
Nicole Merkl
Department of Biodiversity and Land Rehabilitation,
Institute of Plant Production and Agroecology in the Tropics and Subtropics, University of Hohenheim, D-70593 Stuttgart, Germany
Rainer Schultze- Kraft
Department of Biodiversity and Land Rehabilitation,
Institute of Plant Production and Agroecology in the Tropics and Subtropics, University of Hohenheim, D-70593 Stuttgart, Germany
International Journal of Soil Science
Year: 2006 | Volume: 1 | Issue: 2 | Page No.: 108-117







ABSTRACT
Phytoremediation of organics is presumed to be based on the stimulation of microorganisms in the rhizosphere, resulting in a higher microbial population and activity and an altered community composition. Substrate utilization patterns of BIOLOG Eco PlatesTM, consisting of 31 carbon sources, were used to determine the effect of the tropical grass Brachiaria brizantha on microbial carbon source utilization patterns in heavy crude oil contaminated soil and to find a possible correlation with enhanced crude oil degradation found in previous experiments. Counts of numbers of carbon sources used only resulted in differences between contaminated and uncontaminated soil while Principal Component Analysis (PCA) showed different carbon source utilization patterns between rhizosphere soil and non-rhizosphere or unplanted soil. Similar to its influence on oil degradation, B. brizantha had a significant effect on microbial community patterns after 14 weeks but only a minor effect after 30 weeks. In particular, D, L-α-glycerol phosphate was more used by microbial communities of the rhizosphere, pointing to a possible key role of phosphorus in the highly nutrient deficient savannah soils of Venezuela and its mobilization in the rhizosphere. Missing incisiveness of the results can be related to the fact that fungi might play a central role in phytoremediation of petroleum hydrocarbons in the tropics but are not detected by the BIOLOG assay.
PDF Abstract XML References
How to cite this article
Nicole Merkl and Rainer Schultze- Kraft, 2006. Influence of the Tropical Grass Brachiaria brizantha
(Hochst. ex A. Rich.) Stapf on Bacterial Community
Structure in Petroleum Contaminated Soils. International Journal of Soil Science, 1: 108-117.
DOI: 10.3923/ijss.2006.108.117
URL: https://scialert.net/abstract/?doi=ijss.2006.108.117
DOI: 10.3923/ijss.2006.108.117
URL: https://scialert.net/abstract/?doi=ijss.2006.108.117
INTRODUCTION
In recent years phytoremediation has received increasing attention from environmental engineers and scientists since this in situ technology is cost-effective and less destructive to the soil environment than abiotic treatments. It uses plants and their associated microorganisms to remove, contain or render harmless environmental contaminants in soil, water and air (Cunningham et al., 1996; Schwab and Banks, 1999).
Numerous authors reported a higher degradation of petroleum hydrocarbons in soil vegetated with e.g., tall fescue, sorghum, cowpea, alfalfa and black rush compared to unvegetated soil (Lin and Mendelsohn, 1998; Wiltse et al., 1998; Schwab and Banks, 1999; Hutchinson et al., 2001). These studies were all conducted with plant species native to temperate zones. In recent studies in a tropical environment, an enhanced degradation of crude oil under the influence of the pasture grass Brachiaria brizantha (Hochst. ex A. Rich.) Stapf was shown (Mager and Hernández-Valencia, 2003; Merkl et al., 2005a, b): planted soil had 10 to 22% less residual petroleum hydrocarbons than an unplanted control after 14 and 26 weeks, respectively. Saturates were reduced up to 50% compared to the control and aromatics up to 24%.
Mechanisms of phytoremediation are still not well understood and mechanistic explanations are probably as complex as the environment in which they take place (Joner and Leyval, 2003). Phytoremediation of organics like petroleum hydrocarbons is commonly presumed to be based on a stimulation of growth and activity of degrading microorganisms in the rhizosphere (Frick et al., 1999) and is therefore also referred to as phytostimulation, rhizodegradation or plant-assisted bioremediation.
Although enhanced microbial growth and activity are probably a key component in rhizodegradation, several authors could not find increased microbial numbers or activity in planted soil, but nevertheless observed superior degradation of contaminants (Pradhan et al., 1998; Kudjo Dzantor et al., 2000; Merkl et al., 2005a, c). Also Fang et al. (2001a) and Joner et al. (2002) observed that the proportion of pollutant degraders remained unchanged in polluted soil receiving root exudates, even when degradation was enhanced. Since modified conditions in the rhizosphere do not proportionally increase the biomass of all its organisms (Joner and Leyval, 2003), the increase in degradation could result from structural and functional changes in the microbial community (Yoshitomi and Shann, 2001). Crude oil consists of a complex mixture of petroleum hydrocarbons and therefore cannot be degraded by a single strain or species (Leahy and Colwell, 1990). Its degradation occurs synergistically by numerous microbial groups, supporting the hypothesis that microbial community composition is as important as microbial population size and activity.
The BIOLOG Eco Plate™ assay used in the present work provides utilization patterns of 31 carbon sources, which are useful in the classification and characterization of heterotrophic microbial communities (Garland and Mills, 1991; Wünsche et al., 1995). For instance, Zak et al. (1994) detected considerable differences between the communities of six soil ecosystems. Haack et al. (1995) distinguished microbial model communities with different compositions by principal component analysis (PCA) of substrate utilization data. BIOLOG also gives a basis for ecologically relevant monitoring of environmental pollution and bioremediation processes (Wünsche et al., 1995) and assessment of functional microbial diversity in the rhizosphere may be useful for plant selection in phytoremediation projects (Banks et al., 1997).
The objective of the present study was to evaluate the influence of B. brizantha on carbon source utilization patterns of contaminated and uncontaminated soil and to detect a possible correlation of changes in microbial community composition with enhanced oil-degradation in planted soil shown in previous experiments (Merkl et al., 2005a, b). This might provide a better understanding of phytoremediation in general and a particular insight into phytoremediation mechanisms under tropical conditions that might differ from what has been defined so far in temperate zones.
MATERIALS AND METHODS
A heavy crude oil contaminated soil planted with B. brizantha, an unplanted contaminated control and a planted uncontaminated soil were set up in 3 replicates (Table 1). Destructive samples were taken after 14, 22 and 30 weeks.
Soil Preparation
An air-dried loamy sand (89%, sand; 6%, silt; 5%, clay; pH 5.9; 0.9%, organic matter; 0.04%, total N; 4 ppm, total P; 21 ppm, K; 81 ppm, Ca; 17 ppm, Mg) from the eastern savannah of Venezuela (Santa Bárbara, Monagas State) was sieved through a 2 mm-screen and thoroughly mixed by hand with 5% (w/w) of a heavy crude oil (Boscán: 10°C, API gravity; 15.6°C, pour point; 5.6%, sulphur; 13.4%, saturates; 40%, aromatics; 46.6%, polars).
| Table 1: | Experimental set-up and codes of treatments and compartment samples |
 | |
| Table 2: | Carbon sources of BIOLOG EcoPlates™ categorized by chemical structures |
 | |
Cylindrical containers (diameter 20 cm) were filled with contaminated and uncontaminated soil, respectively, to a soil column of approximately 28 cm height. Soil was let to settle one week prior to sowing; moisture content of both planted and unplanted soil was maintained at 60% Water Holding Capacity (WHC). The time of first watering was defined as the beginning of the experiment.
Plant Establishment and Maintenance
Three plants were established per container. All containers were kept in a greenhouse with mean monthly temperatures of 19 to 25°C and mean monthly relative humidity of 55 to 66% during the experimental period. A commercial compound fertiliser (Fertimax NPK 20-20-20, Brimport Seed C.A.) was applied with watering in a concentration of 10 g L-1 after 4 and 18 weeks. Total N, P and K concentrations were 400 mg kg-1 soil each.
Sampling
For sampling of planted soil, we distinguished between rhizosphere (R) and non-rhizosphere (NR) soil. Non-rooted soil and soil that fell off the root after gently shaking it were defined as non-rhizosphere soil, while soil firmly attached to the roots was defined as rhizosphere soil.
Carbon Source Utilization
BIOLOG Eco Plates™ were used to detect utilization patterns of carbon sources. The 96-well microplate contains 31 different carbon sources (Table 2) and one blank in three replicates, respectively. A tetrazolium dye indicates bacterial respiration and thus the use of a carbon source. 10 g soil (rhizosphere samples including 5 g roots) were shaken for 30 min (170 rpm, 25°C) in 90 mL of a sterile saline solution (0.85% NaCl). 100 μL of a 10-4 dilution were filled in each well and incubated at 27°C (Zak et al., 1994; Graystone et al., 1998). The intensity of colour change of the tetrazolium dye was measured with an optical density microplate reader (OD590 nm, SpectraMAX 340, Molecular Devices Corp.) every 24 h for 96 h. In order to eliminate variation which may arise from different cell densities in the samples, each optical density (OD) value was divided by the average well colour development (AWCD) which is the mean of the OD values for all 31 wells minus the blank well (Garland and Mills, 1991). An OD value higher than 0.2 was defined as positive carbon source utilization. ANOVA and principal component analysis (PCA) of OD values were performed with SPSS for Windows 11.5.0.
RESULTS
Number of Carbon Sources Used
There was a significant difference (p = 0.033) in total number of carbon sources used between contaminated and uncontaminated soil after 14 weeks (Table 3). Also when looking at functional groups (Fig. 1), differences existed mainly between contaminated and uncontaminated soil rather than between planted and unplanted soil or rhizosphere and non-rhizosphere soil. Contaminated soil used significantly more amines and carboxylic acids than uncontaminated soil (p = 0.002 and 0.004, respectively). The sampling time had no significant influence on functional groups but on total number of carbon sources.
Only the group of miscellaneous carbon sources (D,L-α-Glycerol phosphate, Glucose-1-phosphate, Pyruvic acid methyl ester) was significantly less used in planted (contaminated or uncontaminated) than in unplanted soil (p = 0.008). Additionally, time had a significant influence on the utilization of these carbon sources (p = 0.012).
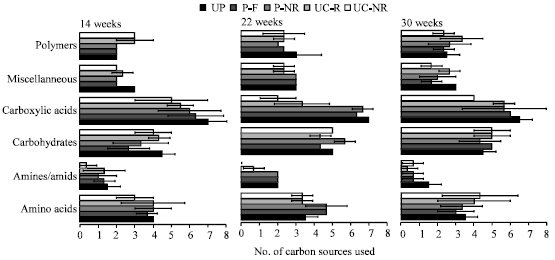 | |
| Fig. 1: | Total number of carbon sources used, grouped by chemical structures at three sampling dates in planted (P) and unplanted (UP) contaminated soil and in planted uncontaminated soil (UC). Rhizosphere soil (R), non-rhizosphere soil (NR). Values are means±standard deviation |
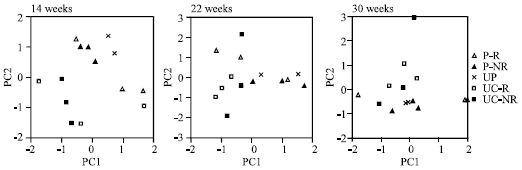 | |
| Fig. 2: | Principal Component (PC) analysis of substrate utilization patterns at three sampling dates of planted (P) and unplanted (UP) contaminated soils and of planted uncontaminated soil (UC). Rhizosphere soil (R), non-rhizophere soil (NR) |
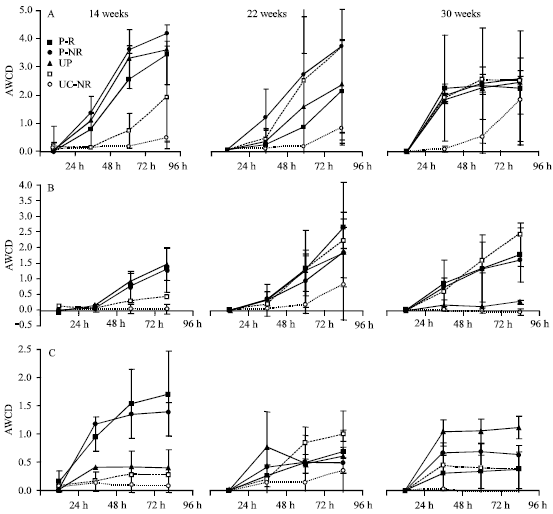 | |
| Fig. 3: | Utilization of D-galacturonic acid (A), D-glucosaminic acid (B) and D, L-α-glycerol phosphate (C) by soil samples of planted (P) and unplanted (UP) contaminated soil and of planted uncontaminated soil (UC), measured as Average Well Color Development (AWCD) after 24, 48, 72 and 96 h incubation, Rhizosphere soil (R), non-rhizosphere soil (NR). Values are means±standard deviation |
Principal Component Analysis
Principal Component (PC) analysis showed a tendency of division of microbial communities of contaminated and uncontaminated soil in two groups after 14 weeks (Fig. 2). The first two PCS accounted for 44 and 16% of the data set variance. In contaminated soil, three sub-groups could be assigned to rhizosphere, non-rhizosphere and unplanted soil, respectively (P-R, P-NR, UP). Microbial communities from P-R soil used itaconic acid, glucose-1-phosphate, I-erythritol, phenylethylamine and L-serine to a higher degree than P-NR and UP soil (PC2 axis). Between P-NR and UP soils, there was a slight separation on the PC1 axis. After 22 weeks, PC1 explained 33% and PC2 26% of total data variance. Division of the microbial communities of different treatments in separate groups was not as evident as after 14 weeks. Uncontaminated soil was divided on the PC1 axis from P-NR and UP soil and on the PC2 axis from P-R soil. Furthermore, P-R was separated from the remaining contaminated treatments on the PC1 axis, indicating a decreased use of several carbon sources (itaconic acid, γ-hydroxybutyric acid, putrescine, N-acetyl-D-glucosamine, α-cyclodextrin), while a slight separation on the PC2 axis showed an increased use of pyruvic acid methyl ester, D-mannitol, D-malic acid and L-arginine.
| Table 3: | Total number of carbon sources used at three sampling dates measured in planted (P) and unplanted (UP) contaminated soil and in planted uncontaminated soil (UC) after 96 h incubation in BIOLOG Eco Plates™. Rhizosphere soil (R), non-rhizosphere soil (NR). Values are means±standard deviation |
 | |
| * Values followed by the same letter do not significantly differ at P=0.05. None of the values measured after 22 and 30 weeks differed significantly | |
After 30 weeks, a division of microbial communities could not be detected. The P-R soil tended to separate on the PC1 axis which accounted for 29% of the variance of data. This indicated that soil microbial communities of P-R used several carboxylic acids (itaconic acid, D-galacturonic acid, γ-hydroxybutyric acid, D-malic acid, D-galactonic acid γ-lactone), putrescine, L-asparagine and N-acetyl-D-glucosamine to a higher degree than the remaining communities. On the PC2 axis, explaining 20% of data variance, UC soil had a tendency to separate from contaminated soil indicating a higher use of some amino acids (L-serine, L-arginine, glycyl-L-glutamic acid), pyruvic acid methyl ester and I-erythritol.
Utilization Patterns of Single Carbon Sources with Incubation Time
Data on utilization of single carbon sources at various incubation times in the BIOLOG plates (partly presented in Fig. 3) give additional information on differences in microbial communities. After 14 weeks, D-galacturonic acid (Fig. 3A) and β-methyl-D-glucoside (data not shown) were preferentially and more rapidly used by microbial communities of contaminated soil (p = 0.046). This preference, however, disappeared during the course of the experiment. At the end of the experiment, D-glucosaminic acid (Fig. 3B) was used to an higher extend by planted than by unplanted contaminated soil (p = 0,012). Also D,L-α-glycerol phosphate (Fig. 3C) showed a significantly increased use by microbial communities of planted vs. unplanted contaminated soil (p<0.05) after 14 weeks but not after 22 and 30 weeks.
DISCUSSION
Organic contaminants like petroleum hydrocarbons in soil mean toxic exposure for plants and soil microorganisms. The main mechanism of phytoremediation of organics is the stimulation of microbes living in the adjacent environment of the plant root, leading to an enhanced degradation of contaminants and thus a decreased toxic hazard for the plant. One possible mechanism by which plants can alter their rhizosphere community in response to environmental stresses is via exudates leading to detoxifying communities in the rhizosphere (Siciliano and Germida, 1998; Zak et al., 1994; Garland, 1996) found that rhizosphere communities from different plants have different substrate utilization patterns which should result from the qualitative (taxonomic diversity) and quantitative abundance of each taxon composition (Wünsche et al., 1995).
Number of Carbon Sources Used
With the BIOLOG Eco Plate™ assays we detected differences in microbial carbon source utilization patterns that were particularly evident between contaminated and uncontaminated soil. These results were expected since crude oil contaminated soil contains numerous additional carbon sources that promote the growth of a different and probably also more extensive set of microorganisms. A possible phytoremediation effect which we expected to be expressed in a higher number of carbon sources used in the rhizosphere could not be found.
Principal Component Analysis
The expected difference between contaminated rhizosphere soil and non-rhizosphere/unplanted soil became more evident by applying PCA, particularly after 14 weeks, documenting the rhizosphere effect on microbial community composition. This difference could hardly be detected by sole evaluation of numbers of carbon sources used. After 30 weeks, microbial community patterns (PCA) distinguished in the utilization of carboxylic acids and amino acids. These two groups were also mentioned by Banks et al. (1997) and Heinonsalo et al. (2000) to be utilized at a higher percentage by microbial communities in soil planted with clover and Scots pine, respectively, compared to unplanted soil. Heinonsalo et al. (2000) suggested that bacterial utilization of a greater range of carboxylic acids and amino acids reflects the increased availability of these carbon sources which are known to be important constituents of exudates.
Correlation of Microbial Community Patterns and Enhanced Oil Degradation
Merkl et al. (2005a) found enhanced crude oil dissipation in soil planted with B. brizantha compared to unplanted soil by 10.5% after 14 weeks. The residual oil of planted soil contained about 32% less saturates and 24% less aromatics than the control. After 22 weeks the difference was still visible but statistically not significant, while at the end of the experiment planted and unplanted soil had the same oil content. When comparing carbon source utilization patterns obtained by PCA with the enhancement of oil degradation in planted soil we can identify a similar development during the course of time. Microbial communities differed between rhizosphere and non-rhizosphere/unplanted soil at the beginning of the experiment and converged towards the end of the experiment. However, contrary to results found by Banks et al. (1997 and 2003), the microbial communities from planted soils with higher oil dissipation rates did not utilize a higher percentage of carbon substrates in the BIOLOG assay. Correspondingly, Heinonsalo et al. (2000) found that reduction in hydrocarbon content in soil over the incubation period was correlated with characteristic types but not overall numbers of utilized carbon sources. This points to a better evaluation of results by PCA and a limited explanatory power of the evaluation of number of carbon sources used.
Utilization Patterns of Single Carbon Sources with Incubation Time
D,L-α-glycerol phosphate (and partly β-methyl-D-glucoside) was the only carbon source that showed a significantly increased utilization in planted contaminated soil (P-R, P-NR) compared to unplanted or uncontaminated soil, accompanied by an enhanced oil degradation reported by Merkl et al. (2005a) after 14 weeks. That means that oil-degrading microbial communities are accompanied by an increased capacity to utilize this type of carbon source. It furthermore indicates a higher availability of structurally comparable carbon sources in the rhizosphere. Savannah soils of eastern Venezuela are particularly poor in phosphorus (P) and available P furthermore decreases during microbial degradation of hydrocarbons and plant growth. Additionally, P availability is usually lower in rhizosphere than in non-rhizosphere soil due to increased uptake by the roots (Marschner, 1995). However, under P deficiency in the soil P availability can increase in the rhizosphere due to plant exudates that temporarily increase P availability. For example, Zhang et al. (2001) stated that larch roots mobilize soil P and thus they found significantly higher P concentrations in rhizosphere than in non-rhizosphere soil. If P availability is higher in the rhizosphere it explains the increased use of structurally analogous compounds by rhizobial microbial communities. At the end of the experiment, utilization of D,L-α-glycerol phosphate is inverse (higher in unplanted than in planted soil) pointing to a P deficiency in the rhizosphere. At this time, no enhanced degradation in planted soil could be found.
D-glucosaminic acid also showed different utilization patterns during time. It was used to a higher extent by planted than by unplanted soil, but unlike D,L-α-glycerol phosphate, this occurred at the end of the experiment. In addition to the PCA results, these results show the plasticity of microbial community composition during time (here 16 weeks). Possible explanations are the dissipation of easily degradable petroleum hydrocarbons (low molecular weight constituents) and a decreased toxicity of contaminated soil.
CONCLUSIONS
The significance of the BIOLOG assay in this experiment was limited. Best results were obtained by PCA and by the evaluation of single carbon source utilization during time. Particularly useful concerning phytoremediation was the utilization of D,L-α-glycerol phosphate which correlated with enhanced oil degradation in soil. We suggest to further study single carbon sources that seem to be particularly indicative for a phytoremediation effect and that serve to identify degradative microbial communities. Reduction of the number of tested carbon sources of the present carbon source utilization assays to only a few decisive ones could furthermore facilitate and accelerate screening procedures of plant species suitable for phytoremediation.
It has to be considered that differences in microbial community structures detected by BIOLOG do not always correlate with enhanced degradation rates, as can be seen for D-glucosaminic acid and as was also shown by Fang et al. (2001b) for atrazine and phenanthrene degradation. Graystone et al. (1998) found that carbon source utilization patterns are strongly influenced by the plant species; Marschner et al. (2001), however, found that community structure in sandy soil and loamy sand was more affected by the root zone than by the plant species, while in clay, the root zone was less important. Also Girvan et al. (2003) stated that soil type is a key factor determining bacterial community composition. This would limit the applicability of the BIOLOG system for prediction and evaluation of phytoremediation.
It has also to be considered that fungi do not contribute to the community colour development profile of BIOLOG plates because they are incapable of utilizing the tetrazolium dye incorporated into them (Preston-Mafham et al., 2002). Moreover, fungal growth might interfere with the development of formazan-forming bacteria by competition for essential medium components (Wünsche et al., 1995). Previous studies showed that fungi play an important role in the decontamination of tropical soils impacted by oil spills (Oudot et al., 1993; Merkl et al., 2005c). Detection of differences in microbial community structures between planted and unplanted soil by the BIOLOG system could therefore be impeded or results be modified by fungi.
The data from the present work consolidated results obtained by oil degradation analyses and determination of microbial populations (Merkl et al., 2005a, c). Since the BIOLOG assay is easy to handle and fast and produces abundant data, we recommend it as a tool for phytoremediation studies. However, further work is needed to determine its importance for evaluating plant species for phytoremediation and predicting phytoremediation success. Focusing on single carbon sources as indicators of enhanced phytoremediation is an interesting aspect to be studied further on to meet the practical needs of plant selection for phytoremediation.
ACKNOWLEDGMENTS
We would like to thank Dr. Marianela Arias, Department of Ecology and Environment of PDVSA-Intevep (Centro de Investigación y Apoyo Tecnológico de Petróleos de Venezuela S.A.) and Dr. Howard Takiff, Centre of Microbiology and Cellular Biology of IVIC (Instituto Venezolano de Investigaciones Científicas) for technical and scientific support. The senior author was funded by a grant of the Gottlieb Daimler-und Carl Benz-Foundation, project number 02-07/01.
REFERENCES
- Fang, C., M. Radosevich and J.J. Fuhrmann, 2001. Characterization of rhizosphere microbial community structure in five similar grass species using FAME and BIOLOG analyses. Soil Biol. Biochem. 33: 679-682.
CrossRefDirect Link - Fang, C., M. Radosevich and J.J. Fuhrmann, 2001. Atrazine and phenanthrene degradation in grass rhizosphere soil. Soil Biol. Biochem., 33: 671-678.
CrossRefDirect Link - Garland, J.L. and A.L. Mills, 1991. Classification and characterization of heterotrophic microbial communities on the basis of pattern of community-level sole-carbon-source utilization. Applied Environ. Microbiol., 57: 2351-2359.
PubMed - Garland, J.L., 1996. Patterns of potential C source utilization by rhizosphere communities. Soil Biol. Biochem., 28: 223-230.
Direct Link - Girvan, M.S., J. Bullimore, J.N. Pretty, A.M. Osborne and A.S. Ball, 2003. Soil type is the primary determinant of the composition of the total and active bacterial communities in arable soils. Applied Environ. Microbiol., 69: 1800-1809.
CrossRefDirect Link - Grayston, S.J., S. Wang, C.D. Campbell and A.C. Edwards, 1998. Selective influence of plant species on microbial diversity in the rhizosphere. Soil Biol. Biochem., 30: 369-378.
CrossRefDirect Link - Haack, S.K., H. Garchow, M.J. Klug and L.J. Forney, 1995. Analysis of factors affecting the accuracy, reproducibility and interpretation of microbial community carbon source utilization patterns. Applied Environ. Microbiol., 61: 1458-1468.
Direct Link - Heinonsalo, J., K. Jorgensen, K. Haahtela and R. Sen, 2000. Effect of Pinus sylvestris root growth and mycorrhizosphere development on bacterial carbon source utilization and hydrocarbon oxidation in forest and petroleum contaminated soils. Can. J. Microbiol., 46: 451-464.
Direct Link - Hutchinson, S.L., M.K. Banks and A.P. Schwab, 2001. Phytoremediation of aged petroleum sludge: Effect of inorganic fertilizer. J. Environ. Quality, 30: 395-403.
Direct Link - Joner, E.J., S.C. Corgie, N. Amellal and C. Leyval, 2002. Nutritional constraints to degradation of polycyclic aromatic hydrocarbons in a simulated rhizosphere. Soil Biol. Biochem., 34: 859-864.
CrossRefDirect Link - Joner, E.J. and C. Leyval, 2003. Phytoremediation of organic pollutants using mycorrhizal plants: A new aspect of rhizosphere interactions. Agronomie, 23: 495-502.
CrossRefDirect Link - Kudjo, D.E., T. Chekol and L.R. Vough, 2000. Feasibility of using forage grasses and legumes for phytoremediation of organic pollutants. J. Environ. Sci. Health, 35: 1645-1661.
Direct Link - Leahy, J.G. and R.R. Colwell, 1990. Microbial degradation of hydrocarbons in the environment. Microbiol. Mol. Biol. Rev., 54: 305-315.
Direct Link - Lin, Q. and I.A. Mendelssohn, 1998. The combined effects of phytoremediation and biostimulation in enhancing habitat restoration and oil degradation of petroleum contaminated wetlands. Ecol. Eng., 10: 263-274.
CrossRefDirect Link - Marschner, P., C.H. Yang, R. Lieberei and D.E. Crowley, 2001. Soil and plant specific effects on bacterial community composition in the rhizosphere. Soil Biol. Biochem., 33: 1437-1445.
CrossRefDirect Link - Merkl, N., R. Schultze-Kraft and M. Arias, 2005. Influence of fertiliser levels on phytoremediation of crude oil with the tropical pasture grass Brachiaria brizantha (Hochst. ex A. Rich.) Stapf. Intl. J. Phytoremed., 7: 217-230.
Direct Link - Kirk, J.L., J.N. Klironomos, H. Lee and J.T. Trevors, 2002. Phytotoxicity assay to assess plant species for phytoremediation of petroleum-contaminated soil. Bioremed. J., 6: 57-63.
CrossRefDirect Link - Merkl, N., R. Schultze-Kraft and M. Arias, 2005. Effect of the tropical grass Brachiaria brizantha (Hochst. ex A. Rich.) Stapf on microbial population and activity in petroleum-contaminated soil. Microbiol. Res., 161: 80-91.
Direct Link - Oudot, J., J. Dupont, S. Haloui and M.F. Roquebert, 1993. Biodegradation potential of hydrocarbon-assimilating tropical fungi. Soil Biol. Biochem., 25: 1167-1173.
Direct Link - Pradhan, S.P., J.R. Conrad, J.R. Paterek and V.J. Srivastava, 1998. Potential of phytoremediation for treatment of PAHs in soil at MGP sites. J. Soil Contamin., 7: 467-480.
Direct Link - Preston-Mafham, J., L. Boddy and P.F. Randerson, 2002. Analysis of microbial community functional diversity using sole-carbon-source utilisation profiles-a critique. FEMS Microbiol. Ecol., 42: 1-14.
Direct Link - Wiltse, C.C., W.L. Rooney, Z. Chen, A.P. Schwab and M.K. Banks, 1998. Greenhouse evaluation of agronomic and crude oil-phytoremediation potential among alfalfa genotypes. J. Environ. Quality, 27: 169-173.
Direct Link - Wunsche, L., L. Bruggemann and W. Babel, 1995. Determination of substrate utilization patterns of soil microbial communities: An approach to assess population changes after hydrocarbon pollution. FEMS Microbiol. Ecol., 17: 295-306.
Direct Link - Yoshitomi, K.J. and J.R. Shann, 2001. Corn (Zea mays L.) root exudates and their impact on 14C-pyrene mineralization. Soil Biol. Biochem., 33: 1769-1776.
Direct Link - Zak, J.C., M.R. Willig, D.L. Moorhead and H.G. Wildman, 1994. Functional diversity of microbial communities: A quantitative approach. Soil Biol. Biochem., 26: 1101-1108.
Direct Link - Zhang, Y., S. Bai, Z. Wan and Q. Wang, 2001. Soil P availability in larch rhizosphere. Chinese J. Applied Ecol., 12: 31-34.
Direct Link