
B.I. Aderiye
Department of Microbiology, University of Ado-Ekiti, P.M.B. 5363, Ado-Ekiti, Nigeria
S.A. Laleye
Department of Microbiology, University of Ado-Ekiti, P.M.B. 5363, Ado-Ekiti, Nigeria
O.R. Ijalana
Department of Food Science and Technology, Federal University of Technology, Akure, Nigeria
International Journal of Soil Science
Year: 2008 | Volume: 3 | Issue: 1 | Page No.: 42-47







ABSTRACT
The mycoflora of the soil of some commercial ventures such as cassava farm, cattle ranch, cocoa farm and sawmill was investigated. Aspergillus sp., Penicillium sp. were isolated from cassava farm and cattle ranch, while Absidia sp., Curvularia sp., Fusarium sp., Neurospora sp. and Rhizopus stolonifer were isolated from the cocoa and sawmill soils. Sawmill soil recorded the highest moisture and organic matter contents. The comparatively high pH value of the cattle ranch soil was attributed to large volume of urine and faeces of the cattle in the ranch. Calcium and potassium ions were the most prominent mineral nutrients especially in the cocoa farm and sawmill soils. Only Fusarium sp. was highly encouraged when cultured on soil extracts from the cassava farm, cocoa farm and sawmill. Spore germination, initiation of germ tubes and elongation of hyphae of Aspergillus niger and Curvularia sp. were adversely affected in all the soil extracts. However, the sawmill soil encouraged the production of an appreciable mycelia growth in species of Fusarium, Neurospora and Penicillium.
PDF Abstract XML References Citation
How to cite this article
B.I. Aderiye, S.A. Laleye and O.R. Ijalana, 2008. Soil Mycoflora of Some Commercial Ventures in South West Nigeria. International Journal of Soil Science, 3: 42-47.
DOI: 10.3923/ijss.2008.42.47
URL: https://scialert.net/abstract/?doi=ijss.2008.42.47
DOI: 10.3923/ijss.2008.42.47
URL: https://scialert.net/abstract/?doi=ijss.2008.42.47
INTRODUCTION
Each soil type has its own distinctive features, colour, texture and structure, in different layers. The topsoil contains a wide variety of microorganisms; algae; bacteria, fungi and protozoa and a host of other inhabitants such as the invertebrates that are highly degradative in nature (Brady, 1990; Sappänen et al., 2007). The layer also supports plant life, which after death undergoes a lot of biodegradative activity (Kronberg et al., 1979). The topsoil contains the most important aerobic microbes where cellulose wastes also accumulate. Many fungi are able to digest cellulose, which constitutes the major plant materials on the forest floor. Most soils and decaying matter habour a fairly defined flora of filamentous fungi (Leglerg, 1980), whose population is less than that of bacteria (Slater and Winpenny, 1983). Furthermore, fungi and bacteria that act as main primary consumers of decomposable materials in soil (Wardle et al., 2004) may be highly sensitive to changes in lignin content and composition of the decomposing plant material.
The ability of different types of microorganisms to utilize the same mineral nutrient is not uniform. Smirnova (1974) reported that cadmium, nickel, silver, copper and zinc are easily utilized by Aspergillus niger and Penicilium glacum. A concentration of copper sulphate CuSO4 greater than 100 mg L-1 suppressed the growth of seven forms of Candida, two forms of Torulopsis and Saccharomyces cerevisae (Kerridge, 1980). However, the utilization of these mineral nutrients depends to a large extent on high soil pH levels, low activity of soil enzymes, dehydrogenase and alkaline phosphatase, higher microbial populations and composition of the medium (Donegan et al., 1999).
Plant and animal waste materials are deposited on the topsoil of environments of small scale commercial ventures such as sawmill, cattle ranch and some farms that are situated near residential buildings, typical of an average village/rural setting in West Africa. Microbial community composition can be more sensitive to amendments with plant residue than microbial biomass (Eriksen, 2005). This study reports on the mycoflora of soils of some commercial ventures and their degradative ability. The role of the soil extract in supporting the growth of the fungal isolates is also discussed.
MATERIALS AND METHODS
Source of Soil Samples
Soil samples were collected from different locations in Akure town and its environs in Ondo State, southwest Nigeria. The sites include sawmill and cocoa farm at Orita-Obele; cassava farm and cattle ranch of the Federal University of Technology, Akure Teaching and Research Farms. Samples were collected from 8 different sites on each location. Sampling was carried out at the onset of rains in March for two consecutive years. At each site, samples were collected at a depth of between 5-20 cm beneath the soil surface using clean sterile spatula and collection bags. The mean of data obtained from these random sampling were recorded and analyzed statistically.
Fungi from the samples were isolated and purified by repeated sub-culturing on 1% cycloheximide in Malt Extract Agar (MEA) (Merck-Germany) and soil extract media. Isolates were identified according to the methods of Aderiye et al. (2006). All the isolates were stored on Malt Extract agar slants and kept in refrigerator until needed.
Determination of Physicochemical Characteristics
Fresh soil samples were analyzed separately for moisture, pH and titratable acidity as described by Aderiye (1985). The oxidizable organic matter was determined using the modified Walley-Blacks procedure (AOAC, 1990) with Cr2O7 oxidizing the oxidizable portion of the organic matter. The reaction was facilitated by the heat generated when sulphuric acid (H2SO4 and K2Cr2O7 were mixed in a ratio of 2:1(v/v). The excess Cr2O7 was determined by titrating with standard ferrous sulphate solution. The amount of substance oxidized was calculated from the amount of Cr2O7 reduced using ortho-phenanthroline-ferrous complex indicator. A colour change from orange to dark green to light green and finally maroon red was observed. The amount obtained from the titer was multiplied by 1.33 to give the percentage organic carbon.
Determination of Soil Mineral Nutrients
Mineral nutrients from air-dried soil samples were extracted using Ammonium acetate solution (prepared from glacial acetic acid and concentrated ammonium hydroxide, made into solution with the addition of distilled water. The resulting solution was used as a sample for determination of various mineral elements present in the samples. Sodium and potassium levels were determined using flame photometer as described by AOAC (1990). Calcium and magnesium were determined by EDTA titration. While Zinc was determined using an Atomic Absorption Spectrophotometer (AAS-Bucks 210) as described by Adeyeye (1997).
Determination of Fungal Growth in Soil
About 10 g of each fresh soil sample was suspended in 90 mL distilled water, homogenized and filtered. To the filtrate was added two gram of dextrose sugar (Analar) and 1% microbiological peptone (Oxoid). The solution was dispensed into test tubes for broth medium preparation. Agar powder (1.5%, Oxoid) was added to another portion of the filtrate in Erlenmeyer flask. One percent cycloheximide was aseptically added to the medium whenever required. Malt extract agar and ME broth (Merck-Germany) served as controls.
The determination of radial mycelia growth and mass were done as described by Aderiye et al. (1998a). All experiments were carried out in triplicates and repeated twice.
RESULTS AND DISCUSSION
A total of about 40 soil samples were analyzed and the following fungi were isolated: Aspergillus niger, A. flavus, Rhizopus stolonifer, Absidia sp., Curvularia sp., Fusarium sp., Neurospora sp. and Penicillium sp. (Table 1). Most of the fungi were isolated from the cassava farm and the sawmill soil samples. However, Aspergillus and Penicillium sp. were frequently isolated in soil samples obtained from the cassava farm, cattle ranch and cocoa farm. The frequency of isolation of Aspergillus and Penicillium sp. may be attributed to their ability to breakdown a wide variety of substrates that are inherent in their high extra cellular enzyme activities on complex polysaccharide (Garrett, 1981; Aderiye et al., 2000, 2006). Sappänen et al. (2007) however posited that soil enzyme activities were enhanced with fresh green manure amendments, i.e., direct incorporation and mulching but not by processed green manure forms i.e., slurry and compost.
Analysis of sawmill soil revealed very high moisture and organic matter contents. Wood components like lignin and cellulose are more difficult to degrade (Begiun and Aubert, 1994). These may contribute to the water retaining capacity of these soils. The high occurrence of fungi in the sawmill soil may be as a result of the ease with which water present in the environment is extracted. It has been reported that the growth of terrestrial fungi is dependent on water availability than the amount of water present (Garrett, 1981). Aspergillus sp. and other Basidiomycotina are unable to grow at high water potentials (Eriksen, 2005) which may explain their high occurrence in cattle ranch and cassava soil samples (Boddy, 1983). Sawdust and wood chippings which are some of the bye-products of sawmill activity have high water retaining ability and also prevent evaporation of moisture from the soil underneath (Begiun and Aubert, 1994). Meanwhile, soil obtained from the cattle ranch had the least moisture and organic matter contents (Table 2).
| Table 1: | Fungi isolated from different soil samples |
 | |
| +: Denotes presence of fungi; -: Denotes absence of fungi | |
| Table 2: | Total titratable acid, pH, moisture and organic Matter contents of the soil samples |
 | |
| *: Mean of data obtained from 8 sample points | |
| Table 3: | Mineral nutrient (mg x10-2 g-1) of the soil samples* |
 | |
| *: Mean of data obtained from 8 sample points | |
The presence of other biological forms such as nematodes and other invertebrates may also limit the presence of fungal species except for those fungi that possess some survival strategies as in the production of resting spores, microconidia and chlamydospores (Aderiye and Ogundana, 1986).
The cattle ranch soil had very high pH value of 9.4 while that of the cocoa farm soil was 6.60. The high pH value in the former may be attributed to the presence of the dung and urine of the cattle deposited in the ranch. It was observed that individual cattle discharged large volumes of urine at a time. If the total urine discharged in a particular environment is taken into consideration, then it is possible to shift the soil pH, more so that urine has an alkaline pH.
However, the high concentration of total titratable acid in cocoa soil may be as a result of the microbial and other biological activities on the leaves and other plant debris that constitute part of the soil structure. Celluloses and lignocelluloses have been reported to constitute about 40-50% of an average soil component (Wood, 1985; Eriksson et al., 1990; Begiun and Aubert, 1994; Aderiye et al., 1998b, 2000). The end product of these biodegraded complex polysaccharide includes organic acids.
In all the soils investigated, calcium (Ca++) and potassium (K++) ions were the most prominent of the soil mineral nutrients (Table 3). The cocoa and sawmill soils were very rich in calcium, while potassium ions were also detected from the cattle ranch. Calcium oxalate crystals have been reported to be found entrusted outside fungal hyphae which may have excretory or antibiotic functions (Cooke and Rayner, 1984). Also, the high values obtained for calcium in the cocoa and sawmill soils may be as a result of different biological life such as invertebrates under the soil. Most of these organisms die off and their exoskeletons which contain calcium are degraded and recycled into soil. Calcium has also been reported to be required for the stabilization of membrane in some lower fungi (Deacon, 1998). Potassium on the other hand is necessary as a cofactor in photosynthesis and respiration in plants (Taylor et al., 1998). The high incidence of potassium in the cattle ranch may be as a result of the digested plant products that were excreted as dung by the cattle.
Availability of carbon ![]() substrates largely controls microbial growth in soil. It is a key factor governing nitrogen (N), phosphorous (P) and sulphur (S) cycles. Mineralization of P and S, mediated by phosphatases and arylsulphatases, is also driven by microbial demand for P and S independently of C. Nitrogen can also be mineralized above the demand of the biota, due to incidental mineralization of N during decomposition (c mineralization (Sappänen et al., 2007). The availability of other mineral nutrients such as sodium (Na) and zinc (Zn) ions in relatively low amounts (0.023-0.07 mg g-1) may be a result of cation exchange within the soil (Alexander, 1986).
substrates largely controls microbial growth in soil. It is a key factor governing nitrogen (N), phosphorous (P) and sulphur (S) cycles. Mineralization of P and S, mediated by phosphatases and arylsulphatases, is also driven by microbial demand for P and S independently of C. Nitrogen can also be mineralized above the demand of the biota, due to incidental mineralization of N during decomposition (c mineralization (Sappänen et al., 2007). The availability of other mineral nutrients such as sodium (Na) and zinc (Zn) ions in relatively low amounts (0.023-0.07 mg g-1) may be a result of cation exchange within the soil (Alexander, 1986).
Only Fusarium sp. was so much encouraged by the extracts of the soil obtained from the cassava farm, cocoa farm and the sawmill. This may be attributed to an efficient selective uptake of nutrients by this microbe that resulted in increased growth advantage (Kushner, 1978). Spore germination and initiation of germ tubes/elongation of hyphae in Aspergillus niger and Curvularia sp. were adversely affected in all the soil extracts broth. This may be as a result of the non-uptake of nutrients in solution at certain pH level or the nutrients may be in undissociated forms (Deacon, 1998). However, the sawmill soil was quite favourable for the production of an appreciable mycelia mass in the species of Fusarium (242 mg), Neurospora (236 mg) and Penicillium (89 mg) (Table 4).
Most of the organisms showed poor growth when cultivated on cassava farm and cattle ranch soils. These soils may contain natural inhibitory substances/compounds that prevent the use of the resources in that environment. Examples of such compounds include tannins, flavonoids, terpenoid, tropolones and stilbenes. This observation agrees with an earlier report of Cooke and Rayner (1984) on decay of heartwood in trees. However, only the species of Curvularia, Fusarium and Penicillium grew well on the different soil extract agar, exhibiting an appreciable increase of between 106.3/212.5% in the growth of hyphae in cattle ranch/cocoa soil; 100/381.8% in cattle ranch/cocoa soil and 120/200% in cassava/cocoa soils, respectively (Table 5). Both cocoa farm and sawmill soils supported the elongation of hyphal strands of all the fungi. The cattle ranch soil did not encourage much fungal growth except in Curvularia and Fusarium sp.
| Table 4: | Mycelia weight (mg) of fungi grown on soil extract broth |
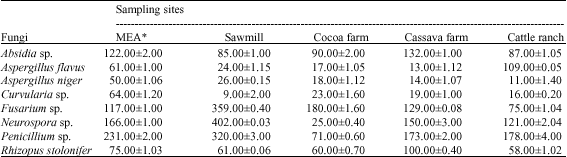 | |
| *: Control | |
| Table 5: | Growth (mm) of fungi on soil extract agar |
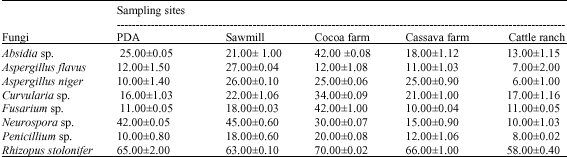 | |
| *: Control; #: Mean of data obtained from 8 sampling points for each site | |
The presence of Penicillium and Aspergillus sp. in the cassava and cattle ranch soils confirms the ubiquity of these environmental contaminants, even in a relatively high alkaline medium.
REFERENCES
- Aderiye, B.I., S.A. Laleye and B. Ojo, 1998. Toxicity of citric and succinic acids on the pycnidiospores of Botryodiplodia theobromae. Folia Microbiol., 43: 147-150.
Direct Link - Aderiye, B.I., I.O. Ogunleye and A.O. Ayeni, 2000. Biochemical changes of cashew pomace during submerged fermentation. Discovery Innovat., 12: 88-91.
Direct Link - Aderiye, B.I., S.A. Laleye and H.A. Akinduro, 2006. Spoilage of some stored fermented foods in Southwest Nigeria. J. Biol. Sci., 6: 659-663.
CrossRefDirect Link - Adeyeye, E.I., 1997. The effect of heat treatment on the in-vitro multienzyme digestibility of protein of six varieties of African yam bean (Sphenostylis stenocarpa) flour. Food Chem., 60: 509-512.
CrossRefDirect Link - Boddy, L., 1983. Effect of temperature and water potential on growth rate of wood-rotting basidiomycetes. Trans. Br. Mycol. Soc., 80: 141-149.
CrossRefDirect Link - Donegan, K.K., R.J. Seidler, J.D. Doyle, L.A. Porteous, G. Digiovanni, F. Widmer and L.S. Watrud, 1999. A field study with genetically engineered alfalfa inoculated with recombinant Sinorhizobium meliloti: Effects on the soil ecosystem. J. Applied Ecol., 36: 920-936.
CrossRefDirect Link - Eriksen, I., 2005. Gross sulphur mineralisation-immobilisation turnover in soil amended with plant residues. Soil Biol. Biochem., 37: 2216-2224.
CrossRefDirect Link - Kronberg, B.I., W.S. Fyfe, O.H. Leonardos and A.M. Santos, 1979. The chemistry of some Brazilian soils: Element mobility during intense weathering. Chem. Geol., 24: 211-229.
CrossRefDirect Link - LeClerg, E.L., 1980. Cultural studies of some soil fungi. Mycologica, 22: 186-210.
CrossRefDirect Link - Sappänen, S.K., H.L. Pasonen, S. Vauramo, J. Vahala and M. Toikka et al., 2007. Decomposition of the leaf litter and mycorrhiza forming ability of silver birch with a genetically modified lignin biosynthesis pathway. Applied Soil Ecol., 36: 100-106.
CrossRefDirect Link - Wardle, D.A., R.D. Bardgett, J.N. Klironomos, H. Setälä, W.H. van de Putten and D.H. Wall, 2004. Ecological linkages between aboveground and belowground biota. Science, 304: 1629-1633.
CrossRefPubMedDirect Link