
B. Baninasab
Department of Horticulture, College of Agriculture, Shiraz University, Shiraz, Iran
M. Rahemi
Iranian Pistachio Research Institute, Rafsanjan, Iran
A. Javanshah
Iranian Pistachio Research Institute, Rafsanjan, Iran
International Journal of Soil Science
Year: 2007 | Volume: 2 | Issue: 1 | Page No.: 40-47







ABSTRACT
This study was conducted to test the effects of different concentrations (0, 0.3 and 0.6%) of nitrogen and application dates (May 6, June 17 and July 18) on flower bud retention of pistachio (cv. Owhadi) trees. The results showed that nitrogen significantly increased the flower bud retention, nitrogen level of leaf, current shoot and bud, photosynthesis rate, chlorophyll content and leaf area. Nitrogen was more effective in increasing flower bud retention of pistachio trees when applied at 0.3% on June 17. Abscission of flower buds decreased progressively as nitrogen content in leaf, current shoot and flower bud and also leaf area increased. It was concluded that application of nitrogen to the pistachio trees might be a useful method in decreasing flower bud abscission and increasing crop in the following year.
PDF Abstract XML References
How to cite this article
B. Baninasab, M. Rahemi and A. Javanshah, 2007. Effects of Time of Foliar Application of Nitrogen and its Concentrations on the Flower Bud Retention in Pistachio Trees. International Journal of Soil Science, 2: 40-47.
DOI: 10.3923/ijss.2007.40.47
URL: https://scialert.net/abstract/?doi=ijss.2007.40.47
DOI: 10.3923/ijss.2007.40.47
URL: https://scialert.net/abstract/?doi=ijss.2007.40.47
INTRODUCTION
Although alternate bearing is a common phenomenon in temperate fruit trees, the mechanism triggering alterance between ON and OFF years in pistachio (Pisatcia vera L.) is unique. In contrast to other fruit tree species, in which fruiting inhibits the formation of flower buds, pistachio forms abundant flower buds on current season’s shoots every year, which abscise during summer, only on fruiting trees or branches (Vemmos et al., 1994; Roussos et al., 2004). This condition is undesirable because it affects management factors such as cash flow, labor needs and use of farm machinery.
Methods for mitigating alternate bearing in tree crops include applications of growth regulators to remove fruit clusters or fruit within the clusters, changing developing buds from fruiting to vegetative and decreasing bud abscission. Fruit thinning, branches girdling, manipulating harvest date and nitrogen application also are used (Ludder and Buneman, 1969; Monselise and Goldschmidt, 1982).
Crane and Nelson (1972) suggested that some chemical thinner are ineffective or deleterious. Ferguson and Maranto (1989) demonstrated that 2, 4-dichloro- phenoxy acetic acid prevented bud abscission. Hand thinning and girdling are impractical. Earlier harvesting decreases the percentage of shell splitting, the major quality factor (Ferguson et al., 1995). Nitrogen application, therefore, appears to be the promising option for controlling alternate bearing of pistachios.
Alternate bearing strongly influences tree demand for nitrogen, as indicated by the severely reduced nutrient content in perennial tissues of mature fruit trees following heavy fruiting (Golomb and Goldschmidt, 1987). Weinbaum et al. (1994 a, b) have provided evidence that in on-year there is a strong reproductive demand for nitrogen. They reported that the greatest decrease in leaflet nitrogen concentration and total leaflet nitrogen content per tree during the period from pre-seed fill to fruit maturation.
This study was undertaken to determine the effect of application date and various concentration of nitrogen on flower bud retention of pistachio trees.
MATERIALS AND METHODS
The experiment was carried out with 30-year-old “ON” year ‘Owhadi’ pistachio trees in the research station of the Iran’s Pistachio Research Institute in Kerman region. The experimental design was a 3x3 factorial experiment in a randomized complete block with four replications. Each block consisted of nine trees. Two branches (north and south of each tree) with similar fruits and flower buds were used for each treatment. Branches were hand sprayed to the point of run-off with nitrogen compounds on three different dates.
Treatments were control, 0.3 and 0.6% nitrogen as urea. Tween 20 was used as a wetting agent. The three different application dates were May 6, June 17 and July 18. These dates were selected to coincide with flower bud initiation, at the beginning and middle of filling kernel, respectively. At harvest time, bud retention (%), leaf chlorophyll content, leaf photosynthesis and leaf area were measured.
The leaves chlorophyll content of leaves was determined non-destructively using a portable chlorophyll meter (SPAD-502, Minolta, Japan). Chlorophyll content was calculated as described by Higuchi et al. (1999). Six readings were taken per leaf and then averaged.
Photosynthesis rates were determined with a portable infrared gas analyzer (LCA-4, from Analytical Development Company (ADC), Hoddesdon, UK) and a Parkinson leaf Chamber type PLC4. The top leaflet of each leaf was enclosed in the chamber and exposed to sun light at ambient temperature and humidity. Measurements were taken between 9:00 and 11:00 am, at photosynthetic flux density above 1600 μmole m-2 s-1.
Leaf area was measured using a portable leaf area meter (AM 200, analytical Development Company (ADC), Hoddesdon, UK).
After measuring the above parameters, samples including fruits, current shoots, flower buds and leaves were collected and were kept in an ice box until brought into the laboratory. Weight of 100 nuts, blankness (%) and dehisced nuts (%) were recorded for each sample. Total nitrogen (% dry weight) was determined using a Kjeltec 2300 Analyzer unit (Foss Tecator, Sweden). Data were analyzed for significant differences using a factorial analysis of variance with applications date and nitrogen concentration as main factors by Least Significant Difference Test (LSD). Relationships between characters were described using linear regression analyses where appropriate. Data presented as percentage, were analyzed after appropriate statistical transformation.
RESULTS
Results showed that nitrogen applied on June 17 caused 19.07 and 41.09% more bud retention than when applied on May 6 or July 18, respectively (Table 1). Both concentrations of nitrogen increased the percentage of bud retention per shoot. However 0.3% nitrogen was more effective (Table 1). There was a significant interaction (p = 0.05) between the date of application and nitrogen concentrations, there was an increase in the bud retention when using 0.3% nitrogen applied on June 17 (Table 1).
The amount of nitrogen in leaves, current shoots and flower buds was also affected by nitrogen application. Both concentrations of nitrogen increased nitrogen content in leaves, current shoots and flower buds as compared with control (Table 2-4). Nitrogen concentration of leaves, current shoots and buds was not affected by application date. However, a significant interaction occurred between nitrogen concentration and the application time, there was an extra increase using 0.6% nitrogen on June 17 (Table 2-4).
Time of application had no effect on leaf photosynthesis (Table 5). Both concentrations of nitrogen increased leaf photosynthesis, with the highest effect at 0.6% nitrogen. There was an interaction between the application date and nitrogen concentrations; a greater increase occurred in leaf photosynthesis when using 0.3% nitrogen on June 17 and 0.6% nitrogen on July 18 (11.52 and 11.46 μmol m-2 s-1, respectively) (Table 5).
The leaf chlorophyll content was increased significantly by application of nitrogen (Table 6). Trees treated with 0.6% nitrogen showed the greatest leaf chlorophyll content (1.69 mg g-1 d.w.). The nitrogen applied on July 18 gave the highest leaf chlorophyll content compared with other dates. However, the interactions between nitrogen concentration and application date showed greater leaf chlorophyll content when 0.6% nitrogen was applied on July (Table 6).
Leaf area was also affected by both nitrogen concentration and application date. Nitrogen application significantly increased leaf area, with the largest effect with 0.3% nitrogen (Table 7). Trees treated on May 6 showed the largest leaf area (87.53 cm2). There was a significant interaction between the application date and nitrogen concentration. The largest increase in leaf area occurred when using 0.6% nitrogen on May 6 (Table 7).
In this study, nut weight, blankness and dehiscence percentage of pistachio were not affected by application date or nitrogen treatments (data not presented).
| Table 1: | Effects of application date and nitrogen concentrations on flower bud retention (%) of pistachio trees |
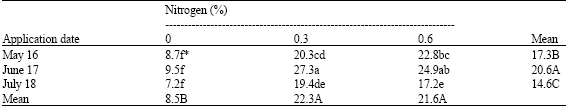 | |
| * Mean separation within columns and rows by LSD, at 5% level | |
| Table 2: | Effects of application date and nitrogen concentrations on nitrogen level (%) in leaves of pistachio trees |
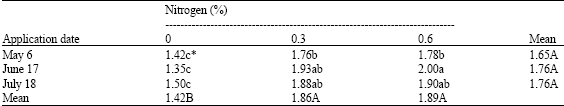 | |
| * Mean separation within columns and rows by LSD, at 5% level | |
| Table 3: | Effects of application date and nitrogen concentrations on nitrogen level (%) in current shoots of pistachio trees |
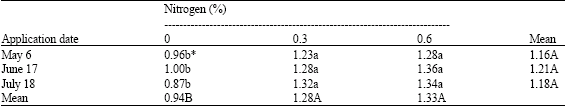 | |
| * Mean separation within columns and rows by LSD, at 5% level | |
| Table 4: | Effects of application date and nitrogen concentrations on nitrogen level (%) in buds of pistachio trees |
 | |
| * Mean separation within columns and rows by LSD, at 5% level | |
| Table 5: | Effects of application date and nitrogen concentrations on leaf photosynthesis (μmol m-2 s-1) of pistachio trees |
 | |
| * Mean separation within columns and rows by LSD, at 5% level | |
| Table 6: | Effects of application date and nitrogen concentrations on leaf chlorophyll content (mg g-1 d.w) of pistachio trees |
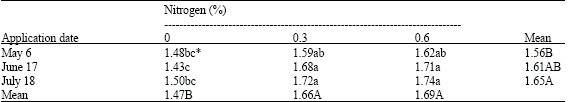 | |
| * Mean separation within columns and rows by LSD, at 5% level | |
| Table 7: | Effects of application date and nitrogen concentrations on leaf area (cm2) of pistachio trees |
 | |
| * Mean separation within columns and rows by LSD, at 5% level | |
DISCUSSION
Our data showed that nitrogen application decreased flower bud abscission in pistachio. In agreement with our results, Lovatt and Ferguson (1994) also showed that foliar application of urea combined with cytokinine decreased flower bud abscission in ‘Kerman’ pistachio trees. In contrast, Ferguson (1986) reported that foliar urea and KNO3 sprays in early spring had no effect on flower bud abscission. However, in the present study, nitrogen application in the first and last date had lower effects on flower bud retention.
Most pistachio flower buds abscise during the period of rapid seed development (Crane and Iwakiri, 1987), suggesting competition between flower buds and developing nuts for metabolites, especially nitrogen (Wolpert and Ferguson, 1990) and carbohydrates (Vemmos, 1999). Durzan (1996) also reported that the total nitrogen of strongly adhering flower buds consistently had 15% higher level of total nitrogen compared to abscising flower buds at the end of growth season. Seed growth is characterized by a rapid increase in nitrogen content and the ability of seed to mobilize nitrogenous substances from other parts of the plant is evident from the decrease of nitrogen in the flower buds (Porlingis, 1974). On the other hand, high levels of prolin in apoptotic flower buds indicated that buds were under physiological stress due to competition for food supply (Durzan, 1996). The low concentrations of nitrogen in plant tissues may cause a decrease in polyamines (Marschner, 1995). Polyamines have been reported to serve either as nitrogenous source or as signal molecules regulating the fruitlet abscission process in grapevine (Aziz, 2003). Roussos et al. (2004) reported that flower buds derived from non-fruiting shoot of pistachio trees had higher polyamines than buds from fruiting ones during the rapid seed growth. Therefore, polyamines could play a crucial role in the flower bud abscission of pistachio. Polyamines are known to act as anti-senescence agents and counteract the activity of abscisic acid and ethylene (Evans and Malmberg, 1989; Chen et al., 2002).
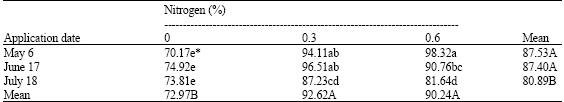
The results of this experiment indicate that foliar treatment of nitrogen results in increased nitrogen concentration in leaves, current shoot and flower buds. A similar response was observed by Felix Ponder and Jones (2001) for black walnut and by Darke et al. (2002) for apple. A positive correlation was obtained between the bud retention with nitrogen concentration in leave (r = 0.80), current shoot (r = 0.73) and flower bud (r = 0.70) (Fig. 1). Correlation between bud retention and nitrogen in leaves has been reported for ‘Kerman’ pistachio trees (Wolpert and Ferguson, 1990).
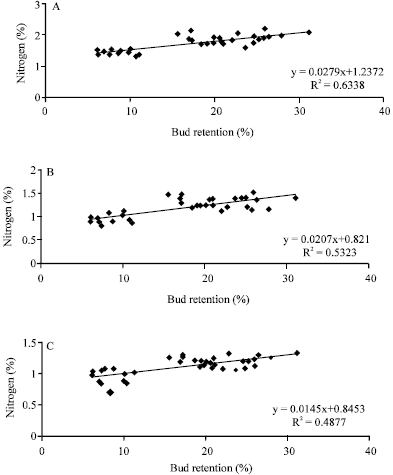
The results presented here showed a correlation (r = 0.55) between leaf nitrogen concentration and leaf photosynthesis (Fig. 2A). The correlation between leaf nitrogen content and leaf photosynthetic potential reported for other fruit tree species may have several ramifications regarding research on plant photosynthesis. If one accepts the concept that photosynthesis is the primary function of green leaves, then the association of leaf nitrogen content with the ability to carry out that function may indicate that leaf nitrogen content represents a primary “Cost” in construction and maintenance of the photosynthetic apparatus.
 | |
| Fig. 1: | Correlation of flower bud retention with leaf nitrogen (A), current shoot nitrogen (B) and flower bud nitrogen (C) in pistachio trees |
 | |
| Fig. 2: | Correlation of nitrogen on leaf with (A) photosynthesis (A) and (B) chlorophyll concentration (Chl.) in pistachio leaves |
 | |
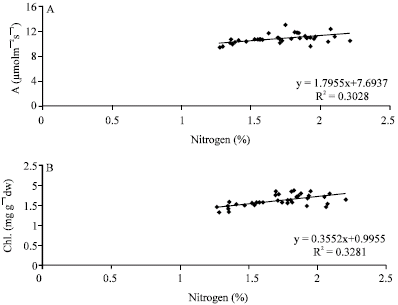
| Fig. 3: | Correlation of flower bud retention with leaf area in pistachio trees |
Indeed, this appears likely, since the primary carboxylating enzyme in leaves, ribulose bishopsphate carboxylase, make up a substantial fraction of the total leaf protein (Dejong, 1982; Chen and Cheng, 2003).
Leaf chlorophyll concentration increased linearly with leaf nitrogen concentration (Fig. 2B). It was similar to the results obtained by Bondada and Syvertsen (2003) in citrus. However, our results implied that nitrogen enhanced chlorophyll synthesis. Because chlorophyll is embedded in the thylakoid membrane, increased chlorophyll synthesis would result in the expansion of thylakoid membrane assembly mediated by increase in the number of grana and thylakoids per granum (Bondada and Syvertsen, 2003).
Nitrogen application increased leaf area in pistachio trees. This effect of nitrogen on leaf was consistent with the effect previously found with kiwifruit (Costa et al., 1997). Thompson et al. (1988) reported that, high nitrogen supply could enhance cell enlargement and thus favor the production of larger leaves. The results presented here showed a positive correlation (r = 0.87) between bud retention and leaf area of ‘Owhadi’ pistachio (Fig. 3). Crane et al. (1973) also showed that, abscission of flower buds increased progressively as leaf area decreased.
The response of pistachio trees to nitrogen treatments outlined in this study suggests that these techniques have a potential application for increasing the flower bud retention. In summary, using nitrogen at 0.3% at the June 17 is most effective for bud retention in pistachio trees.
ACKNOWLEDGMENTS
We greatly appreciate the help from the Iranian Pistachio Research Institute. We also thank Mr. H. Alipour, M.A. Ershadi, R. Shahidi and M. Sadre-Arhami for their valuable help with this experiment.
REFERENCES
- Aziz, A., 2003. Spermidine and related-metabolic inhibitors modulate sugar and amino acid levels in Vitis vinifera L.: Possible relationships with initial fruitlet abscission. J. Exp. Bot., 54: 355-363.
Direct Link - Bondada, B.R. and J.P. Syvertsen, 2003. Leaf chlorophyll, net gas exchange and chloroplast ultrastructure in citrus leaves of different nitrogen status. Tree Physiol., 23: 553-560.
Direct Link - Chen, S., S. Wang, A. Huttermann and N. Altman, 2002. Xylem abscisic acid accelerates leaf abscission by modulating polyamine and ethylene synthesis in water stressed intact poplar. Trees, 16: 16-22.
Direct Link - Chen, L.S. and L. Cheng, 2003. Carbon assimilation and carbohydrate metabolism of concord grape (Vitis vinifera L.) leaves in response to nitrogen supply. J. Am. Soc. Hortic. Sci., 128: 754-760.
Direct Link - Costa, G., O. Lain, G. Vizzotto and G. Johnson, 1997. Effect of nitrogen fertilization on fruiting and vegetative performance, fruit quality and post-harvest life of kiwifruit cv. Hayward. Acta Hortic., 444: 279-284.
Direct Link - Drake, S.R., J.T. Raese and T.J. Smith, 2002. Time of nitrogen application and its influence on Golden Delicious apple yield and fruit quality. J. Plant Nutr., 25: 143-157.
Direct Link - Durzan, D.J., 1996. Nitrogen reallocation during female bud abscission in alternate bearing of pistachio. Adv. Hortic. Sci., 10: 219-225.
Direct Link - Evans, P.T. and R.L. Malmberg, 1989. Do polyamines have roles in plant development? Annu. Rev. Plant Physiol. Plant Mol. Biol., 40: 235-269.
CrossRefDirect Link - Felix Ponder, J. and J.E. Jones, 2001. Annual applications of N, P and K interrupt alternate- year nut crops in black walnut. J. Plant Nutr., 24: 661-669.
Direct Link - Ferguson, L., J. Moranto and R. Beede, 1995. Mechanical topping mitigates alternate bearing of Kerman pistachio (Pistacia vera L.). HortScience, 30: 1369-1372.
Direct Link - Higuchi, H., T. Sakuratani and N. Utsunimiya, 1999. Photosynthesis, leaf morphology and shoot growth as affected by temperatures in cherimoya (Annona cherimola Mill.) trees. Scientia Hortic., 80: 91-104.
Direct Link - Roussos, P.T., C.A. Pontikis and M.A. Zoti, 2004. The role of free polyamines in the alternate bearing of pistachio (Pistacia vera cv. Pontikis). Trees, 18: 61-69.
Direct Link - Vemmos, N., C.A. Pontikis and A.P. Tolia-Marioli, 1994. Respiration rate and ethylene production in inflorescence buds of pistachio in relation to alternate bearing. Scientia Hortic., 57: 165-172.
Direct Link - Vemmos, S.N., 1999. Carbohydrate content of inflorescent buds of defruited and fruiting pistachio (Pistacia vera L.) branches in relation to biennial bearing. J. Hortic. Sci. Biol., 74: 94-100.
Direct Link - Weinbaum, S.A., G.A. Picchioni, T.T. Muraoka, L. Ferguson and P.H. Brown, 1994. Fertilizer nitrogen and boron uptake, storage and allocation vary during the alternate bearing cycle in pistachio trees. J. Jap. Soc. Hortic. Sci., 119: 24-31.
Direct Link - Wolpert, J.A. and L. Ferguson, 1990. Inflorescence bud retention in 'Kerman' pistachio: Effect of defruiting date and branch size. Hortic. Sci., 25: 919-921.
Direct Link