
Joice J. Bana
School of Life Sciences and Technology, Institut Teknologi Bandung (ITB), Jalan Ganesa No.10, Bandung 40132, Indonesia
LiveDNA: 62.31413
Anggraini Barlian
School of Life Sciences and Technology, Institut Teknologi Bandung (ITB), Jalan Ganesa No.10, Bandung 40132, Indonesia
Ahmad Ridwan
School of Life Sciences and Technology, Institut Teknologi Bandung (ITB), Jalan Ganesa No.10, Bandung 40132, Indonesia
International Journal of Poultry Science
Year: 2021 | Volume: 20 | Issue: 6 | Page No.: 249-255







ABSTRACT
Background and Objective: Prolactin is a steroid hormone that induces broodiness in chickens. In addition to the prolactin hormone, the incubating nature of chickens is also influenced by genes. The purpose of this study was to analyze changes in the profile of the prolactin hormone; the pattern and level of expression of the genes Prolactin (Prl), Vasoactive Intestine Peptide (VIP), Pituary specific transcription factor-1 (Pit-1) and prolactin regulatory element binding protein (PREB) due to the administration of each dose of antiprolactin (2-Bromo-α-Ergocryptine Methanesulfonate salt/Bromocriptine); as well as the relationship between prolactin hormone secretion and these genes. Materials and Methods: Blood and brain samples from 25 hens (Kampung chickens) fed various doses of anti-prolactin were used in this study. Serum from each blood sample was used to calculate the concentration of the hormone prolactin by ELISA method. Meanwhile, the brains of each free-range chicken were used to measure the pattern and level of expression of the Prl, VIP, Pit-1 and PREB genes using the Quantitative Real Time-Polymerase Chain Reaction (RT-qPCR) method. Results: Increase in the doses of anti-prolactin decreased the concentration of the hormone prolactin as well as the pattern and level of expression of the Prl, VIP, Pit-1 and PREB genes. Conclusion: Anti-prolactin can inhibit the secretion of the hormone prolactin and the expression of Prl, Pit-1, VIP and PREB genes.
PDF Abstract XML References Citation
Copyright: © 2021. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Joice J. Bana, Anggraini Barlian and Ahmad Ridwan, 2021. Prolactin Hormone Profile, Patterns and Expression Level of Prolactin, Pit-1, VIP and PREB Gene in Kampung Chicken (Gallus gallus domesticus) Induced by Anti-Prolactin. International Journal of Poultry Science, 20: 249-255.
DOI: 10.3923/ijps.2021.249.255
URL: https://scialert.net/abstract/?doi=ijps.2021.249.255
DOI: 10.3923/ijps.2021.249.255
URL: https://scialert.net/abstract/?doi=ijps.2021.249.255
INTRODUCTION
Prolactin hormone is a steroid hormone produced by the anterior pituitary and is responsible for various physiological processes in vertebrates, including reproduction, osmoregulation, growth and development, metabolism, immune regulation, energy balance and behavior1. In mammals the hormone prolactin is responsible for lactation, whereas in fowl this hormone is responsible for inducing brooding behavior 2.
The prolactin hormone encoded by the prolactin gene is not the only determinant of broodiness. The genes and/or candidate genes that are known to be associated with broodiness have been summarized by Romanov3 --namely, the estrogen receptor (ESR) gene; the progesterone receptor gene (PGR); the prolactin gene (Prl); the prolactin receptor gene (PRLR); the vasoactive intestinal peptide (VIP) gene; the vasoactive intestinal peptide receptor gene (VIPR); the luteinizing hormone/choriogonadotropin receptor gene (LHCGR); the luteinizing hormone-releasing hormone 1 (LHRH1); the dopamine D1D receptor (D1LR) gene; the growth hormone gene (GH1); the gene growth hormone receptor (GHR) and the gonadotrophin releasing hormone 1 (GNRH). Other genes that have also been found and are thought to be associated with broodiness are the prolactin promoter gene, the Pit-1 gene, the GARLN1 gene, the prolactin regulatory element binding protein gene and the dopamine D2 receptor (DRD2) gene4-8.
The genes associated with broodiness are thought to be related to the regulation of the secretion of the hormone prolactin. For this reason, this study was conducted to examine the relationship between these genes and prolactin hormone secretion and the mechanism of regulation of prolactin hormone secretion related to brooding properties.
In this study, four genes (Prl, Pit-1, PREB and VIP) associated with broodiness were used. Various doses of anti-prolactin were administered to observe the expression pattern of the four genes associated with the secretion of the hormone prolactin.
MATERIALS AND METHODS
Ethical clearance: All experimental procedures were approved by the Animal Ethics Committee of the Faculty of Veterinary Medicine, Nusa Cendana University (KEH/FKH/NPEH/2019/001). All efforts were taken to minimise pain and discomfort to the animal while conducting experiment.
Study sites: This research was conducted for six months, from July to December 2020, in the Laboratory of Molecular Genetics, School of Life Sciences and Technology, ITB, Bandung, West Java (Indonesia).
Prolactin hormone analysis: A total of 25 hens of reproductive age (7-8 months) were used in this study. Hens were divided into five treatment groups: control group (P0) water was provided without 2-Bromo-α-Ergocryptine Methanesulfonate salt; treatment 1 (P1) 2-Bromo-α-Ergocryptine Methanesulfonate salt was provided at a dose of 0.12 mg kg–1 body weight; treatment 2 (P2) 2-Bromo-α-Ergocryptine Methanesulfonate salt was provided at a dose of 0.35 mg kg–1 body weight; treatment 3 (P3) 2-Bromo-α-Ergocryptine Methanesulfonate salt was provided at a dose of 0.70 mg kg–1 body weight and treatment 4 (P4) 2-Bromo-α-Ergocryptine Methanesulfonate salt was provided at a dose of 1.40 mg kg–1 bw orally9 using table of conversion of dose calculations10. The compound 2-Bromo-α-Ergocryptine Methanesulfonate salt was given when the chicken showed signs of brooding.
Prolactin hormone analysis was carried out using the ELISA method, according to the manufacturer’s protocol (Fisher Invitrogen 2017). Blood samples from each test animal were taken from the branchial vein in the wing using a vacutainer on days 1, 2, 3 and 4 of the brooding phase. The blood was left for a while until clotting occurred and serum was formed. The formed serum was transferred to an Eppendorf tube stored in a freezer and then taken to the laboratory for the ELISA test.
Statistical analysis: The one-way analysis of variance (ANOVA) was applied to determine effect of treatment on the prolactin hormone profile. If there was a significant effect, the analysis continued with Duncan's test at 5% level of significance. All statistical analyses were performed using SPSS 25.0 program.
Measurement of VIP, Prl, Pit-1, PREB and GAPDH gene expression patterns and levels: The test animals were sacrificed by cervical dislocation and then the brain was isolated to measure the pattern and level of expression of the VIP, Prl, Pit-1, PREB and GAPDH genes as housekeeping genes.
Total RNA isolation: Brain organs of test animals from each treatment and replication were isolated and immediately put in vials containing RNA Later solution. Then they were stored at 4°C and brought to the laboratory and stored at -20°C. Brain samples were isolated for total RNA using the Promega Kit according to the manufacturer’s specification. The obtained RNA solution was measured for its RNA concentration using a Nano Drop, then stored at -70 for further analysis.
Expression testing of Prl, Pit-1, VIP and PREB genes: Brood gene expression was tested by using web-based (www.Primer3plus.ut) specific primer. The following primers were used in this study:
Reverse transcriptase-polymerase chain reaction (RT-PCR): The total RNA solution obtained with known concentration was converted into cDNA (complementary DNA) through the Reverse Transcription (RT) PCR process using the RevertAid First Strand cDNA Synteis Kit (Thermo Scientific) with 1 μL Oligo dT primer and 1 μL Reverse Transcriptase enzyme. The results of cDNA synthesis were stored at -70°C.
Quantitative real time polymerase chain reaction (RT-qPCR): Genes amplification of Pit-1, VIP, PRL, PREB and GAPDH used applied biosystems QuantStudio1 Thermal Cycler (Therma Fisher Scientific) and Kit GoTag®2-Step RT-qPCR System according to protocol (Promega, 2010) with cDNA sample concentration of 10 ng μL–1.
RESULTS AND DISCUSSION
Prolactin hormone concentration: Table 1 and Fig. 1 shows the average concentration of prolactin hormone after anti-prolactin treatment. The concentration of prolactin hormone decreased with increase in the dose of anti-prolactin. The control treatment (without anti-prolactin) showed the highest concentration of prolactin hormone and was significantly different from other treatments (p<0.05). The high concentration of prolactin in the control group indicates that the hormone prolactin plays a very important role in inducing brooding properties in chickens. High concentrations of the hormone prolactin are associated with incubation of eggs or parental behavior of some bird species11. Sharp et al.12 stated that increase in the secretion of the hormone prolactin induces the brooding behavior in chicken and thus plays an important role in production performance.
The greatest decrease (1.4 mg kg–1 body weight) in prolactin hormone concentration was observed in the P4 treatment, (Fig. 1), however, it was not significantly different (p<0.05) from other treatments (P1, P2 and P3). This decrease in the concentration of the prolactin hormone in P4 resulted in the shortest average egg-laying time (10.4-10.6 days) (unpublished data). This condition indicates that the administration of anti-prolactin (bromocriptine) can inhibit the secretion of the prolactin hormone. Berry13 stated that brooding activity in poultry is controlled by the prolactin hormone, one of the gonadotropin hormones produced by the adenohypophysis. This opinion is reinforced by Molik and Błasiak14 who stated that bromocriptine can inhibit the production and secretion of prolactin. Meanwhile, Freeman et al.15 stated that bromocriptine, which is a dopamine agonist, can act as a dose-dependent prolactin stimulator and inhibitor. At high doses, it can inhibit the secretion of the prolactin hormone. It is suspected that in this study bromocriptine at various doses also inhibited the secretion of the hormone prolactin and the greatest inhibition occurred at a dose of 1.4 mg kg–1 body weight. There was no significant difference between treatments (P1, P2, P3 and P4), due to a minor decrease in the prolactin hormone concentration making it difficult to detect the difference.
Expression pattern of Prl, VIP, Pit-1 and PREB genes: Figure 2 shows the expression patterns of Prl, VIP, Pit-1 and PREB genes treated with anti-prolactin at various doses. Histograms of gene expression patterns of the four genes showed a decreasing effect in all treatments compared to controls. The decrease in the expression pattern of these genes indicates that these genes play a role in inducing broodiness behaviour in chickens. The incubation behaviour is induced by the prolactin hormone16 and incubating genes3. Prl, VIP, Pit-1 and PREB genes play a role in inducing brooding properties3,4,8. The anti-prolactin hormone decreased the concentration of Prl (Fig. 1) and the decrease was in line with the decrease in the expression of Prl, VIP, Pit-1 and PREB genes. This finding indicates a close relationship between the inhibition of Prl hormone secretion due to anti-prolactin administration and the expression level of brood traits genes. Gene, which codes for the hormone prolactin, is activated by a number of factors including pituitary specific transcription factor (Pit-1) and vasoactive intestinal peptide (VIP)17,18.
The administration of bromocriptine (anti-prolactin) decreased the secretion of the hormone prolactin (Fig. 1) due to the inhibition of VIP. Talbot et al.19 stated that the release of prolactin from the anterior pituitary gland and the expression of the Prl gene are regulated first by the hypothalamic factor, vasoactive intestinal polypeptide (VIP). VIP injection increased Prl plasma concentrations and Prl mRNA levels in the pituitary gland of laying hens while passive anti-VIP immunization decreased Prl plasma concentrations and Prl mRNA levels in the anterior pituitary gland of brooding hens. Furthermore, Kansaku et al.,20 stated that the cAMP-PKA pathway is involved in the release and expression of the Prl gene in poultry because VIP induces an increase in intracellular cAMP levels.
The greatest decrease was observed in the Prl gene in treatment P4, followed by PREB, VIP and Pit-1. This finding shows that there is a close relationship between inhibition of Prl hormone secretion due to anti-prolactin administration and the level of expression of incubating genes.
Relationship of bromocriptine, prolactin hormone secretion with gene expression level of broodiness: Bromocriptine is a dopamine agonist commonly used for the treatment of hyperprolactinemia. Dopamine is a neurotransmitter that is abundant in the hypothalamus and is a prolactin inhibitor (PIF) factor15. It was further stated that the presence of high concentrations of dopamine in the median eminence and hypoxic blood vessels reinforce the assumption that dopamine is the greatest PIF of the pituitary physiologically. Several studies have shown that dopamine inhibits the release of prolactin under both in vitro and in vivo conditions. A decrease in the expression level of the Prl gene was followed by a decrease in the expression level of the VIP gene. The VIP gene is a releasing factor of the hormone prolactin in poultry21 and high concentrations of VIP in the hypothalamus correlate with plasma Prl22. Furthermore, Ottinger and Bakst23 stated that VIP is important in regulating the secretion of prolactin, which is a hormone associated with brooding behavior.
Xu et al.24 found that hens treated with dopamine terminated brooding behavior due to inhibition of prolactin secretion. Al Kahtane et al.25 reported that in avian species, dopamine inhibits VIP, which stimulates the amount of Prl mRNA at both transcriptional and post-transcriptional stages.
This study also showed a decrease in the expression level of the Pit-1 and PREB genes due to the administration of bromocriptine and this decrease in expression level increased with increasing doses of anti-prolactin. Pit-1 and PREB are transcription factors of the Prl gene where PREB specifically binds to Pit-1 at the Prl promoter site to regulate Prl gene expression in mammals. This result indicates that Pit-1 is associated with increased level of PRL mRNA and then interacting with PREB to support the further increase in PRL mRNA expression in late stages of embryogenesis7. Fliss et al.26 suggested that these two transcription factors work together to increase the expression level of the Prl gene. The results of Hiyama et al.7 showed an increase in Prl, Pit-1 and PREB mRNA levels in the anterior pituitary gland of White Leghorn chickens. From these results, it is suspected that Pit-1 plays a role in the basal expression of PRL mRNA but PREB and Pit-1 may have an additional effect on changes in PRL mRNA levels in the laying and incubating phases as seen in 18-day-old embryos until hatching. A follow-up study conducted by Hiyama et al.8 showed that increase in the levels of Prl mRNA is in line with changes in PREB levels and the highest levels of Prl and PREB were reached on the 25th day of observation. Hiyama et al.7,8 stated that an increase in the transcription factor Pit-1 and PREB will result in an increase in PRL levels8.
Molik and Błasiak14 stated that the inhibitors of the secretion of the hormone prolactin are dopamine (DA), gamma amino bituric acid (GABA), progesterone and bromocriptine (exogenous component) which are used in D2 receptor agonist hyperprolactinemia. The biological activity of this component is to stimulate D2 receptors on the pituitary, which inhibits the secretion of the hormone prolactin through the tuberoinfundibular dopaminergic (TIDA) pathway27. Dopaminergic tuberoinfundibular is a form of the nervous system that secretes dopamine to the anterior pituitary via the long portal vessel system.
The inhibition of prolactin hormone secretion by bromocriptine (dopamine agonist) is thought to follow the dopamine pathway where bromocriptine binds to D2R receptors on the surface of lactotrope cells and inhibits the secretory activity of these cells (Fig. 3). Through the D2 receptor, bromocriptine inhibits prolactin secretion and causes inhibition of adenylate cyclase (AC) through Gi proteins by G0 proteins mediated calcium channels blockage. The hyperpolarization of the cell membrane and the closure of calcium channels induced by bromocriptine stimulates the potassium channel via the G13 protein to decrease the calcium level in the cell and inhibit the secretion of prolactin. Lincoln et al.28 stated that dopamine binds to DA-D2 in GPCR inhibit Adenylate cyclase. This DA-G complex causes cell membrane hyperpolarization and closes Ca2+ ions and opens K+ ion channels, causing a decrease in Ca2+ levels and inhibiting Prl hormone secretion. Allegedly, in this study, this reaction occurred after the binding of bromocriptine administered to the D2R receptor. Elsholtz et al.4 stated that the addition of dopamine to GH4ZR7 cell cultures reduced cAMP levels. The results of this study strengthen the notion that the use of bromocriptine (dopamine agonist) inhibits the action of adenylate cyclase which in turn inhibits the formation of cAMP.
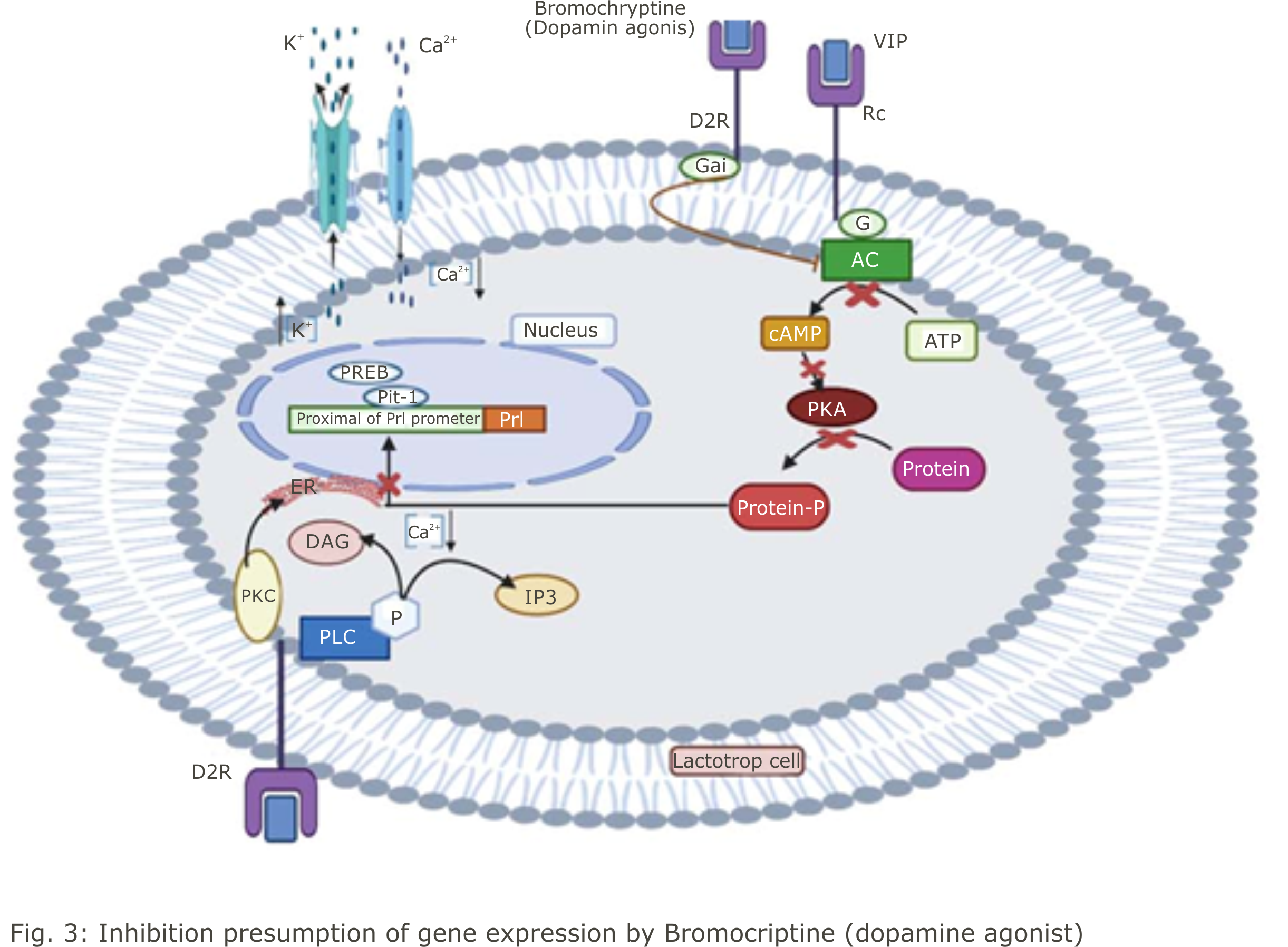
Meanwhile, labeled VIP exhibits binding to lactotrope cells and stimulates prolactin secretion via adenyl cyclase activation29. Bromocriptine inhibits adenyl cyclase activity which inhibits the work of VIP which in turn inhibits all processes to activate the expression of the Pit-1, PREB and prolactin genes, thereby reducing the production of the hormone prolactin. cAMP is a key in intracellular signal transduction in hormone, neurotransmitter and chemokinesis synthesis30. Further Elsholtz, et al.4 stated that cAMP stimulates transcription of the prolactin gene and reduces the inhibitory effect of ergocryptine on that gene. Ergocryptine is a another dopamine agonist.
CONCLUSION
Administration of anti-prolactin (2-Bromo-α-Ergocryptine Methanesulfonate salt) decreased the concentration of the hormone prolactin and the pattern and level of expression of the genes Prl, VIP, Pit-1 and PREB.
ACKNOWLEDGMENTS
This study was financially supported by the Indonesian Endowmen Fund for Education (LPDP), the Ministry of Indonesia. Financial support was awarded to the first author with grant number {PJR-5716/LPDP.3/2016}.
SIGNIFICANCE STATEMENT
This study revealed the effect of anti-prolactin on the secretion of the hormone prolactin and the pattern and level of expression of brooding genes. The outcome of this study can be used to improve the productivity of kampung chickens by manipulating their reproduction both hormonally and molecularly.
REFERENCES
- Marano, R.J. and N. Ben-Jonathan, 2014. Minireview: Extrapituitary prolactin: An update on the distribution, regulation, and functions. Mol. Endocrinol., 28: 622-633.
CrossRefDirect Link - Riddle, O., R.W. Bates and E.L. Lahr, 1935. Prolactin induces broodiness in fowl. Am. J. Physiol., 111: 352-360.
CrossRefDirect Link - Romanov, M.N., 2001. Genetics of broodiness in poultry - A review. Asian-Australas. J. Anim. Sci., 14: 1647-1654.
CrossRefDirect Link - Elsholtz, H.P., A.M. Lew, P.R. Albert and V.C. Sundmark, 1991. Inhibitory control of prolactin and pit-1 gene promoters by dopamine. Dual signaling pathways required for D2 receptor-regulated expression of the prolactin gene. J. Bio. Chem., 266: 22919-22925.
CrossRefDirect Link - Sartika, T., 2005. Broodiness trait of chicken through molecular investigation. Wartazoa: Indonesian Bull. Anim. Vet. Sci., 15: 206-212.
CrossRefDirect Link - Shen, X., H. Zeng, L. Xie, J. He and J. Li et al., 2012. The GTPase activating Rap/RanGPA domain-like 1 gene is associated with chicken reproductive traits. PLoS ONE, Vol. 7.
CrossRefDirect Link - Hiyama, G., N. Kansaku, T. Tanaka, S. Wakui and D. Zadworny, 2015. Characterization of chicken prolactin regulatory element binding protein and its expression in the anterior pituitary gland during embryogenesis and different reproductive stages. J. Poult. Sci., 52: 42-51.
CrossRefDirect Link - Hiyama, G., N. Kansaku, S. Wakui, R. McQuaid and D. Zadworny, 2015. Characterization and expression of Turkey prolactin regulatory element binding in the anterior pituitary gland and pancreas during embryogenesis. J. Poult. Sci., 53: 67-75.
CrossRefDirect Link - Bacharach, A.L. and D.R. Laurence, 1964. Preface. In: Evaluation of Drug Activities: Pharmacometrics Laurence, D.R. and A.L. Bacharach, (Eds.). Academic Press, New York, Pages: 4.
CrossRefDirect Link - Buntin, J.D., R.M. Hnasko and P.H. Zuzick, 1999. Role of the ventromedial hypothalamus in prolactin-induced hyperphagia in ring doves. Physiol. Behav., 66: 255-261.
CrossRefDirect Link - Sharp, P.J., A. Dawson and R.W. Lea, 1998. Control of luteinizing hormone and prolactin secretion in birds. Comp. Biochem. Physiol. Part C: Pharmacol., Toxicol. Endocrinol., 119: 275-282.
CrossRefDirect Link - Molik, E. and M. Błasiak, 2015. The role of melatonin and bromocriptine in the regulation of prolactin secretion in animals - A review. Ann. Anim. Sci., 15: 849-860.
CrossRefDirect Link - Freeman, M.E., B. Kanyicska, A. Lerant and G. Nagy, 2000. Prolactin: Structure, function, and regulation of secretion. Physiol. Rev., 80: 1523-1631.
CrossRefDirect Link - Kansaku, N., G. Hiyama, T. Sasanami and D. Zadworny, 2008. Prolactin and growth hormone in birds: Protein structure, gene structure and genetic variation. J. Poult. Sci., 45: 1-6.
CrossRefDirect Link - Youngren, O.M., Y. Chaiseha and M.E.E. Halawani, 1998. Regulation of prolactin secretion by dopamine and vasoactive intestinal peptide at the level of the pituitary in the Turkey. Neuroendocrinology, 68: 319-325.
CrossRefDirect Link - Usman, M., A. Basheer, M. Akram, M.E. Babar and I. Zahoor, 2014. Prolactin: Candidate gene for egg production or broodiness traits in chicken. Sci. Int., 26: 1191-1195.
Direct Link - Talbot, R.T., M.C. Hanks, R.J. Sterling, H.M. Sang and P.J. Sharp, 1991. Pituitary prolactin messenger ribonucleic acid levels in incubating and laying hens: effects of manipulating plasma levels of vasoactive intestinal polypeptide. Endocrinology, 129: 496-502.
CrossRefDirect Link - Kansaku, N., K. Shimada, N. Saito and H. Hidaka, 1998. Effects of protein kinase A inhibitor (H-89) on VIP-and GRF-induced release and mRNA expression of prolactin and growth hormone in the chicken pituitary gland. Comp. Biochem. Physiol. C: Pharmacol. Toxicol. Endocrinol., 119: 89-95.
CrossRef - Kang, S.W., O.M. Youngren and M.E.E. Halawani, 2002. Influence of VIP on prolactinemia in Turkey anterior pituitary cells: Role of cAMP second messenger in VIP-induced prolactin gene expression. Regul. Peptides, 109: 39-44.
CrossRefDirect Link - Guemene, D. N. Kansaku and D. Zadworny, 2001. Incubation behaviour expression in Turkey hens and its control in commercial flocks. Prod. Anim., 14: 147-160.
Direct Link - Ottinger, M.A. and M.R. Bakst, 1995. Endocrinology of the avian reproductive system. J. Avian Med. Surg., 9: 242-250.
Direct Link - Xu, H., X. Shen, M. Zhou, M. Fang, H. Zeng, Q. Nie and X. Zhang, 2010. The genetic effects of the dopamine D1 receptor gene on chicken egg production and broodiness traits. BMC Genet., Vol. 11.
CrossRefDirect Link - Kahtane, A.A., Y. Chaiseha and M.E. Halawani, 2003. Dopaminergic regulation of avian prolactin gene transcription. J. Mol. Endocrinol., 31: 185-196.
CrossRefDirect Link - Fliss, M.S., P.M. Hinkle and C. Bancroft, 1999. Expression cloning and characterization of PREB (prolactin regulatory element binding), a novel WD motif DNA-binding protein with a capacity to regulate prolactin promoter activity. Mol. Endocrinol., 13: 644-657.
CrossRefDirect Link - Fitzgerald, P. and T.G. Dinan, 2008. Prolactin and dopamine: What is the connection? A review. J. Psychopharmacol., 22: 12-19.
CrossRefDirect Link - Lincoln, G.A., H. Andersson and D. Hazlerigg, 2003. Clock genes and the long-term regulation of prolactin secretion: Evidence for a photoperiod/circannual timer in the pars tuberalis. J. Neuroendocrinol., 15: 390-397.
CrossRef - Bjøro, T., O. Sand, B.C. Østberg, J.O. Gordeladze, P. Torjesen, K.M. Gautvik and E. Haug, 1990. The mechanisms by which vasoactive intestinal peptide (VIP) and thyrotropin releasing hormone (TRH) stimulate prolactin release from pituitary cells. Biosci. Rep., 10: 189-199.
CrossRefDirect Link - Hurley, J.H., 1999. Structure, mechanism, and regulation of mammalian adenylyl cyclase. J. Bio. Chem., 274: 7599-7602.
CrossRefDirect Link