
Nishi Mathur
Department of Botany, Microbial Biotechnology and Biofertilizer Laboratory,
J.N.V. University, Jodhpur, India-342 003
Joginder Singh
Department of Botany, Microbial Biotechnology and Biofertilizer Laboratory,
J.N.V. University, Jodhpur, India-342 003
Sachendra Bohra
Department of Botany, Microbial Biotechnology and Biofertilizer Laboratory,
J.N.V. University, Jodhpur, India-342 003
Anil Vyas
Department of Botany, Microbial Biotechnology and Biofertilizer Laboratory,
J.N.V. University, Jodhpur, India-342 003
International Journal of Soil Science
Year: 2007 | Volume: 2 | Issue: 2 | Page No.: 119-127







ABSTRACT
A field study of 8 saline districts processing salt lakes and salt basins in arid zones of Rajasthan was undertaken to evaluate the occurrence of three selected halophytic plant species (Suaeda fruticosa (Linn.) Forsk, Salsola baryosma (Roem. et Schult) Dandy, Haloxylon recurvum (Moq.) Bunge ex Boiss) and Arbuscular Mycorrhizal Fungal (AMF) associations with them. Five genera were identified in the rhizosphere of these selected plant species. A high diversity of AMF was observed which varied between different host plant species. Among the five genera, Glomus occurred most frequently, with eight species, Acaulospora, Gigaspora and Scutellospora were found with three species, respectively, while Sclerocystis was detected with two species each. Gigaspora margarita, Gigaspora rosea, Glomus deserticola, Glomus ambisporum, Glomus fasciculatum, Glomus mosseae and Scutellospora calospora were the most dominant species. The AMF spore density was not clearly affected by the host plant suggesting that biotic factors may be relatively less important than abiotic/edaphic factors for establishing population pattern. The spore density of AMF had a strong positive correlation with soil pH and organic carbon content and a negative correlation with Olsen's P content of the soil. The association with AMF of these plant species native to the harsh environmental conditions of the Indian Thar Desert may play a significant role in the re-establishment and conservation of these multipurpose halophytic plants.
PDF Abstract XML References
How to cite this article
Nishi Mathur, Joginder Singh, Sachendra Bohra and Anil Vyas, 2007. Arbuscular Mycorrhizal Status of Medicinal Halophytes in Saline Areas of Indian Thar Desert. International Journal of Soil Science, 2: 119-127.
DOI: 10.3923/ijss.2007.119.127
URL: https://scialert.net/abstract/?doi=ijss.2007.119.127
DOI: 10.3923/ijss.2007.119.127
URL: https://scialert.net/abstract/?doi=ijss.2007.119.127
INTRODUCTION
Suaeda fruticosa (Linn.) Forsk, Salsola baryosma (Roem. et Schult) Dandy, Haloxylon recurvum (Moq.) Bunge ex Boiss are important halophytic plant species (Chenopodiaceae, shrub) of the saline areas of Indian Thar Desert (Table 1). Huge demand and multipurpose uses of these plants in pharmaceutical industries, population bloom, urbanization, over exploitation and recurring drought and famine in this region make these plant species valuable (Bhandari, 1990; Shetty and Singh, 1991). All these plant species are found in arid regions of Thar desert, which are characterized by poor sandy soils, low organic matter, uncertain and erratic rainfall, high wind velocity and generally experience water deficit during the growth period.
Salinity affects the establishment, growth and development of plants in billions of hectares of earth lands. These alter physio-chemical and metabolic activities of host plant and it is a world wide problem (Giri and Chamola, 2000). In arid and semi-arid areas concentration of salts and availability of water is enough to damage the growth of plant. Major areas of Indian Thar Desert are represented by saline belts of Luni, Pachpadra Pokaran, Sambhar, Baap, Phalodi etc. Large area of this land are either barren or are represented sparse vegetation. Arbuscular mycorrhizal fungi (AMF) are a major component of rhizosphere microflora in natural ecosystems, and play significant role in the re-establishment of nutrient cycling in native ecosystems (Peterson et al., 1985).
| Table 1: | Important uses/medicinal values of the three selected halophytic plants |
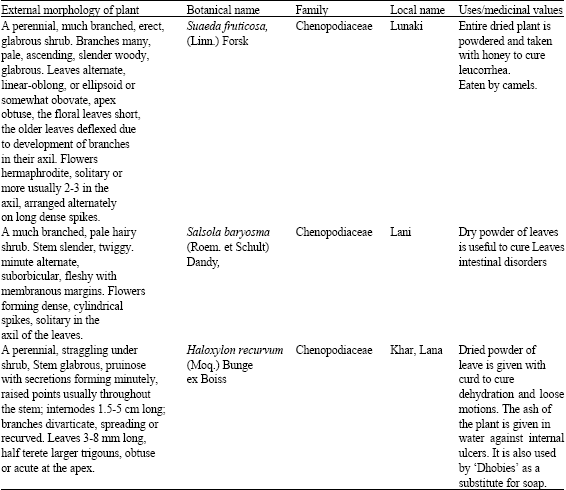 | |
AMF associations are ubiquitous and play an important role in ecosystem diversity. They can modify the structure and function of plant communities (Giovannetti and Gianinazzi-Pearson, 1994; Douds and Millner, 1999) and may be useful as indicators of ecosystem change (McGonigle and Miller, 1996). Studies on the distribution and activity of AMF can help elucidate the ecological significance of AMF associations (Sanders, 1990). The population of AMF varies greatly and their distribution is affected by various biotic and abiotic factors (Mohammad et al., 2003). Preliminary studies have indicated that AMF are very common in arid soils and form associations with most of the plants growing in Indian desert (Kiran Bala et al., 1989; Pande et al., 1999). Hayman (1983) reported better establishment of vegetation in arid areas by using AMF as these fungi may/often enhance plant absorption of P and other elements, improve water uptake and its transport to plants and enable the plants to withstand high temperatures. Further, Panwar and Vyas (2002) indicated the significance of AMF in re-establishment and conservation of endangered plants in arid areas. Turnau and Haselwandter (2002) also considered AMF as a tool for re-establishment of endangered plant species. In general, knowledge of the mycorrhization of halophytic plant species is lacking. On the other hand, information of this kind must be considered as a pre-requisite for making plant re-establishment programmes successful (Van der Heijden et al., 1998).
Prior to exploiting the biofertilizers potential of AMF in relation to these halophytic plant species, it is necessary to examine the spatial distribution and colonization of these microbes in soil, since AMF species vary with ecosystems (McGonigle and Miller, 1996) and are affected by edaphic factors (Sanders, 1990).
Keeping these facts in mind, an extensive field investigation was carried out to evaluate spatial distribution and colonization of AMF species present in the rhizosphere of three selected halophytic plants and to study effects of edaphic factors on AMF populations in the rhizosphere.
MATERIALS AND METHODS
Site Description
The Indian Thar Desert comprises about 70% part of the Western Rajasthan, incorporating 8 saline districts processing salt lakes and salt basins viz., Barmer (Pachpadra, Thob and Sanwarla), Bikaner Lunkaransar and Khajuwala), Churu (Tal Chappar), Jaisalmer (Lanela and Pokaran), Jodhpur (Bap and Phalodi), Nagaur (Didwana), Pali (Luni) and Sri Ganganagar (Sakhi). An intensive field survey of these 8 districts was undertaken in order to find out occurrence of selected halophytic plant species i.e., S. fruticosa, S. barysoma, H. recurvum and AMF associations with them. Important climatological characteristics of surveyed districts are summarized in Table 2.
Soil Sampling
Rhizosphere soil samples (soil adhering to the roots) were collected at 30-90 cm depths along with root samples in five replicates from each plant. Before sampling, the soils from the upper layer were scrapped off to remove foreign particles and litter. The collected soil and root samples were placed in an insulated carrier for transport and immediately refrigerated at 4°C upon arrival. The roots were processed immediately. All the soil samples collected from the rhizosphere of a particular plant species of a district were homogenized replication wise before processing by sieving (<2 mm mesh size) to remove stones, plant material and coarse roots. Subsample of each soil was air dried and used for estimation of various physico-chemical properties and to establish successive pot cultures (trap cultures).
Trap Cultures
Successive pot cultures (trap cultures) have been shown to be a useful tool in inducing sporulation of AMF from field soils in arid ecosystems to facilitate the detection of AMF species that are present in the rhizosphere and roots but do not sporulate readily in the field at the time of sampling (Stutz and Morton, 1996). To establish successive pot cultures, 500 g dry wt. field soil was mixed with autoclaved sand (1:1, v/v) and planted with surface-sterilized seeds (by 0.1% w/w mercuric chloride solution for 2 min and then washed with distilled water) of Cenchrus ciliaris L. as host.
| Table 2: | Important site characteristics of surveyed districts in Western Rajasthan (India) |
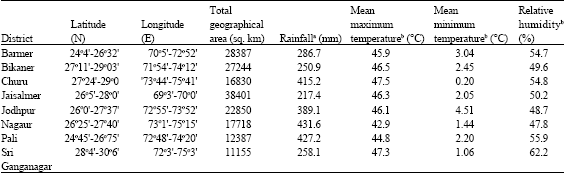 | |
| aAverage of last 10 years. bAverage of last 5 years | |
Root Colonization by AMP
To determine the percent root colonization, root samples collected from different sites were washed in tap water and staining was done by the method of Phillips and Hayman (1970) for rapid assay of mycorrhizal association. The root samples were cut into pieces of 1 cm length and placed in 10% KOH solution, which was kept at boiling point for about 10 min (depending upon the hardness of the root sample). The root samples were captured on a fine sieve and rinsed with distilled water until the brown colour disappeared. Post-clearing bleaching was done with alkaline hydrogen peroxide (0.5% NH4OH and 0.5% H2O2 v/v in distilled water). Roots were rinsed with distilled water, treated with 1% HCl and stained with 0.05% w/v trypan blue in lactic acid-glycerol. Assessment of colonization was conducted on each sample by the glass slide method, in which 100 randomly selected root segments of each replication were determined microscopically. A segment was counted as infected when hyphae, vesicles, or arbuscules were observed. The infection percentage was determined by the method given by Biermann and Lindermann (1981).
Spore Extraction
Spores of AMF were extracted from the field and successive pot culture soils by the wet sieving and decanting technique of Gerdemann and Nicolson (1963). Total spore numbers of mycorrhizal fungi in the soil samples were estimated by the method of Gaur and Adholeya (1994) and spore densities were expressed as the number of spores per 100 g of soil. The isolated spores were picked up with needle under a dissecting microscope and were mounted in polyvinyl lactoglycerol (PVLG). However, PVLG was mixed with Meltzer's reagent (1: 1, v/v) in case of Scutellospora species. All the spores (including broken ones) were examined using Medilux-20 TR compound microscope. Taxonomic identification of spores up to species level was based on spore size, spore colour, wall layers and hyphal attachments using the identification manual of Schenck and Perez (1990) and the description provided by the International collection of vesicular and AMF (http://invam.caf. wvu.edu).
Soil Parameters
Soil samples were analysed for pH and electrical conductivity on 1: 2.5, soil: water suspension. Organic carbon was estimated by the method of Walkley and Black (1934) using 1 N potassium dichromate and back titrated with 0.5 N ferrous ammonium sulphate solution. Available phosphorus in soil was determined by extraction with 0.5 M sodium bicarbonate for 30 min (Olsen et al., 1954). Soil texture was estimated gravimetrically by hydrometer method (Jackson, 1967).
Microsoft Excel 2000 was used in the statistical processing of the data (Standard errors of mean. correlation analysis).
RESULTS AND DISCUSSION
During the survey of entire 8 districts (174972 sq. km) it was found that these three halophytic plant species occurred at all the districts. Physicochemical properties of the soils of each site are presented in Table 3. Soil texture varies from sandy gravel to clay loam. The soil had a pH ranged from 8.10 to 8.42, organic carbon between 0.20 and 0.40% and Olsen P level of 4.2-7.5 mg kg-1. In general, soils are alkaline in reaction, low in organic matter content and available P status.
Nineteen species of AMF were identified in the rhizosphere soils collected from field and successive pot cultures scattered over five genera viz., Acaulospora, Gigaspora, Glomus, Sclerocystis and Scutellospora (Table 4). Glomus species were most dominant and made up for more than 50% of the total isolates followed by Acaulospora (3 species), Scutellospora (3 species), Gigaspora (3 species) and Sclerocystis (2 species). It is evident that the occurrence of various species of AMF varied considerably with different plant species.
| Table 3: | Physicochemical characteristics of site soils for halophytic plants |
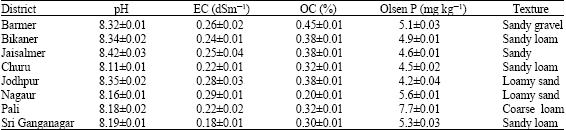 | |
| ±Standard error of mean | |
| Table 4: | Distribution of AMF species associated with different halophytic plants |
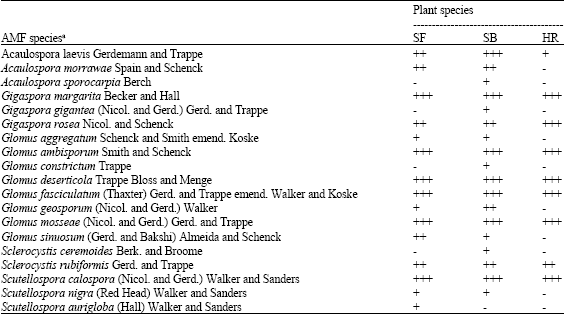 | |
| - = Absent, + = Low (< 20%), ++ = Moderate (20-50%), +++ = High (>50%), SF = Suaeda fruticosa, SB = Salsola baryosma, H = Haloxylon recurvum. aGiven species were found in at least one site | |
S. baryosma colonized with 18 of the total species found associated with selected plants followed by S. fruticosa (15) and H. recurvum (9). This reveals a high specific consortium to each rhizosphere with a high degree of variance in species composition. Hence, a very high AMF diversity index in Thar Desert soils was apparent. Gigaspora margarita, Gigaspora rosea, Glomus ambisporum, Glomus deserticola, Glomus fasciculatum, Glomus mosseae and Scutellospora calospora were the most dominant species (Table 4). Glomus is to be the most abundant of all AMF genera under arid environment (Tarafdar and Kumar, 1996), which may be due to its resistance to high soil temperature (Al-Raddad, 1993).
The density of viable AMF spores recovered from the rhizosphere soil samples collected from field and successive pot cultures were ranged between 84 and 147 spores 100 g-1 soil for studied plants (Table 5). However, a considerable variation was found at different study sites (Table 6). The spore density is relatively low, which is common for arid and semi-arid lands (Requena et al., 1996). It is evident from the data that maximum spore densities were recorded for S. baryosma, followed by S. fruticosa and H. recurvum. These findings agree with that of Al-Raddad (1993), who attributes these differences to the length of the growing season and the type of root systems of trees, which make the rhizosphere more favourable to spore propagation and AMF colonization. It is clear from the results (Table 5) that the rhizosphere soils collected from field and successive pot cultures in Jodhpur have higher AMF spore densities compared to other sites. This may be because of poor soil fertility (in terms of available phosphorus) which results in higher AMF populations (Norani, 1996).
| Table 5: | Viable AMF spore population (100 g-1 soil) and percent root colonization in rhizosphere or three halophytic plants at different sites in Western Rajasthan (India) |
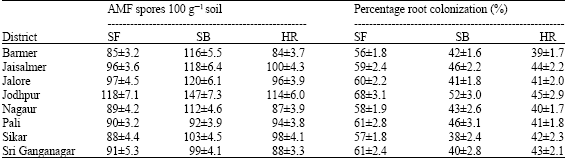 | |
| ±Standard error of mean, SF = Suaeda fruticosa, SB = Salsola baryosma, H = Haloxylon recurvum | |
| Table 6: | Spatial distribution of different AMF spores in rhizosphere soils of three halophytic plants |
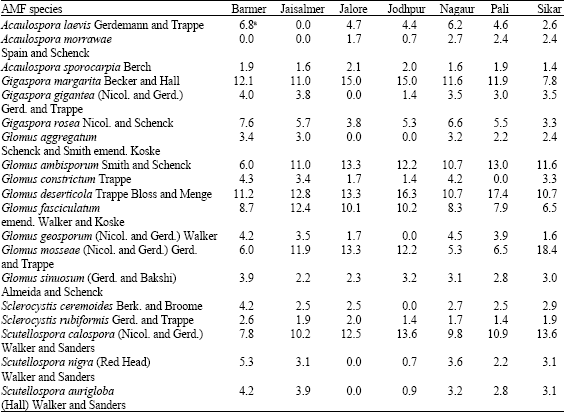 | |
| aPercent values of total spores | |
Natural AMF colonization of root samples varied between 38 and 68% (Table 5). Cleared and stained roots showed the presence of globose to subglobose or ellipsoid bodies (vesicles or spores), dichotomously branched structures (arbuscules) and hyphae in all the sites. Extrametrical hyphae bearing resting spores were also seen associated with the roots of selected plants during study. Considerable variation in percent root colonization and number of different AMF spores associated with plant rhizosphere was observed but no definite correlation could be established between them (Kalita et al., 2002). However, contradictory results were reported by Mutabaruka et al. (2002), as a significant positive correlation and by Louis and Lim (1987), as a negative correlation between percent root colonization and AMF spores.
Table 7 reveals correlation analysis between AMF spore population and different edaphoclimatic factors. It is evident from the results that AMF spore populations were affected by soil pH, organic carbon and Olsen P content. A significant positive correlation with pH (r = 0.85, p<0.01) and organic carbon (r = 0.68, p<0.05) was recorded during present investigation.
| Table 7: | Relationship between AMF spore population and different edaphic factors |
 | |
| *p<0.05; **p<0.01; n = 12 | |
In contrast, a strong negative correlation was observed with soil Olsen P content (r = -0.86, p<0.01). Blaszkowski (1993) while investigating plant communities in Poland, observed a significant positive correlation between AMF spore density and soil pH. A positive correlation with organic carbon content in soil coincide with the findings of Mohammad et al. (2003), who reported the same while investigating under semi-arid environment of Jordan. Organic matter content in the soil increases the water-holding capacity of the soil (Brady and Weil, 1996) and, therefore, may facilitate a more favourable soil moisture condition for the AMF population. When plants have high nutrient availability (especially phosphorus), a negative response and low AMF spore population should be expected (Dhillon and Zak, 1993).
Our results pioneered to identify the status and occurrence of the selected halophytic plant species in Thar Desert environment as well as AMF diversity with them, indicating the mycorrhizal dependency of these plants. Glomus is considered to be the most common arbuscular mycorrhizal genus in this region. No host plant or geographic location specificity was observed, suggesting the population of AMF species was affected mainly by edaphic factors. Recovery of large AMF diversity with these plants reveals the rich wealth of AMF diversity in harsh environmental conditions in Thar Desert. These native AMF isolates with the capacity to survive under stress conditions may be instrumental in the re-establishment of these halophytic plant species. Thus the present study of mycorrhizal status may well prove crucial for any attempts of re-establishment of such endangered plants. Appropriate strategies can be drawn for the artificial inoculation of one or some of these indigenous AMF, which would make the re-establishment and regeneration attempts ecologically and economically viable in such constrained ecosystems. These approaches will increase our scope to manipulate the symbiosis in conservation schemes.
REFERENCES
- Al-Raddad, A., 1993. Distribution of different Glomus species in rainfed areas in Jordan. Dirasat, 20: 165-182.
Direct Link - Biermann, B. and R.G. Linderman, 1981. Quantifying vesicular-arbuscular mycorrhizae: A proposed method towards standardization. New Phytol., 87: 63-67.
CrossRefDirect Link - Blaszkowski, J., 1993. The occurence of arbuscular fungi and mycorrhizae (Glomales) in plant communities of maritime dunes and shores of Poland. Bull. Polish Acad. Sci. Biol. Sci., 41: 377-392.
Direct Link - Douds Jr., D.D. and P.D. Millner, 1999. Biodiversity of Arbuscular mycorrhizal fungi in agroecosystems. Agric. Ecol. Environ., 47: 77-93.
CrossRefDirect Link - Gerdemann, J.W. and T.H. Nicolson, 1963. Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans. Br. Mycol. Soc., 46: 235-244.
CrossRefDirect Link - Giovannetti, M. and V. Gianinazzi-Pearson, 1994. Biodiversity in arbuscular mycorrhizal fungi. Mycol. Res., 98: 705-715.
Direct Link - Hayman, D.S., 1983. The physiology of vesicular-arbuscular endomycorrhizal symbiosis. Can. J. Bot., 61: 944-963.
Direct Link - Kalita, R.K., D.P. Bora and D. Dutta, 2002. Vesicular arbuscular mycorrhizal association with some native plants. Indian J. For., 25: 143-146.
Direct Link - Louis, I. and G. Lim, 1987. Spore density and root colonization of vesicular-arbuscular mycorrhizas in tropical soil. Trans. Mycol. Soc., 88: 207-212.
CrossRefDirect Link - Mcgonigle, T.P. and M.H. Miller, 1996. Development of fungi below ground in association with plants growing in disturbed and undisturbed soils. Soil Biol. Biochem., 28: 263-269.
CrossRefDirect Link - Muhammad, M.J., S.R. Hamad and H.I. Malkawi, 2003. Population of arbuscular mycorrhizal fungi in semi-arid environment of Jordan as influenced by biotic and abiotic factors. J. Arid. Environ., 53: 409-417.
CrossRefDirect Link - Mutabaruka, R., C. Mutabaruka and I. Fernandez, 2002. Diversity of arbuscular mycorrhizal fungi associated to tree species in semiarid areas of machakos, Kenya. Arid Land Res. Manage., 16: 385-390.
CrossRefDirect Link - Ahmad, N., 1996. An assessment and enumerations of vesiculararbuscular mycorrhiza propagules in some forest sites of Jengka. J. Trop. For. Sci., 9: 137-146.
Direct Link - Olsen, S.R., C.V. Cole, F.S. Watanabe and L.A. Dean, 1954. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate. U.S. Department of Agriculture, Washington, D.C., Pages: 19.
Direct Link - Pande, J., C.M.T. Arafdar and G.N. Gupta, 1999. Vesicular arbuscular mycorrhizal infection in arid zone trees of agroforestry systems. J. Indian Soc. Soil Sci., 47: 54-57.
Direct Link - Panwar, J. and A. Vyas, 2002. AM fungi: A biological approach towards conservation of endangered plants in Thar Desert, India. Curr. Sci., 82: 576-578.
Direct Link - Peterson, R.I., A.E. Ashford and W.G. Allaway, 1985. Vesicular-arbuscular mycorrhizal associations of vascular plants on heron island, a great barrier reef coral Ca. Aus. J. Bot., 33: 669-676.
CrossRefDirect Link - Phillips, J.M. and D.S. Hayman, 1970. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc., 55: 158-161.
CrossRefDirect Link - Requena, N., P. Jeffries and J.M. Barera, 1996. Assessment of natural mycorrhizal potential in a desertified semiarid ecosystem. Applied Environ. Microbiol., 62: 842-847.
Direct Link - Sanders, I.R., 1990. Seasonal patterns of vesicular-arbuscular mycorrhizal occurrence in grasslands. Symbiosis, 9: 315-320.
Direct Link - Stutz, J.C. and J.B. Morton, 1996. Successive pot cultures reveal high species richness of arbuscular mycorrhizal fungi in arid ecosystems. Can. J. Bot., 74: 1883-1889.
Direct Link - Tarafdar, J.C. and P. Kumar, 1996. The role of vesicular arbuscular mycorrhizal fungi on crop, tree and grasses grown in an arid environment. J. Arid Environ., 34: 197-203.
Direct Link - Van-der-Heijden, M.G.A., T. Boller A. Wiemken and I.R. Sanders, 1998. Different arbuscular mycorrhizal fungal species are potential determinants of plant community structure. Ecology, 79: 2082-2091.
Direct Link - Walkley, A. and I.A. Black, 1934. An examination of the degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci., 37: 29-38.
CrossRefDirect Link