
Rufus M. Ubom
Department of Botany, University ofUyo, P.M.B. 101 7, Uyo, Nigeria
International Journal of Soil Science
Year: 2006 | Volume: 1 | Issue: 3 | Page No.: 207-217







ABSTRACT
The influence of soil factors affecting the abundance and distribution of Isoberlinia woodlands in savanna ecosystem was studied. Fourteen woodland plots (ca 62 ha) were sampled systematically for frequency, density, basal area, crown cover and height of woody species ≥3 m high. Three soil pits were excavated in each woodland and soil physical, morphological and chemical properties described and analyzed, for each soil horizon. Results show that the soils are sandy loam to clay loam with dark grayish brown (10 YR 3/2) A-horizons over dark brown (7.5 YR 4/4) to yellowish red (5 YR) 4/6) subsurface horizons; moderate medium subangular blocky peds of friable consistence and contain large amounts of iron concretions. The pH is low with high mobile H+; low capacity for cation absorption and low mineral nutrients. Matrix of PCA scores of soil variables used to establish relationships between vegetation and soil parameters give very high significant values indicating the joint structural patterns between these two systems. Canonical correspondence analysis between the soil parameters (PCA) scores and species data elucidates the distribution and grouping patterns of Isoberlinia and its associes. Thus, variation in species composition in the woodlands reflects microsite differences in soil properties and their ecological amplitudes.
PDF Abstract XML References
How to cite this article
Rufus M. Ubom, 2006. Soil Properties Influencing the Abundance and Distribution of Isoberlina Woodlands in Nigeria. International Journal of Soil Science, 1: 207-217.
DOI: 10.3923/ijss.2006.207.217
URL: https://scialert.net/abstract/?doi=ijss.2006.207.217
DOI: 10.3923/ijss.2006.207.217
URL: https://scialert.net/abstract/?doi=ijss.2006.207.217
INTRODUCTION
Isoberlinia Craib and Stapf is a tree of the family Fabaceae and subfamily Caesalpinioidae. It is used as fuelwood, timber, planks; for shaft lining; as hut poles and the wood ash as soap (Wickens, 1980). It occurs in clusters forming woodlands within the Nigerian Guinea Savanna. These woodlands serve as refuge for (animal) wildlife. The woodlands are sporadically located at discrete positions within the Guinea Savanna, that is, they exhibit zonation in spatial context related to a complex environmental factors (Ubom and Isichei, 1998). The woodlands are associated with quartzite ridges, steep eroding slopes with shallow soils, stony grounds not suitable for farming, laterite platenux and more extensive on higher grounds. Thus, Isoberlinia has the capacity to adapt to certain soil conditions and its woodlands are not found in all areas of Guinea Savanna and never present in any other savanna type in Nigeria. The influence of soil factors in the composition and distribution of savanna vegetation in Nigeria has been reported (Child, 1974, Muoghalu and Isichei, 1991). Others have discussed relationships between soil characteristics and distribution of plant communities in the Guinea savanna (Menaut et al., 1985; Cole, 1986; Sharma, 1988; Abdulhameed et al., 2004). Evidence shows that vegetation associations within the Guinea Savanna and tree species reflect differences in soil texture, structure and mineral contents (Ubom, 1998; Ubom and Isichei, 1998, Abdulhameed, 2005). The aim of this research is to elucidate the complex interrelationships between Isoberlinia woodlands and soils based on measurements of vegetation and soil variables at same woodland sites. The hypothesis is that the occurrence and distribution of Isoberlinia woodlands are influenced by soil characteristics. The basic approach is quantitative involving the use of multivariate approaches. The premise is that vegetation is multivariate viewed in terms of gradients while the soil consists of independently varying properties. Having elucidated the important soil properties that influence the woodland components, the soil system may be viewed as multivariate and a joint structure of interrelationships sought between vegetation and soil systems. The procedures involve the use of Principal Component Analysis (PCA), Canonical Correlation Analysis (COR) and Canonical Correspondence Analysis (CCA). PCA is used to interpret the vegetation and soil structure within their covariance matrices. The appropriateness of this in ecological research is achieved when data are standardized; it requires minimum assumptions and is computationally unambiguous (Greigh-Smith, 1983). COR is used to extract the main directions of vegetation and soil variation when both systems are considered jointly. COR assumes linearity of input variables (Ter Braak and Prentice, 1988), its interpretation is relied upon if the original data matrices are replaced by matrices of eigenvectors. CCA extracts axes of known soil variables by imposing linearity of the variables and relating these to vegetation variation directly. That is CCA focuses on grouping patterns of vegetation composition variation that can be explained by soil variables.
MATERIALS AND METHODS
Study Area
The study area is Kainji Lake National Park located between latitudes 9°45’ and 100°23’ and longitudes 30°40’ and 50°47’E in Northwestern Nigeria (Fig. 1).
The area is in the Northern Guinea Savanna Zone (Keay, 1959). It covers 5340.82 km2 with two non-contiguous sectors. Borgu and Zugurma. The climate of the area has been described by Afolayan (1982) as follows: mean annual rainfall of 100-1200 mm with a peak in July-August. Rain does not occur between November and March. Annual evapotranspiration ranges from 1400 to 1560 mm with rainfall/evapotranspiration ratio ranging from 0.64 to 0.89.
Mean annual relative humidity is 53%. The park is underlain by rocks of he basement complex and Nupe sandstones. An important geomorphological feature is the occurrence of plinthite or ironstone on or close to the surface in most parts (Child, 1974). The topography is thus gently undulating. The Borgu sector is drained by Oil River and its tributaries while the Zugurma sector has few channels and is poorly drained. This study was conducted from October 2002 to October 2005 for two seasons of each year.
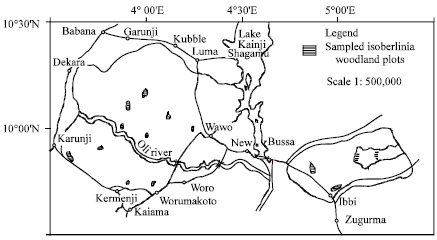 | |
| Fig. 1: | Kainji lake national park: sampled isoberling woodland plots |
Vegetation/soil Sampling and Analyses
Fourteen woodland plots (totaling 62 ha) were used for this study (Fig. 1). In each woodland plot, five belt transects were laid and in each transect, vegetation and soil were systematically sampled (Cochran, 1963) in two hundred and fifty 10x10 m quadrats regularly spaced at 50 m intervals. In each quadrat, frequency and density of all woody species (≥3 m high), basal area as cross sectional area of individuals, crown cover as crown horizontal area were determined; height using Haga altimater was measured. Three soil pits were excavated in each woodland and a total of forty-two of them were excavated. Some soil physical and morphological properties were described in the field. These include colour (using Munsel Soil Colour Charts, 1973 edition), texture, consistence, horizon boundary, concretion and structure according to USDA (1981) Soil Survey Manual. Rooting depth was also determined in each pit. Soil samples were collected from each horizon of each pedon, air-dried and description for colour was again made on them. The soil samples were ground and analyzed for particle size (hydrometer method), pH, exchangeable acidity (Al+3, H+) and cations (Black et al., 1965), organic carbon (Walkey-Black method), available phosphorus (Bray No. 1 method) and total nitrogen (Kjeldahl method) (Juo, 1979).
Data Analysis
The hypothesis was that the distribution of Isoberlinia woodlands was influenced by variation in soil properties. Testing the hypothesis required the use of ordination techniques to analyze the floristic and soil gradients. The first step is indirect gradient analysis, which was used to summarize the main variation in the species and soil data through Principal Component Analysis (PCA). The matrix of PCA scores (varimax rotation) of soil variations was used to establish patterns and joint structure of interrelationships between vegetation and soil parameters using canonical correlation analysis (Gittins, 1985). This analysis utilized the original vegetation data (Aweto, 1981). Secondly, matrix of PCA scores of species data were used to elucidate the distribution and grouping patterns (in relation to soil variables) of 22 species that had importance value >5% from all the woodlands. Importance value was derived as the sum of relative frequency, relative density and relative dominance of a species (Babour et al., 1987). This relationship was explored using the canonical correspondence analysis technique (Ter Braak, 1986).
RESULTS
Vegetation Components
The components of the woodlands and their attributes are given in Table 1. Species with a frequency of less than 10% is not included in the Table 1. Of the 26 woody species recorded, 22 have girth size >5 m in twelve families. The most common species with 50% occurrence include Isoberlinia sp., Burkea africana, Daniellia, oliveri, Detarium microcarpum, Monotes kerstingii and Vitellaria paradoxa. Table 1 shows that the vegetation is dominated by species of the family Fabaceae. There are variations in the vegetation attributes of the woody species. Isoberlinia sp has the highest values of al the attributes except height. Daniellia oliveri is the tallest plant and Gardenia ternifolia the shortest. Acacia hockii has the smallest crown size, girth size and density. The influence of the variations of the species attributes is exemplified in their importance values. Accordingly the first six important woody species (in descending order) in the woodlands are Isoberlinia sp., Burkea africana, Viterllaria paradox, Monotes kerstingii, Daniellia oliveri and Detarium microcarpum.
Soil Physical Properties
The soils of the woodlands have been described as ferruginous tropical soils according to the Soil Map of Africa (D’ Hoore, 1963). Depending on the structure, texture and topographical position the soils are classified as Paleustalfs, Grossarenic and Typic ustipsamments.
| Table 1: | Mean woody species attributes of Isoberlinia woodlands |
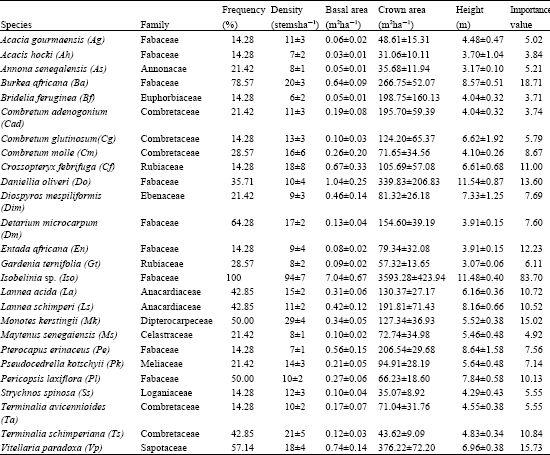 | |
Analysis of the soils reveals that in their physical properties (Table 2) sand is the most variable and silt the least. Sand proportions decrease while those of silt increase with soil depth, clay content is low but more variable than silt and increases with soil depth.
Morphological Properties
The morphological properties of selected pedons studied in the Isoberlinia woodlands are presented in Table 3. The soils are fairly shallow (<150 cm deep), poorly drained with sandy loam A-horizons. The soils have variable colours depicting the drainage status of each of the woodlands. Generally, they have very dark grayish brown (10 YR 3/2) A-horizons over dark brown (7.5 YR 4/4) through dark red (2.5 YR 3/6) to yellowish red (5 YR 4/6) subsurface horizons and moderate medium subangular blocky peds of very friable to friable consistence. The soils contain large amounts of iron concretions. The horizon boundaries are mostly gradual (to subtle). The rooting depth varies from 30 to 85 cm.
Chemical Properties
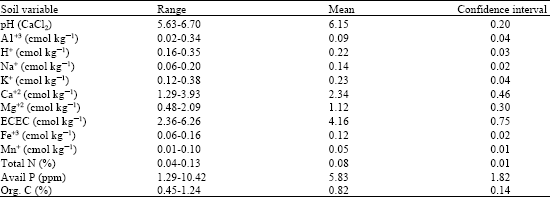
The chemical properties of the soils (Table 4) show that pH values are low, indicating the acidic conditions of the woodland soils. The pH (CaCl2) values decrease as soil depth increases. Thus, A-horizons are less acidic than B-horizons and in some instances tend toward neutrality. High mobility of H+ ions contribute to the acidic nature of soils. The woodland soils also have low capacity for cation absorption and this may be a reflection of low levels of clay which is kaolinitic in nature.
| Table 2: | Mean (+95% confidence interval) physical characteristics of representatives soils of Isoberlinia woodlands in northwestern Nigeria |
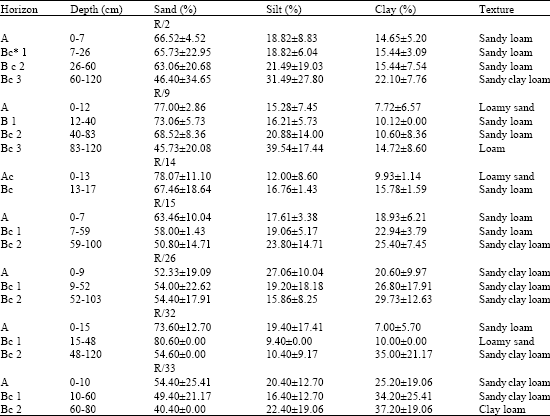 | |
| *c represents concretions and or nodules | |
| Table 3: | *Morphology of selected pedons in representative Isoberlinia woodlands in Nigeria |
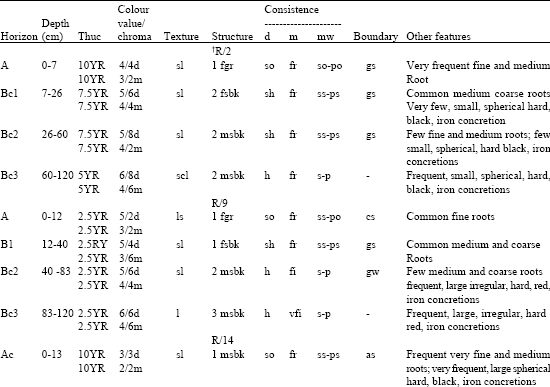 | |
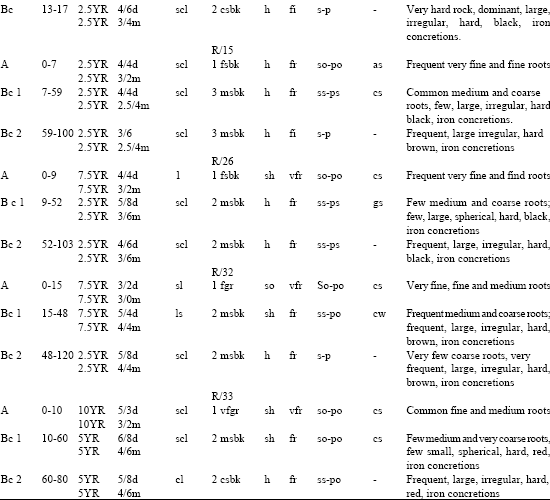 | |
| *Coded according to soil survey staff (1981). Colours are Munsell notations. Codes are as follows: d = dry, m moist, w = wet, Texture: 1 = loam, ls = loamy sand, cl = clay loam, sl = sandy loam, scl = sandy clay loam. Structure: 1 weak, 2 = moderate, 3 = strong, vf = very fine, f = fine, m = medium, o = coarse, gr = granular, Sbk = subangular blocky. Consistence-dry: so = soft, sh = slightly hard, h = hard Moist: vfr = very friable, fr = friable, fi = firm, vfi = very firm Wet: po = non plastic, ps = slightly plastic, p = plastic, so = non sticky, ss = slightly sticky, s = sticky, Boundary: c = clear, g = gradual, s = smooth, w = wavy, a = abrupt. †Pedon identification numbers | |
| Table 4: | Ranges, means and confidence intervals of soil variables in Isoberlinia woodlands, northwestern Nigeria |
 | |
| Table 5: | Canonical factor structure of soil-vegetation interrelationship of Isoberlinia woodlands in Nigeria |
 | |
Thus, the levels of exchangeable cations and micronutrients are low with almost similar variation. The soils also exhibit low levels of total nitrogen, organic carbon and available phosphorus; the later being probably due to its fixation by iron and aluminium because of acidic soil conditions.
Canonical Correlation Analysis
This produced three significant canonical correlations (Table 5). The significance of the correlations represents the patterns of interrelationships between the vegetation and soil characteristics. On the first canonical correlation vegetation attributes of density, crown cover and basal area have high negative weights. The soil attributes of nutrient status and pH have high negative weights while organic carbon, substrate texture and Fe have high positive weights. This correlation indicates a reciprocal relationship between vegetation and soil attributes. All the soil properties are significant in determining vegetation properties. The second correlation indicates a strong positive relationship between the soil nutrient status and woody plant height and basal area, confirming the importance of nutrients in growth of Isoberlinia and its associes. The joint structure on the third correlation indicates a strong positive relationship between soil pH and crown cover as influenced by nutrient status. In all, the most highly weighted soil attributes that influence species distribution and abundance in the woodlands are pH and soil texture.
Canonical Correspondence Analysis (CCA)
Figure 2 shows that the soil properties relate well to the first two species axes of CCA. The first axis is defined by absorption capacity of woodland soils, which differentiates into ECEC probably controlled by Ca+2 silt and iron. The second axis is defined principally by soil pH which differentiates H+ and Al+3; soil texture (sand and clay) as well as organic carbon and nutrient status differentiating into N, P, K, Mg, Mn and Fe components. Figure 2 accounts for 68% of the variance and shows that the weighed averages of the species and soil properties result in similar rankings. Species whose distributions are controlled or affected by say, H+ and Al+3 lie to the top right hand corner of the diagram. Similarly, those species restricted to silt and Na are found on the left hand corner of the diagram. The scores of most species lie outside the centre of the diagram. Only species influenced or restricted by iron and manganese are found around the centre of the diagram. In all, the eigenvalues (not included here) of the soil properties show that the gradients are long, thus indicating that the soil properties have control over the distribution of Isoberlinia sp. and its associes.
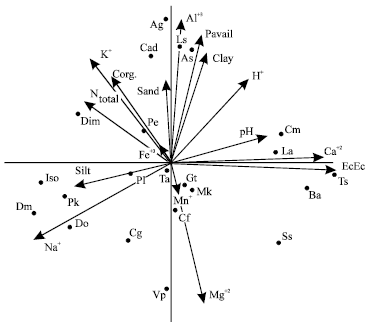 | |
| Fig. 2: | Detrended correspondence analysis (DCA) of soil-species variable of isoberlinia woodlands. The diagram represents 68.01% (Axis I, 35.91%, Axis II, 32.10%) of variance accounted for by the first 4 ordination axes. Species nomenciature as in Table 1 |
DISCUSSION
Variations occur in the distribution of the woodland (species) components which relate the species responses to complex soil variables. Differences in species attributes, which culminate in differences in species importance values, are indications of these responses. Isoberlinia sp. is the most dominant and numerous (with highest importance value) giving a monospecific picture of the woodlands. However, due to less capability of Isoberlinia to regenerate, other competitive species such as Burkea africana, Vitellaria paradoxa, Monotes kerstingii, Terminalia schimperiana, D. microcarpum and Crossopteryx febrifuga establish in the woodlands in gaps created by dead or fallen individuals of Isoberlinia species. This culminates in the variation of species density and other measuration data obtained from the woodlands, giving the woodlands the variable structural patterns and physiognomy. Variation in species composition may also be attributable to microsite differences in soil properties and wide ecological amplitudes possessed by some species such as Burkea africana and Detarium microcarpum (Ubom, 1998); their distribution to competition for environmental factors (Kernel, 1986). The trees, through less dense, have canopy that depicts a continuous upper storey. The understorey vegetation is sparse and of low taxonomic diversity probably as a result of shading by almost closed canopy formed by upper storey species. This suggests that the understorey species are heliosciophytes and that at this instance they are unable to compete with upper storey species.
Surface soils are sandy loam and subsoils sandy clay loam in texture resulting in their poor structural stability, exchange capacity and water-holding capacity. The increase of clay with soil depth is an indication of argilluviation in the soils but is not strong enough to result in argillic horizon (Raji et al., 1996). The soils are nutrient-poor due to their porosity and unabiilty to retain nutrients and organic carbon. Low soil nutrients is attributed to low litter and low mineralization rate, low clay content and moisture in relation to length of wet season (Jones, 1973). Low cation exchange capacity suggests that the soils have low sink for cations and other nutrients and from observation soils of upper slopes have lower nutrient contents than soils of lower slopes.
The variable colour of the woodland soils, the granular surface and subangular blocky sub-surface peds together with very friabe to friable consistence and large amounts of iron concretions are indications of structural of the soils. These suggest that the drainage potential of the woodland soils is related to periodic and seasonal alternating wetting and drying (Siderius, 1974). The woodlands exhibit spatial distribution and show niche relations to these soil attributes (Table 3), suggesting these soil properties could serve as indicators for the woodlands establishment. Their presence on ironstone outcrops where there is low water table suggests that the species components are drought-tolerant. Ironstone outcrops and quartzite ridges offer secondary sites availability to the woodlands as they are drier than slopes, valleys and higher grounds which are the primary sites because they provide more soil moisture to woodlands species. Establishment of woodlands on sites may not only be limited by soil moisture but may imply dependence also on nutrients availability and substrate texture (Ubom, 2005) and competition (Ukpong, 1997). Evidence from observation shows that soils of the upper slopes of ironstone outcrops and quartzite ridges have lower concentrations of nutrients than soils of the lower slopes and consequently woodlands in lower slopes are more luxuriant than those at upper slopes. There is no evidence to show that competitive effects of Isoberlinia with other species are important in woodland establishment since Isoberlinia does not decrease in abundance or incidence in the woodlands. The presence of other species in the woodlands is in gaps if they are canopy species (e.g., Daniellia oliveri) heliosciophytes if understorey species (e.g., Acacia hockii, Entada africana). Hence, temporal variation in species availability may be determined by soil moisture (though not measured in this study) and spatial variation associated with soil properties gradients (Boeken and Shackak, 1998).
The presence of woodlands and their species components on soils with high iron concretions suggests restricted establishment of the woodlands in these specific sites where the species can perform well (Ubom, 1998). Siderius (1974) makes an assertion that Isoberlinia woodlands are located on soils underlain by soft plinthite at 64 cm depth and hard plinthite at about 130cm depth where soils are yellow, red and sandy clay loam in texture. Observations reveal that tall woodlands occupy yellow red sandy clay loam soils underlain by soft plinthite with weak structure while short woodlands are associated with soils underlain by fairly hard plinthite.
The efficacy of Canonical Correlation (COR) and Canonical Correspondence Analysis (CCA) techniques as indirect gradient analysis tools (Ter Braak, 1986) has distinguished patterns of vegetation and species variations in the woodlands. COR shows that the Isoberlinia woodlands is influenced or encouraged by soil texture (silt, clay fractions) and nutrient status. Silt and clay fractions probably enhance growth of woodland species by improving soil water-holding capacity, porosity, cation exchange capacity and play a cementing role between mineral particles. The distribution of the woodlands correlates with organic carbon, soil reaction (pH) and iron contents of the soil. Thus, the vegetation attributes of the woodlands may be determined by these soil parameters, as the latter determine the soil conditions of the woodlands resulting in woodland variation. But Huntley and Birks (1979) assert that vegetation variation is not only related to soil conditions but to aspects of biotic environment such as grazing and burning. Field observations reveal that burning affects woodlands vegetation variation as it removes Isoberlinia saplings and creates gaps for other species (e.g. Burkea africana, Daniethlia oliveri) to establish. The CCA indicates different relationships between species composition of the woodlands and soil properties. This discrepancy suggests that soil properties constitute complex environmental gradients to which the species respond (Ter Braak, 1986). Accordingly, Isoberlinia, Pericopsis, Pseudocedrella and Terminalia are best developed on silt, iron and calcium rich soil sites within the woodlands (Fig. 2). Vitellaria paradoxa is a better indicator of Mg2+ rich soils while Acacia gourmaensis and Annnona senegalensis are associated with Al3+, P and clay rich soils. Spatial segregation of Lannea acida and L. schimperi suggests niche separation between species. The former is probably sensitive to soil reaction (pH) while the latter is sensitive to phosphorus (Ubom, 1998). Lannea acida occurs among short woodlands and L. schimperi occurs among tall woodlands (Ubom and Isichei, 1998). The pattern of segregation of Gardenia ternifolia, Pterocarpus erinaceus and Terminalia avicennioides suggests that their distribution may be monotonic probably due to their insensitivity to changing conditions of the soil properties.
CONCLUSION
This study shows that woody plant species in Isoberlinia woodlands possess qualitative and quantitative response patterns to major axes of habitat hyperspace (Whitney and Moelier, 1982; Ubom, 1998) as influenced by soil properties. Such soil properties include texture, nutrient status, pH, consistence, structure and soil inclusions such as concretions. Their quality and quantity determine the type of vegetation components found within the woodlands and hence, influence their abundance and distribution. Isoberlinia sp. can therefore be described as habitat-specialist (Maliakal-Witt et al., 2005) as its woodlands are restricted to a narrower range of microhabitats within the Guinea Savanna. As a specialist Isoberlinia abundance and distribution are due to high resource but selective requirements in terms of suitability of sites. Since suitable sites are harder to find, its distribution and population are less common within the Guinea Savanna. Hence conservation and management approaches to forestall the disappearance of Isoberlinia and its woodlands and the utilization of the species are recommended.
REFERENCES
- Benson, A.K., 1992. A case study of integrating and modelling seismic, gravity, radar and geotechnical data to identify and characterize shallow geology especially concealed faulting. Proceedings of the 28th Symposium on Engineering Geology and Geotechnical Engineering, 1992, Idaho State University, Rena, NV, pp: 1-17.
- Boadu, F.K., 1997. Fractured rock mass characterization parameters and seismic properties: Analytical studies. J. Applied Geophy., 36: 1-19.
Direct Link - Cardarelli, E. and M. Bernabini, 1997. Determination of parameters of urban waste dumps, two case histories. J. Applied Geophy., 36: 167-174.
Direct Link - Cardarelli, E., C. Marrone and L. Orlando, 2003. Evaluation of tunnel stability using integrated geophysical methods. J. Applied Geophy, 52: 93-102.
CrossRefDirect Link - Davis, J.L. and A.P. Annan, 1989. Ground-Penetrating Radar for high-resolution mapping of soil and rock stratigraphy. Geophys. Prospect., 37: 531-551.
CrossRef - Gilbert, P., 1972. Iterative methods for three-dimensional reconstruction of an object from projections. J. Theor. Biol., 36: 105-117.
PubMedDirect Link - Grandjean, G. and J.C. Gourry, 1996. GPR data processing for 3D fracture mapping in a marble quarry (Thassos, Greece). J. Applied Geophy., 36: 19-30.
CrossRefDirect Link - Leucci, G., S. Negri, M.T. Carrozzo and L. Nuzzo, 2002. Use of ground penetrating radar to map subsurface moisture variations in an urban area. J. Environ. Eng. Geophy., 7: 69-77.
Direct Link - Leucci, G., S. Margiotta and S. Negri, 2003. Geophysical and geological investigations in a karstic environment (salice salentino, lecce, italy). J. Environ. Eng. Geophy., 9: 25-34.
Direct Link - Loke, M.H. and R.D. Barker, 1996. Rapid least-squares inversion of apparent resistivity pseudosections using a quasi-Newton method. Geophys. Prospect., 44: 131-152.
Direct Link - Orlando, L., 2003. Semiquantitative evaluation of massive rock quality using ground penetrating radar. J. Applied Geophy., 52: 1-9.
CrossRefDirect Link - Maliakal-Witt, S., E.S. Menges and J.S. Denshow, 2005. Microhabitat distribution of two Florida scrub endemic plants in comparison to their habitat-generalist congeners. Am. J. Bot., 92: 411-421.
Direct Link - Ter Braak, C.J.F., 1986. Canonical Correspondence Analysis: A New Eigenvector Technique for Multivariate Direct Gradient Analysis. Ecology, 67: 1167-1179.
Direct Link - Ukpong, I.E., 1997. Vegetation and its relation to soil nutrient and salinity in the Calabar mangrove swamp, Nigeria. Mangroves Salt Marshes, 1: 211-218.
CrossRefDirect Link