1 for type B. For nitrate, drain type A leachates again had the highest content but in contrast to ammonium, drain type B leachates had extremely low nitrate concentrations whereas the type C leachates had medium contents. Overall, the leached concentrations of DOC, ammonium and nitrate were in a range commonly reported for soil solutions of arable soils, i.e., 15-50 mg L-1 DOC, 0.2-4 mg L-1 NH4+ and 20-200 mg L-1 NO3– (Scheffer and Schachtschabel, 2002). In water samples from type B drains very high chloride concentrations were detected (mean value = 449 mg L-1), whereas chloride amounts in type A and type C drain waters were 4.8 and 6.9 times lower. The high chloride contents of type B drain waters are probably due to the longterm input of drinking water to the lysimeter soils. In contrast, the sulphate content of type B leachates were the lowest. This finding is considered to be a consequence of high adsorption capacities for sulphate in organic soils. The sulphate concentrations in type A and type C leachates were in the same range but 2.3 to 3 times higher. The same picture was found for the phosphorus distribution, i.e., relatively low concentration in type B water samples and similar amounts in type A and C drain waters, but 4.4 to 4.9 times higher in concentration. The range of calcium concentrations in the different drainwater types was not very distinct. The sequence with increasing concentration was: type C < type A < type B. The dominant anion in type C water samples was sulphate (SO4/Cl = 2.2), in type A leachates the portion of each anion is similar (SO4/Cl = 1.2) and in type B leachates chloride was the dominant anion (SO4/Cl = 0.1). Soils that exhibit a substantial amount of SO4 adsorption are considered to be resistant to accelerated cation leaching (Harrison et al., 1989), but this could not be confirmed by type B leachates, as these samples showed the highest Ca concentration compared to type A and C leachates.
| Table 1: | Chemical composition of different drain water types |
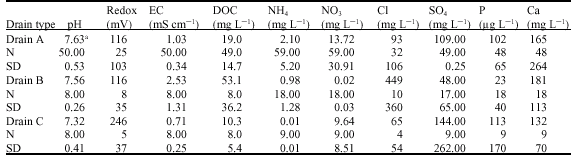 | |
| a: Mean values, N: No. of samples, SD: Standard deviation | |
| Table 2: | Extractable fluorescein diacetate hydrolase activities in different soil substrates and respective activities in leachates |
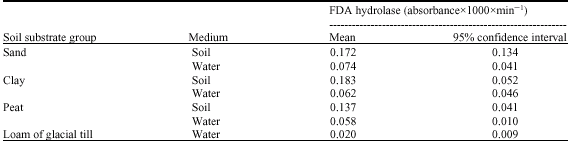 | |
Enzyme Activities in Soils and Respective Leachates
Extracts from soil samples and corresponding drain water samples were assayed for the activity of FDA hydrolase to provide a measure of over-all microbial activity, including the activities of esterases and proteases (Schnürer and Rosswall, 1982). Thus, a soluble extracellular enzyme complex was analyzed, which is potentially prone to vertical transport. With respect to the soils and sites under study, FDA hydrolase activities were highly variable and in a similar range for sand and clay samples, while activities in peat were lower compared to the mineral substrates (Table 2). The FDA hydrolase activities in the leachates were two up to three orders of magnitude lower as compared to the respective soil substrates. An additional leachate sample derived from a moraine field site displayed significantly lowest FDA hydrolase activity. Concerning the different drain types under study, FDA hydrolase activities in leachates derived from lysimeters were up to threefold higher as compared to a field drain (Table 3, drain type C). The main factors determining soil enzyme activity are stocks and availability of substrate, or other soil properties such as acidity or temperature. Consequently, close associations between FDA hydrolase activity and organic carbon were described, e.g., in forest soil (Wirth, 1992), but in our leachate samples only weak correlations were found with DOC concentrations (Table 4), probably due to the high variability and comparatively low activities. Low activities of FDA hydrolase were also confirmed in corresponding leachate samples at a later date in May 2006 (data not detailled here). In a general view, enzymes active in soil are extracellular and highly stabilized to mineral or organic surfaces (Burns and Dick, 2002), or forming enzyme-substrate complexes. Consequently, soil enzymes are considered to be protected from transport by free percolating soil water, but our results give evidence for considerable translocation of a hydrolytic enyzme. Only few reports are available on direct evidence of enzyme activity in soil leachates (e.g., Toor et al., 2003), whereas preferential flow paths are already recognized as biological hotspots in soil (Bundt et al., 2001). Further studies are required to underpin these findings, such as tracing marked enzymes to determine the pattern of transport through intact lysimeter soil cores.
Enzyme Activities at Different Drainage Events
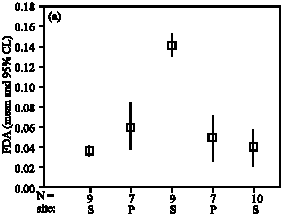
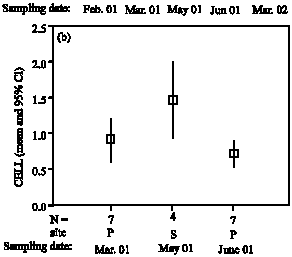
FDA hydrolase activities of drain water samples were generally low in late winter or even hardly detectable after prolonged incubation times (24 h, 40°C, pH 7). Maximum activities were found in
| Table 3: | Fluorescein diacetate hydrolase activities in leachates from different drainage systems |
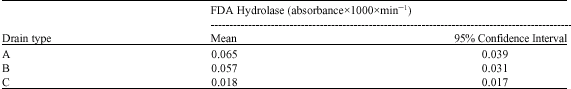 | |
| Table 4: | Significant correlations between Fluorescein diacetate hydrolase in the drainwater and chemical properties of drainwater (All samples, n = 60, α = 0.05) |
 | |
Parameters not listed are not significant early summer (May), but activities decreased again in June (Fig. 2a). Similar values of FDA hydrolase activities in early spring were found in the following year and confirmed in spring 2006 (data not detailled here). Correspondingly, the activities of endo-cellulases in leachates were low in spring and highest in early summer samples (Fig. 2b), but due to high variability without significant differences. In general, we found evidence for almost constant enzyme activities in soil leachates throughout the year, except a significant higher FDA activity and increased cellulase activity in May 2001, due to a discharge immediately after a heavy rainstorm of 40 mm resulting in preferential flow. Evidently, the soil pore water flow conditions are critical for potential transport of enzymes, but more analyses during a vegetation period would be required to document such findings more clearly. Preferential flow is a common phenomenon in soils with pathways for infiltrating water which can be very persistent with time (Hagedorn and Bundt, 2002). Preferential transport was reported for anions in clay pan soil, accounting for 35 % of the total transport (Wilkison and Blevins, 1999). In other cases, pesticides were transported in drain water by preferential flow (Michaelsen, 1998), or nitrate and pesticide were transported after periods of excess precipitation (Bosch and Truman, 2002), but to our knowledge no reports are available about translocation of hydrolytic enzymes with preferential flow. Besides excessive rainfall, also partly frozen soil needs to be considered to be responsible for rapid discharge of water (Derby and Knighton, 2001). Moreover, evidence was reported for the relative contribution of macropores to the movement of water and transport of organic compounds in dried tire soil (Shipitalo and Edwards, 1996). Thus, we conclude from present study, that more observance should be directed on soil enzymes as these biocatalytic compounds can be subject to vertical translocation with water flow under different soil conditions.
Abundance of Bacteria in Leachates
In order to provide more evidence for the generally low enzyme activities in leachates, bacterial densities were analysed in several drain waters by using an agar plate technique. As a result, population densities of 50 up to 300 colony-forming units (cfu) of bacteria were determined mL-1 of leachate samples but with a high variability between variants (Fig. 3). In general, the detected densities of bacterial cfu were rather low, few samples contained >100 cfu mL-1 which is in a range of cfu concentrations reported for potable waters (Ramalho et al., 2001). In contrast, densities of bacterial cfu of arable or forest top soil layers are commonly reported to contain 106 up to 107 cfu g-1 soil (Kauri, 1983; Ulrich and Wirth, 1999). Thus, soil bacteria are supposed to be rather unlikely transported vertically by percolating water, or leachates supporting general view that
| |||
| Fig. 2: | FDA hydrolase activities (A) and cellulase activities (B) of leachates at different drainage periods. FDA: Fluorescein diacetate hydrolase activity (absorbance x 1000x min-1), CELL: cellulase activity (1.0 μmol glucose h-1). Site S: Lysimeter Seelow, site P: Lysimeter Paulinenaue | ||
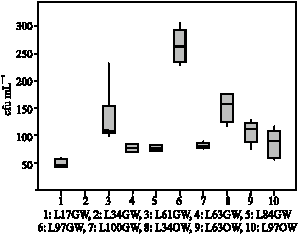 | |
| Fig. 3: | Population densities of bacteria in drain water and surface water samples. cfu: colony-forming units. TSA: Trypticase Soy Agar. 1-7: lysimeter samples, 8-10: surface samples |
microorganisms are sticking to surfaces of particles within the soil matrix, or are even enmeshed in micro-sites with a steep gradient of microbial population densities with increasing soil depth in most cases (Mills and Powelson, 1996). Experimental evidence confirm the view of rather restricted vertical transport of microbial cells in soil, but depending on soil properties (Huysman and Verstraete, 1993) or depending on the presence of preferential flow paths (Bundt et al., 2001). On the other hand, evidence was provided for vulnerability of soils to leaching of microbes, such as faecal coliforms, into local surface and groundwater (McLeod et al., 2003). Further studies would be required to explore translocation of soil bacteria on species level with special focus on the abundance of unculturable bacteria.
Overall, we thus conclude from present study, that leachates from arable drainage systems are unlikely to translocate considerable amounts of enzymes, or bacteria originating from top soil layers, except for preferential flow events that may cause translocation especially in shallow free drainage systems.
ACKNOWLEDGMENTS
This study was supported by the Federal Ministry of Consumer Protection, Food and Agriculture (BMVEL) and the Ministry of Agriculture, Environmental Protection and Area Planning of the State Brandenburg (MLUR). Chemical analyses were performed by ZALF Central Laboratory.
Dr. Axel Behrendt and Dr. Uwe Schindler supported collection of data and analysis of some lysimeter samples. Coordination of sampling and data analysis, field testing of water samples, creating and checking of primary data files was done by Mrs. Dipl. Ing. (FH) Ute Moritz.
REFERENCES
- Amarger, N., 2002. Genetically modified bacteria in agriculture. Biochimie, 84: 1061-1072.
Direct Link - Bosch, D.D. and C.C. Truman, 2002. Agrochemical transport to groundwater through Coastal plain soils. Trans. ASAE, 45: 1385-1396.
Direct Link - Bundt, M., F. Widmer, M. Pesaro, J. Zeyer and P. Blaser, 2001. Preferential flow paths: Biological hot spots in soils. Soil Biol. Biochem., 33: 729-738.
Direct Link - Cook, F.J. and F.M. Kelliher, 2006. Determining vertical root and microbial biomass distributions from soil samples. Soil Sci. Soc. Am. J., 70: 728-735.
Direct Link - Derby, N.E. and R.E. Knighton, 2001. Field- scale preferential transport of water and chloride tracer by depression-focused recharge. J. Environ. Qual., 30: 194-199.
Direct Link - Foster, R.C., 1988. Microenvironments of soil microorganisms. Biol. Fert. Soils, 6: 189-203.
CrossRefDirect Link - Hagedorn, F. and M. Bundt, 2002. The age of preferential flow paths. Geoderma, 108: 119-132.
Direct Link - Harrison, R.B., D.W. Johnson and D.E. Todd, 1989. Sulfate adsorption and desorption reversibility in a variety of forest soils. J. Environ. Qual., 18: 419-426.
Direct Link - Holden, P.A. and N. Fierer, 2005. Microbial processes in the vadose zone. Vadose Zone J., 4: 1-21.
Direct Link - Huysman, F. and W. Verstraete, 1993. Water-facilitated transport of bacteria in unsaturated water columns: Influence of cell surface hydrophobicity and soil properties. Soil Biol. Biochem., 25: 83-90.
CrossRef - Kauri, T., 1983. Fluctuations in physiological groups of bacteria in the horizons of a beech forest soil. Soil Biol. Biochem., 15: 45-50.
CrossRef - Lawrence, J.R. and M.J. Hendry, 1996. Transport of bacteria through geological media. Can. J. Microbiol., 42: 410-422.
Direct Link - McLeod, M., J. Aislabie, J. Ryburn, A. McGilla and M. Taylor, 2003. Microbial and chemical tracer movement through two Southland soils, New Zealand. Aust. J. Soil Res., 41: 1163-1169.
CrossRefDirect Link - Miller, G.L., R. Blum, W.E. Glennon and A.L. Burton, 1960. Measurement of carboxymethylcellulase activity. Anal. Biochem., 2: 127-132.
CrossRef - Nunan, N., K. Ritz, M. Rivers, D.S. Feeney and I.M. Young, 2006. Investigating microbial micro-habitat structure using X-ray computed tomography. Geoderma, 133: 398-407.
Direct Link - Ramalho, R., J. Cunha, P. Teixeira and P. Gibbs, 2001. Improved methods for the enumeration of heterotrophic bacteria in bottled mineral waters. J. Microbiol. Meth., 44: 97-103.
Direct Link - Salinas-Garcia, J.R., F.M. Hohns and J.E. Matocha, 1997. Long-term effects of tillage and fertilization on soil organic matter dynamics. Soil Sci. Soc. Am. J., 61: 152-159.
Direct Link - Schulze, W.X., 2005. Protein analysis in dissolved organic matter: What proteins from organic debris, soil leachate and surface water can tell us-a perspective. Biogeoscience, 2: 75-86.
Direct Link - Shipitalo, M.J. and W.M. Edwards, 1996. Effects of initial water content on macropore/matrix flow and transport of surface- applied chemicals. J. Environ. Qual., 25: 662-670.
Direct Link - Staley, T.E. and D.K. Brauer, 2006. Survival of a genetically modified root-colonizing pseudomonad and rhizobium strain in an acidic soil. Soil Sci. Soc. Am. J., 70: 1906-1913.
Direct Link - Toor, G.S., L.M. Condron, H.J. Di, K.C. Cameron and B.J. Cade-Menun, 2003. Characterization of organic phosphorus in leachate from a grassland soil. Soil Biol. Biochem., 35: 1317-1323.
Direct Link - Ulrich, A. and S. Wirth, 1999. Phylogenetic diversity and population densities of culturable cellulolytic soil bacteria across an agricultural encatchment. Microb. Ecol., 37: 238-247.
CrossRefDirect Link - Unc, A. and M.J. Goss, 2004. Transport of bacteria from manure and protection of water resources. Applied Soil Ecol., 25: 1-8.
Direct Link - Wilkison, D.H. and D.W. Blevins, 1999. Observations on preferential flow and horizontal transport of nitrogen fertilizer in the unsaturated zone. J. Environ. Qual., 28: 1568-1580.
Direct Link - Wirth, S.J., 1992. Detection of soil polysaccharide endo-hydrolase activity profiles after gel permeation chromatography. Soil Biol. Biochem., 24: 1185-1188.
Direct Link - Wirth, S.J. and G.A. Wolf, 1992. Micro-plate, colourimetric assay for endo-acting cellulase, xylanase, chitinase, 1,3-β-glucanase and amylase extracted from forest soil horizons. Soil Biol. Biochem., 24: 511-519.
CrossRef