
T. Cao
Noveltech, LLC., 16600 43rd Ave N., Minneapolis, MN 55446, United States
J.T. Weil
Center of Excellence for Poultry Science, University of Arkansas, Fayetteville, AR 72701, United States
P. Maharjan
Center of Excellence for Poultry Science, University of Arkansas, Fayetteville, AR 72701, United States
J. Lu
Syngenta, 410 S. Swing Rd., Greensboro, NC 27409, United States
C.N. Coon
Center of Excellence for Poultry Science, University of Arkansas, Fayetteville, AR 72701, United States
LiveDNA: 1.35097
International Journal of Poultry Science
Year: 2020 | Volume: 19 | Issue: 5 | Page No.: 232-243







ABSTRACT
Background and Objective: Two experiments were conducted to determine the total sulfur amino acid requirements in laying hens. The objective of Experiment 1 was to determine the digestible methionine and cystine requirements for laying hens. An additional experiment (Experiment 2) was conducted to determine the cystine requirement for laying hens and determine the utilization efficiencies of supplemental methionine and cystine to meet the cystine requirement. Materials and Methods: In Experiment 1, one hundred and seventy-six laying hens were randomly assigned into 11 dietary treatments for a six-week period. One group of hens received a corn-soybean meal control diet containing 2,899 kcal ME kg–1 and 19.5% CP, while the remaining ten groups of hens received 10 test diets containing 2,850 kcal ME kg–1 and 15% CP. Five diets were deficient in cystine (0.148% digestible cystine), containing digestible methionine levels of 0.143, 0.240, 0.337, 0.434 and 0.531% and another five diets were excessive in cystine (0.450% digestible cystine), containing digestible methionine levels of 0.143, 0.231, 0.317, 0.407 and 0.495%. An additional experiment (Experiment 2) was conducted by assigning one hundred sixty laying hens to one of two series of diets, which were formulated to contain 0.319% digestible methionine and 0.148% digestible cystine, same as that in Experiment 1, with exception of the methionine level. Four levels of equimolar amounts of methionine or cysteine (½ cystine) were added to the basal diet. The added levels were 0.05, 0.10, 0.15 and 0.20% for methionine and 0.04, 0.08, 0.12 and 0.16% for cystine since the molecular weight of cysteine (½ cystine) is 80% of that of methionine. Data generated from each experiment was analyzed using the general linear models (GLM) and analysis of variance procedures with the help of statistical analysis software (SAS). A second-order polynomial regression analysis was conducted in order to determine the methionine requirements for laying hens. Results: The results showed that the requirement of digestible methionine and digestible cysteine for laying hens were 354 and 184 mg hen–1 day–1 for egg mass (EM), 349 and 193 mg hen–1 day–1 for feed conversion, 437 and 325 mg hen–1 day–1 for body weight change (BWC) and 367 and 189 mg hen–1 day–1 for EM+BWC, respectively. Deficient or excessive dietary methionine produced an increase of methionine degradation due to the increased body weight loss or the excessive dietary methionine, correspondingly. Optimum dietary methionine levels resulted in increased liver SAM/SAH concentration ratios (s-adenosylmethionine/s-adenosylhomocysteine) and decreased homocysteine (Hcy) levels. Conclusion: The results demonstrated that the utilization efficiencies of methionine and cysteine (½ cystine) were 100% on an equimolar basis for egg mass and 90% on an equimolar basis to prevent loss of body weight. When methionine was used to meet the cystine requirement, an utilization efficiency of 80% was adequate on a weight and concentration basis for egg mass and 72% for body weight maintenance. The practice of feeding ingredients with a substantial digestible cystine level for supporting body weight may be beneficial for laying hens.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
T. Cao, J.T. Weil, P. Maharjan, J. Lu and C.N. Coon, 2020. The Digestible Methionine and Cystine Requirements for Commercial Layers. International Journal of Poultry Science, 19: 232-243.
DOI: 10.3923/ijps.2020.232.243
URL: https://scialert.net/abstract/?doi=ijps.2020.232.243
DOI: 10.3923/ijps.2020.232.243
URL: https://scialert.net/abstract/?doi=ijps.2020.232.243
INTRODUCTION
The sulfur amino acid requirement of laying hens has been traditionally expressed as a methionine requirement and total sulfur amino acid (TSAA) requirement1. The concept of the requirement implies that the demand for sulfur amino acids of laying hens can be satisfied with either methionine alone or through the combination of methionine and cystine. This is supported by the fact that methionine can be endogenously converted to cysteine whereas the opposite is not possible because of a lack of enzymes2,3. However, due to the limited or lack of information that exists about the efficiency of methionine conversion to cysteine, the quantitative relationship of cystine for sparing methionine and the utilization efficiency of supplemental methionine and cystine, there is confusion about the reported requirement.
In recent years, the indicator amino acid oxidation method has been used to determine the methionine requirement in broiler chicks, humans and farm animals such as lambs4-6. However, a frequently used method to determine the methionine requirement in laying hen is still a traditional one, which involves adding graded levels of methionine to a basal diet deficient in the amino acid7. The methionine requirement for laying hens has been reviewed at several occasions over time and reported to be in a range of 250, 320, 400, 440 and 451 mg hen–1 day–1 and at least 356 mg hen–1 day–1 with variable levels of dietary cystine8-13. Noticeably, Coon and Zhang14 recommended a cystine inclusion of 216 mg hen–1 day–1, while Bregendahl et al.15 reported a higher requirement of 253 mg hen–1 day–1. The National Research Council has reported different methionine requirements for laying hens over time: 280 mg hen–1 day–1 in 1977 and 300 mg hen–1 day–1 in 19941,16. Although, many factors such as protein levels, choline and energy level could affect the methionine requirement of laying hens, the primary reason for the differences among reports can be related to dietary cystine levels. In order to eliminate the confusion on methionine requirement caused by dietary cystine levels, the methionine requirement should be, by definition, the amount of methionine needed only to satisfy the need for biological functions of methionine whereas the cystine function is satisfied by dietary cystine alone12,17,18. Therefore, the methionine requirement of laying hens should be determined with diets sufficient in cystine so that no extra methionine is needed for the biological function of cystine. Moreover, the key metabolites and enzyme activities involved in methionine metabolism (Fig. 1) should be determined in this type of study2. However, no such work has been reported in previous studies on the methionine requirement in laying hens.
A typical experiment for determining the TSAA requirement involves the addition of graded levels of methionine to a basal diet deficient in both methionine and cystine. The deficiency of cystine in test diets is crucial for the determination of the TSAA requirement since extra cystine in test diets can inflate the TSAA requirement19-22. Additionally, the amino acids other than that of interest, should be included in the diet at a rate of at least 90% of the amino acid recommendation23. Currently, there is no information on the efficiency of transsulfuration reactions in laying hens in the literature. Also, cysteine is not the only metabolic outlet for dietary methionine2,3,24-26. The efficiency of converting methionine to cysteine has presented another problem to use the TSAA requirement in practice. Due to the molecular weight difference between methionine and cysteine, the efficiency of utilizing methionine for cysteine on a weight or concentration basis is in the order of 80%, whereas an assumption of 100% molar efficiency is frequently used19-22. Moreover, utilization efficiencies of supplemental methionine and cystine for meeting the TSAA requirement were not equal, especially when the basal diet was limited in methionine27. In order to eliminate the confusion, separation of methionine and cystine requirements is needed. Kalinowski et al.28,29 separately determined the methionine and cystine requirement of slow- and fast-feathering broiler males by using a corn-soybean basal diet. However, no enzymes activities and metabolites involved in methionine and cystine metabolism were determined in the aforementioned study.
The reported experiments herein were conducted to determine digestible methionine and digestible cystine requirements of laying hens for optimum production and to determine the utilization efficiency of supplemental methionine and cystine for the cystine requirement of laying hens. At same time, levels of key metabolites and enzymes activities involved in methionine metabolism were also obtained.
MATERIALS AND METHODS
All procedures regarding the use of live animals in this study were conducted in accordance with the University of Arkansas Institutional Animal Care and Animal Use Protocol #03008.
Experiment 1
Experimental birds, bird care and diets: One hundred and seventy-six 29-week old Dekalb-XL laying hens were randomly assigned to 44 cages (49.9 cm deep and 48.9 cm wide), four hens per cage.
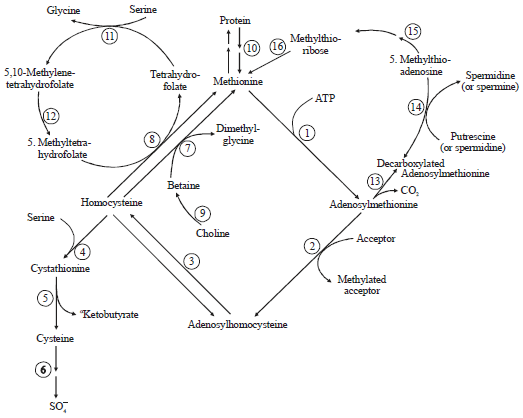 | |
| Fig. 1: | Methionine metabolism in mammals |
The numbers represent the following enzymes or sequences, 1: Methionine adenosyltransferase (EC 2.5.1.6), 2: AdoMet-dependent transmethylations, 3: Adenosylhomocysteinase (EC 3.3.1.1), 4 cystathionine-b-synthase (EC 4.2.1.22), 5: g-cystathionase (EC 4.4.1.1), 6: Further metabolism of cysteine, 7: Betaine-homocysteine methyltransferase (EC 2.1.1.5), 8: Methylfolate-homocysteine methyltransferase (EC 2.1.1.13), 9: Choline+betaine aldehyde dehydrogenases (EC1.1.99.1 and EC1.2.1.8), 10: Equilibrium between free and protein methionine, 11: Serine hydroxymethylase (EC 2.1.2.1), 12: Methylenetetrahydrofolate reductase (EC 1.7.99.5), 13: AdoMet decarboxylase (EC 4.1.1.50), 14: Spermidine (spermine) synthase (EC 2.5.1.16 and EC 2.5.1.22), 15-16: Methylthioadenosine phosphorylase (EC 2.4.2.28)+methionine formation via methylthioribose-1-phosphate. Pathway drawing of methionine and cysteine metabolism is from the publication of Finkelstein2 | |
Hens from four cages received one of the 11 dietary treatments for a six-week period. Feed and water were provided ad libitum. The housing temperature was maintained at 21°C. The hens were provided 15 h of incandescent light (1 foot candle, 6:00 am to 9:00 pm) and a 9-hour dark period (9:00 pm to 6:00 am) each day for the first week of the experiment. The light period of a day was increased 15 min weekly until 16 h light a day was reached.
Two series of diets were mixed with either low cystine (0.148% digestible) or high cystine (0.416% digestible) levels in a basal diet containing 2,850 kcal ME kg–1, 15% CP, 0.143% digestible methionine and 0.148% digestible cystine (Table 1). The basal diet was formulated to be equal to or in excess of NRC1 requirements for all nutrients except methionine and cystine. Additions of methionine and cystine were made to prepare a 0.416% digestible cystine diet series consisting of five levels of digestible methionine (0.143, 0.231, 0.317, 0.407 and 0.495%, respectively) and a 0.148% digestible cystine diet series consisting five levels of digestible methionine (0.143, 0.240, 0.337, 0.434 and 0.531%, respectively). A control diet containing 2,899 kcal ME kg–1 and 19.5% CP (Table 1) was fed to one group of hens. Analysis for percent digestible amino acids for methionine and cysteine in diets was performed as discussed in Ekmay et al.30.
Experiment 2
Experimental birds, bird care and diets: One hundred and sixty 35-week old Dekalb-XL laying hens were randomly assigned to 40 cages (49.9 cm deep and 48.9 cm wide), four hens per cage. Hens from four cages received one of the 10 dietary treatments for a six-week period. Feed and water were provided ad libitum. The housing temperature was maintained at 21°C. The hens were provided 16 h of incandescent light (1 foot candle) (6:00 am to 10:00 pm) and a 8 h dark (10:00 pm to 6:00 am) period each day during the experiment.
Two series of diets were formulated by either adding methionine or cystine to the basal diet containing 0.319% digestible methionine and 0.148% digestible cystine, same as that in Experiment 1 (Table 1) with exception of methionine level. Four levels of equimolar amounts of methionine or cysteine (½ cystine) were added to the basal diet.
| Table 1: | Diet composition of nutrients and ingredients |
 | |
aLimestone, Shell and Bone Builder, had mean particle size of 3260.61 microns and consisted of 39.63% Ca (Shell and Bone Builder, ILC Resources, Des Moines, Iowa). bLimestone, Unical F, had mean particle size of 2166.85 microns and consisted of 39.53 % Ca (Unical F, ILC Resources, Des Moines, Iowa). cThe vitamin premix provided the following per kilogram of diet; Vitamin A: 3,300 IU, Vitamin D3: 3,307 IU, Vitamin E: 40.6 IU, Vitamin K: 1.46 mg, Pantothenic acid: 23.06 mg and Vitamin B12: 0.017 mg. dThe mineral premix provided the following in milligram per kilogram of diet; Mn: 91.88, Fe: 0.027, Zn: 82.65, Cu: 9.48 and I: 1.04. eNonnutritive filler (cellulose), Brown Co., Berlin, NH | |
The added levels were 0.05, 0.10, 0.15 and 0.20% for methionine and 0.04, 0.08, 0.12 and 0.16% for cystine since the molecular weight of cysteine (½ cystine) is 80% of that of methionine. The control diet (Table 1) was also fed to two groups of hens. The objective of the experiment was to study the utilization efficiencies of supplemental methionine and cystine.
Production parameters and sampling: In both Experiments 1 and 2, feed intake and body weight gain were determined every two weeks. Egg production was determined daily. Egg weight was determined with one day's egg collection each week. At the end of each experiment, six hens from each dietary treatment, excluding the control treatment, were sacrificed at 3:00 pm by carotid exsanguination. Immediately after removal, liver samples were frozen and stored in liquid nitrogen for further processing.
Enzyme and metabolite assays: The enzyme extracts were prepared following the procedure introduced by Finkelstein and Martin31. A Tissuemizer® was used at 85% maximum speed for 1 min to homogenize liver samples in 4 volumes of ice-cold 1 mM potassium phosphate, pH 7.5. After the homogenate was centrifuged at 27,000 g for 20 min at 4°C, the supernatant was decanted into Sephadex G-25 columns for the removal of small molecular weight solutes through filtration. The protein-containing fraction was then lyophilized and stored at -40°C. All enzyme assays for each experiment were conducted in less than 10 days, which is the maximum time that enzyme activity has been shown to remain unchanged31,32. Immediately before conducting the enzyme assay, the enzyme extract was reconstituted with water and the final volume was such that 0.5 mL was the equivalent of 1 g of wet tissue. The enzyme activities were determined with the published methods described by Mudd et al.32 for methionine s-adenosyltransferase (EC 2.5.1.6; MAT) and cystathionine ß-synthase (EC 4.2.1.22; CS); by Mudd et al.33 and Finkelstein et al.34 for N5-methyltetrahydrofolate-homocysteine methyltransferase (EC 2.1.1.13; MFMT) and by Finkelstein and Mudd35 with a modification in determining the product formed in the assay36 for betaine-homocysteine methyltransferase (EC 2.1.1.5; BHMT).
The metabolite extract was prepared following the procedure described by Cao and Coon36. Three volumes of 0.2 M HClO4 were added to liver tissue and homogenized using a Tissuemizer® at 85% maximum speed for 1 minute at 4°C. After the homogenate was centrifuged at 27,000 g for 10 min, the supernatant was filtered through a 0.22 μm Millex-GS Millipore filter. The filtrate was treated for blocking thio-group of metabolites37. Fifty μL of 250 mM 2-mercaptoethanol in 700 mM HClO4 and 67 μL of 6 N NaOH were added to 767 μL of filtrate sample. After mixing, 50 μL of 200 mM iodoacetic acid was added to each sample. After 30 min, 66 μL of 6 N HCl was added to each sample. Concentrations of methionine (Met), SAM, SAH, homocysteine and cysteine (Cys) were then analyzed for each treated sample by HPLC37. Due to the reaction of blocking thio-groups of metabolites that occurred due to the presence of 2-mercaptoethanol, the metabolites of oxidized forms could proceed to their corresponding reduced forms. Therefore, the determined concentration was the total amount of metabolite including both oxidized and reduced forms.
Statistical analysis: Data were analyzed using the General Linear Models and one-way analysis of variance (ANOVA) procedures with the help of Statistical Analysis System (SAS38). Differences of p<0.05 were considered statistically significant. Differences between treatment means were compared by Duncan’s Multiple range test. Methionine requirements with high or low cystine levels for laying hens were determined as 95% of the asymptote from the second-order polynomial regression analysis. The cystine requirement was obtained by the difference between two methionine requirements with high or low cystine level.
RESULTS AND DISCUSSION
Experiment 1: The effects of dietary digestible methionine and digestible cystine levels on laying performance determined in Experiment 1 are presented in Table 2. The egg production, egg weight, egg mass, feed intake, feed conversion and body weight gain were significantly influenced by dietary methionine levels. Hens on the high cystine (0.416% digestible) diets, containing 0.317% or greater digestible methionine and hens on the low cystine (0.148% digestible) diets, containing 0.337% or greater digestible methionine, demonstrated an equal amount of egg production to that of hens on the corn-soybean meal control diet. The high cystine (0.416% digestible) diets with variable methionine levels were used to determine the methionine requirement of laying hens since 0.416% digestible cystine is thought to be adequate or in excess for laying hens. Harms and Russell39 reported the commercial layer had a cystine requirement about 175 mg hen–1 day–1 or 3.19 mg for 1 g of egg mass. Schutte et al.40 reported that the cystine requirement of laying hens was 372.2 mg hen–1 day–1 when producing 54.7 g egg mass per hen per day.
Hens fed the high cystine (0.416% digestible) diets showed significant increases of egg production, egg weight, egg mass, feed intake and body weight gain and improved feed conversion when the dietary digestible methionine level was increased from 0.143-0.317% (Table 2). Further increases of dietary methionine levels resulted in non-significant changes of laying performance, as observed in other studies41-43. Polynomial regressions showed the digestible methionine requirement of laying hens fed 0.416% digestible cystine was 354 mg hen–1 day–1 for egg mass (Fig. 2a), 349 mg hen–1 day–1 for feed conversion (Fig. 3a), 437 mg hen–1 day–1 for body weight change (Fig. 4a) and 367 mg hen–1 day–1 for EM+ BWC (Fig. 5a). Results of egg mass, feed efficiency, body weight change and egg mass plus body weight change for hens fed 0.145% digestible cystine are shown in Fig. 2b-5b, respectively. In a meta-analysis study of methionine and cysteine requirements, the predicted values for combined methionine and cysteine requirements were 670 mg day–1 for egg mass and 675 mg day–1 for feed conversion for 95% of maximal hen response44. The digestible methionine requirement, as found in this study, was slightly higher than the previously reported requirements of 0.29-0.32% because of the higher egg mass produced8-10,40,43,45-48. Additionally, the estimated requirement is closer to the reported requirement of 0.36% when 16% of protein is provided in the diet and 0.34% when 17.5% of protein is provided in the diet12,49.
The cystine requirement was obtained by the difference between two methionine requirements with high or low cystine levels. Since the digestible cystine level of 0.148% was approximately 55% of the reported cystine requirement of 0.33%, extra cysteine (½ cystine) should be synthesized from methionine through the transsulfuration reactions40.
| Table 2: | Effects of dietary digestible methionine and cystine levels on laying performance in experiment 1 |
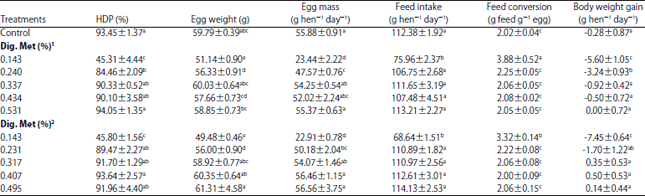 | |
1Diets containing 0.148% digestible cystine. 2Diets containing 0.416% digestible cystine. Each value represents Means±SEM (n = 4 cages). Each cage contained 4 laying hens. a-eMeans within a column with no common superscript differ significantly | |
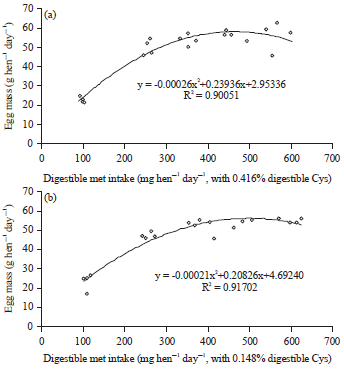 | |
| Fig. 2(a-b): | Polynomial regressions depicting impact of digestible Met intake on egg mass (g hen–1 day–1) in laying hens fed (a) 0.416% and (b) 0.148% digestible Cys in experiment 1 |
Hens fed the low digestible cystine (0.148%) diets showed significant increments in egg production, egg weight, egg mass, feed intake and body weight gain and improved feed efficiency when the dietary digestible methionine level was increased from 0.143-0.337% (Table 2). Further increases of dietary methionine levels resulted in non-significant changes of laying performance in hens. Polynomial regressions showed the digestible methionine requirement of laying hens fed 0.148% digestible cystine for egg mass production was 380 mg hen–1 day–1 with 110 g feed intake day–1 (Fig. 2b). This estimation was 26 mg higher than that of the determined methionine requirement of 354 mg with high cystine diets. It is possible that the extra 26 mg of methionine was used for the synthesis of cysteine (½ cystine) in order to meet the cystine requirement of laying hens. Since the utilization efficiency of methionine for cysteine on a weight or concentration basis is in the order of 80%, the cysteine synthesized from methionine had provided an additional 21 mg cystine to the 0.148% dietary digestible cystine (163 mg cystine intake)19-22. Ultimately, hens required a dietary cystine level of 184 mg hen–1 day–1 for 55.2 g egg mass production. This dietary cystine intake was higher than 0.195% (or 175 mg hen–1 day–1 or 3.19 mg for 1 g of egg mass) as reported by Harms and Russell39.
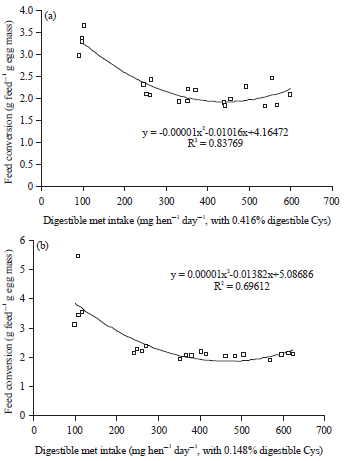 | |
| Fig. 3(a-b): | Polynomial regressions depicting impact of digestible Met intake on feed efficiency in laying hens fed (a) 0.416% and (b) 0.148% digestible Cys in experiment 1 |
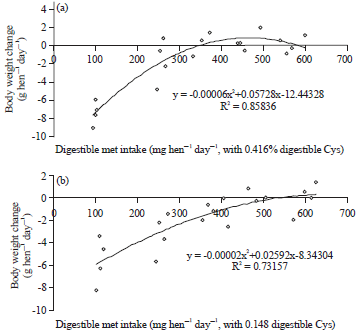 | |
| Fig. 4(a-b): | Polynomial regressions depicting impact of digestible Met intake on body weight change (g hen–1 day–1) in laying hens with (a) 0.416% and (b) 0.148% digestible Cys in experiment 1 |
This could be due to the higher feed intake by hens (110 g versus 90 g hen–1 day–1) than in the previously reported study39. Based on same calculation method, the digestible cystine requirement was 193 mg hen–1 day–1 for feed conversion, 325 mg hen–1 for body weight change, 189 mg hen–1 day–1 for EM+BWC. These results show that the body weight change of laying hens is more sensitive to differences in digestible cystine intake.
The hepatic methionine, cysteine (½ cystine), SAM, SAH and homocysteine concentrations of laying hens were significantly influenced by dietary methionine levels with 0.416 or 0.148% digestible cystine (Table 3).
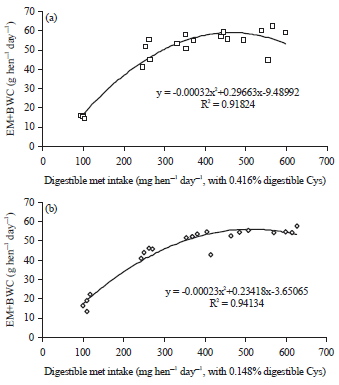 | |
| Fig. 5(a-b): | Polynomial regressions depicting impact of digestible Met intake on egg mass (EM) plus body weight change (BWC) (g hen–1 day–1) in laying hens with (a) 0.416% and (b) 0.148% digestible Cys in experiment 1 |
The hepatic Met/Cys concentration ratio was relatively lower in hens fed the 0.143% digestible methionine diet containing either 0.148 or 0.416% cystine (Fig. 6a and b). The ratio was increased to the highest level when dietary digestible methionine level was increased to 0.240 or 0.231% for the low and high cystine diets, respectively. Further increases of methionine level in diets containing either one of the two cystine levels gradually decreased the Met/Cys ratios.
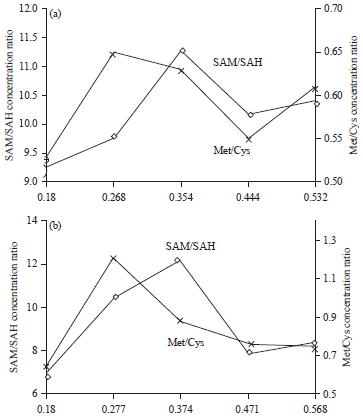 | |
| Fig. 6(a-b): | Effects of dietary methionine level (X-axis, in percent) on methylation and the ratio conversion of methionine to cysteine in laying hen livers fed (a) Cystine sufficient diets (0.450%), (b) Cystine deficient diets (0.182%) |
| Table 3: | Laying hen liver metabolite concentrations (nmoles g–1 wet liver) as affected by dietary digestible methionine and cystine levels in experiment 1 |
 | |
1Diets containing 0.148% digestible cystine. 2Diets containing 0.416% digestible cystine. Each value is Means±SEM (n = 6). a-fMeans within a column with no common superscript differ significantly. Met: Methionine, SAM: s-adenosylmethionine, SAH: s-adenosylhomocysteine, Cys: Cysteine | |
| Table 4: | Laying hen liver enzyme activities (nmoles min–1 g–1 wet liver) as affected by dietary digestible methionine and cystine levels in experiment 1 |
 | |
1Diets containing 0.148% digestible cystine. 2Diets containing 0.416% digestible cystine. Each value is Means±SEM (n = 6). a-dMeans within a column with no common superscript differ significantly. MAT: Methionine s-adenosyltransferase, BHMT: Betaine-homocysteine methyltransferase, MFMT: N5-methyltetrahydrofolate-homocysteine methyltransferase, CS: Cystathionine ß-synthase | |
The changes of the Met/Cys ratio were largely caused by the liver cysteine concentrations (Table 3) due to the significant changes of liver CS activities (Table 4). Although the function of the enzyme BHMT and MFMT is opposite to that of enzyme CS in the methionine metabolism, the higher CS activities observed with layer hens in present experiment on the diets with either extremely deficient or excessive methionine levels was an indication of cysteine/GSH/taurine demand or methionine degradation, respectively. Finkelstein and Martin31,50 reported that at higher levels of CS activity, the methionine metabolism was in favor of transsulfuration reactions than transmethylation reactions even though the BHMT activity was high.
The lower Met/Cys ratio observed with hens fed the diet containing 0.143% digestible methionine with the high or low cystine level was an indication of increased degradation of methionine for cysteine because of the increased CS activity. The elevated degradation of methionine might be stimulated by the increased body protein mobilization as evidenced by the significant body weight loss of hens. Hens fed the 0.143% digestible methionine diets also showed significant reductions of feed intake and laying performance (Table 2), indicating the inadequacy of dietary methionine level. The lower Met/Cys ratios observed with hens fed the diets containing 0.317% or greater digestible methionine were possibly caused by the increased CS activities that degraded the extra dietary methionine, as the hens did not show any significant improvement in laying performance with increased methionine intake. This is consistent with the finding that significantly increased CS activity with age in hatched broiler chicks51.
Table 3 and 4 demonstrate the highest hepatic SAM/SAH concentration ratios for hens fed the diet containing the optimum digestible methionine levels (0.337 and 0.317%, respectively, for the 0.148 and 0.416% digestible cystine diets). Deficient or excessive dietary methionine levels caused reduced liver SAM/SAH concentration ratios. The reduced liver SAM/SAH concentration ratios of hens fed the diets containing deficient levels of methionine (<0.317% digestible) were possibly caused by the decreased available dietary methionine for SAM formation even though the MAT activities were slightly increased (Table 3 and 4). The decreased SAM/SAH ratios of hens fed the diets containing excessive methionine (>0.337% digestible) could be related to the reduced MAT activities although the liver methionine levels were slightly increased. The optimum methionine level in either the low or high cystine diets also resulted in a lack of detectable liver homocysteine concentrations. The cause of the decrease of liver homocysteine might be related to the enhanced methionine utilization for egg protein synthesis.
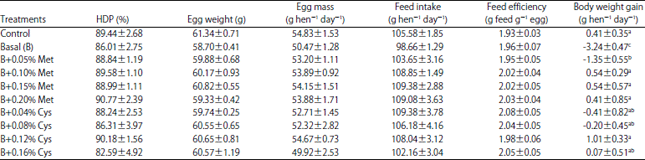
Experiment 2: This experiment was conducted to determine cystine requirement for laying hen and the utilization efficiencies of supplemental methionine and cystine to meet the cystine requirement. The basal diet contained sufficient methionine (0.319% digestible) and deficient cystine (0.148% digestible). The results of the experiment showed significant body weight gain of hens on diets with either methionine or cystine addition level up to 0.12% cystine equivalent. The addition of 0.12% cystine equivalent to the basal diet produced 3.68 g and 4.20 g egg mass per hen per day for hens that received the methionine addition diet and for hens that received the cystine addition diet, respectively. The hens fed the basal diet also showed a feed intake reduction of 10.72 g hen day–1 and a body weight loss of 4.25 g hen–1 day–1 than those of hens on the test diets, indicating the digestible cystine level of 0.148% was deficient.
| Table 5: | Effects of dietary digestible methionine and cystine levels on laying performance in experiment 2 |
 | |
| Each value is Means±SEM (n = 4 cages). Each cage contained 4 laying hens. a-eMeans within a column with no common superscript differ significantly | |
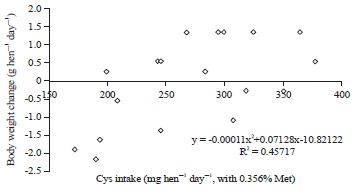 | |
| Fig. 7: | Polynomial regressions depicting impact of Cys intake on body weight change (g hen–1 day–1) in laying hens fed 0.356% Met in experiment 2 |
Polynomial regressions showed the digestible cystine requirement of laying hen with 0.319% digestible methionine (methionine requirement level from Experiment 1) is 276 mg hen–1 day–1 for body weight change in Experiment 2 (Fig. 7). Also, polynomial regressions showed that 157 mg hen–1 day–1 methionine were needed to provide for 113 mg plus 163 mg in basal diet to meet 276 mg hen–1 day–1 cystine requirement for prevent body loss (Fig. 8). Considering so, in this case, conversion efficiency from methionine to cysteine is about 90% (113/156/0.8*100%) (Fig. 9) based on weight or concentration basis.
The results also showed that there was a no significant difference in egg mass production between hens receiving the methionine addition treatment and the hens receiving the cystine addition treatment at each of the four addition levels (Table 5). The lack of change in egg mass also suggests that the cystine requirement of laying hens for optimal production of egg mass could be satisfied with either methionine or cysteine (½ cystine) on an equimolar basis. Since the molecular weight of cysteine is 80% of that of methionine, the utilization efficiency of methionine for cystine is in the order of 80% on a weight and concentration basis. In another study done in monogastric species (pig), the inferior performance of growing pigs on diets with cystine addition to that of pigs on diets with methionine addition was most likely caused by the limited methionine levels in the diets (0.20 and 0.16% for the 30-60 and 60-90 kg body weight ranges, respectively) since the methionine addition could provide pigs both additional methionine and cystine whereas the cystine addition could only provide the pigs with additional cystine27.
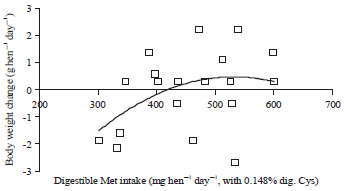 | |
| Fig. 8: | Polynomial regressions depicting impact of Met intake on body weight change (g hen–1 day–1) in laying hens fed 0.148% digestible Cys in experiment 2 |
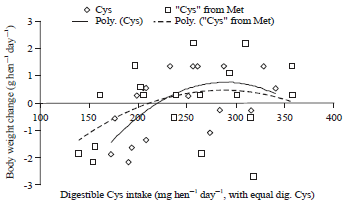 | |
| Fig. 9: | Polynomial regressions depicting impact of Cys intake on body weight change (g hen–1 day–1) in laying hens fed 0.182% Cys in experiment 2 |
This is explained with the irreversible conversion of methionine to cystine in the methionine metabolism2,3.
The hepatic methionine, cysteine, SAM and SAH concentrations of laying hens were significantly influenced by dietary methionine and cystine levels (Table 6).
| Table 6: | Laying hen liver metabolite concentrations (nmoles g–1 wet liver) as affected by dietary digestible methionine and cystine levels in experiment 2 |
 | |
Each value is Means±SEM (n = 6). a-dMeans within a column with no common superscript differ significantly. Met: Methionine, SAM: s-adenosylmethionine, SAH: s-adenosylhomocysteine, Cys: Cysteine | |
| Table 7: | Laying hen liver enzyme activities (nmoles min–1 g–1 wet liver) as affected by dietary digestible methionine and cystine levels in experiment 2 |
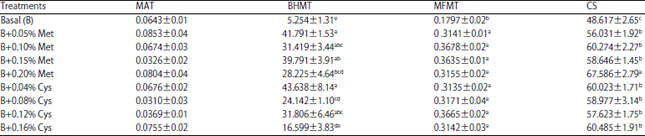 | |
Each value is Means±SEM (n = 6). a-dMeans within a column with no common superscript differ significantly. MAT: Methionine s-adenosyltransferase, BHMT: Betaine-homocysteine methyltransferase, MFMT: N5-methyltetrahydrofolate-homocysteine methyltransferase, CS: Cystathionine ß-synthase | |
Hens fed the diets with methionine additions from 0.05-0.20% compared to the hens fed the basal diet containing 0.319% digestible methionine and 0.148% digestible cystine demonstrated gradual decreases of liver Met/Cys ratios from 0.98-0.70. The decreased Met/Cys ratios were caused by the increased liver cysteine concentrations due to the elevated CS activities (Table 7). The hens fed the diets with the methionine addition also showed increase in liver enzyme activities of BHMT and MFMT greater than those of hens fed the basal diet (Table 7). Methionine additions of more than 0.05% also showed a non-significant change in liver methionine level and MAT activity. The hens fed the diet with 0.05% methionine also showed an increase in liver SAM/SAH concentration ratio by 3.07 units from that of the value of 7.25 of hens fed the basal diet. Further increase of methionine addition level resulted in a non-significant change in SAM/SAH ratio. These observations generally agreed with the results from Experiment 1.
The cystine addition compared to the methionine addition to the basal diet produced similar changes of liver Met/Cys and SAM/SAH ratios in laying hens. The liver metabolite concentrations and enzyme activities of hens showed the same changes for the cystine and methionine addition treatments, with an exception that there was a significant increase of liver methionine concentration of hens fed the diets with an increased level of cystine. Additionally, there was a significant decrease of liver methionine concentration of hens fed the diets with an increased level of methionine in diets. The increased liver methionine concentration of hens fed the diets with an increased level of dietary methionine most likely resulted from the increased need for methionine degradation.
Although, the current findings are crucial to better understanding the utilization efficiencies of methionine and cystine in laying hens, further research should be conducted in order to create a more representative database. The digestible methionine and cystine requirements are reflective of genetics, gender, age and performance. Moreover, the hepatic enzymes and metabolites of total sulfur amino acids experience daily variations throughout light and dark periods52. Each of the aforementioned variables represents the need for additional experimentation.
CONCLUSION
Overall, the study demonstrates that the utilization efficiencies of methionine and cysteine (½ cystine) were 100% on an equimolar basis for egg mass and 90% on an equimolar basis to prevent body weight loss. When methionine is used to meet the cystine requirement, the utilization efficiency of 80% should be adequate on a weight and concentration basis for egg mass and 72% for body weight maintenance. The practice of feeding ingredients with a substantial digestible cystine level for supporting body weight may be beneficial for laying hens.
SIGNIFICANCE STATEMENT
This study discovers the utilization efficiencies of methionine and cysteine for maintaining egg mass and preventing body weight loss in laying hens. This study will help the researcher to understand the digestible methionine and cystine requirements for optimum production and to determine the utilization efficiency of supplemental methionine and cystine for the cystine requirement of laying hens.
REFERENCES
- National Research Council, 1994. Nutrient Requirements of Poultry. 9th Revised Edn., The National Academies, Washington, DC, 176.
CrossRefDirect Link - Finkelstein, J.D., 1990. Methionine metabolism in mammals. J. Nutr. Biochem., 1: 228-237.
CrossRefDirect Link - Finkelstein, J.D., 1998. The metabolism of homocysteine: Pathways and regulation. Eur. J. Pediatr., 2: S40-S44.
CrossRefPubMedDirect Link - Chamruspollert, M., G.M. Pesti and R.I. Bakalli, 2002. Determination of the methionine requirement of male and female broiler chicks using an indirect amino acid oxidation method. Poult. Sci., 81: 1004-1013.
Direct Link - Kurpad, A.V., M.M. Regan, S. Varalakshmi, J. Vasudevan, J. Gnanou, T. Raj and V.R. Young, 2003. Daily methionine requirements of healthy Indian men, measured by a 24-h indicator amino acid oxidation and balance technique. Am. J. Clin. Nutr., 77: 1198-1205.
CrossRefDirect Link - Wei, G., L. Chen, G. Xinmei, Z. Fan, C. Daofu and L. Chenli, 2017. Investigation of the postruminal methionine requirement of growing lambs by using the indicator amino acid oxidation technique. Anim. Feed Sci. Technol., 228: 83-90.
CrossRefDirect Link - Harms, R.H., 2001. What levels of amino acids should a feed contain for the commercial laying hen? J. Applied Anim. Res., 19: 1-24.
CrossRefDirect Link - Harms, R.H. and B.L. Damron, 1969. Protein and sulfur amino acid requirement of the laying hen as influenced by dietary formulation. Poult. Sci., 48: 144-149.
CrossRefDirect Link - Novacek, E.J. and C.W. Carlson, 1969. Low protein cage-layer diets and amino acids. Poult. Sci., 48: 1490-1497.
CrossRefDirect Link - Schutte, J.B. and E.J.V. Weerden, 1978. Requirement of the hen for sulphur‐containing amino acids. Br. Poult. Sci., 19: 573-581.
CrossRefPubMedDirect Link - Schutte, J.B., J. de Jong and H.L. Bertram, 1994. Requirement of the laying hen for sulfur amino acids. Poult. Sci., 73: 274-280.
CrossRefPubMedDirect Link - Calderon, V.M. and L.S. Jensen, 1990. The requirement for sulfur amino acid by laying hens as influenced by the protein concentration. Poult. Sci., 69: 934-944.
CrossRefDirect Link - Strathe, A.B., A. Lemme, J.K. Htoo and E. Kebreab, 2011. Estimating digestible methionine requirements for laying hens using multivariate nonlinear mixed effect models. Poult. Sci., 90: 1496-1507.
CrossRefDirect Link - Coon, C.N. and B. Zhang, 1999. Ideal amino acid profile for layers examined. Feedstuffs, 71: 13-15, 31.
Direct Link - Bregendahl, K., S.A. Roberts, B. Kerr and D. Hoehler, 2008. Ideal ratios of isoleucine, methionine, methionine plus cystine, threonine, tryptophan and valine relative to lysine for white leghorn-type laying hens of twenty-eight to thirty-four weeks of age. Poult. Sci., 87: 744-758.
CrossRefDirect Link - Harms, R.H., N. Ruiz, D. Milesr, 1990. Research note: Conditions necessary for a response by the commercial laying hen to supplemental choline and sulfate. Poult. Sci., 69: 1226-1229.
CrossRefDirect Link - Harms, R.H. and G.B. Russell, 1998. Layer performance when returned to a practical diet after receiving an amino acid-deficient diet. J. Applied Poult. Res., 7: 175-179.
CrossRefDirect Link - Creek, R.D., 1968. Non equivalence in mass in the conversion of phenylalanine to tyrosine and methionine to cystine. Poult. Sci., 47: 1385-1386.
CrossRefDirect Link - Graber, G. and D.H. Baker, 1971. Sulfur amino acid nutrition of the growing chick: Quantitative aspects concerning the efficacy of dietary methionine, cysteine and cystine. J. Anim. Sci., 33: 1005-1011.
CrossRefDirect Link - Sowers, J.E., W.L. Stockland and R.J. Meade, 1972. L-methionine and L-cystine requirements of the growing rat. J. Anim. Sci., 35: 782-788.
CrossRefDirect Link - Stockland, W.L., R.J. Meade, D.F. Wass and J.E. Sowers, 1973. Influence of levels of methionine and cystine on the total sulfur amino acid requirement of the growing rat. J. Anim. Sci., 36: 526-530.
CrossRefDirect Link - Schutte, J.B. and J. de Jong, 1996. Biological efficacy of DL-methionine hydroxy analog-free acid compared to DL-methionine in broiler chicks as determined by performance and breast meat yield. Agribiol. Res., 49: 74-82.
Direct Link - Benevenga, N.J., 1984. Evidence for alternative pathways of methionine catabolism. Adv. Nutr. Res., 110: 1-18.
CrossRefDirect Link - Livesey, G., 1984. Methionine degradation: ‘anabolic and catabolic’. Trends Biochem. Sci., 9: 29-29.
CrossRefDirect Link - Stipanuk, M.H., 2004. Sulfur amino acid metabolism: Pathways for production and removal of homocysteine and cysteine. Annu. Rev. Nutr., 24: 539-577.
CrossRefDirect Link - Roth, F.X. and M. Kirchgessner, 1987. Biological efficiency of dietary methionine or cystine supplementation with growing pigs. J. Anim. Physiol. Anim. Nutr., 58: 267-280.
CrossRefDirect Link - Kalinowski, A., E.T. Jr. Moran and C. Wyatt, 2003. Methionine and cystine requirements of slow- and fast-feathering male broilers from zero to three weeks of age. Poult. Sci., 82: 1423-1427.
Direct Link - Kalinowski, A., E.T. Moran and C.L. Wyatt, 2003. Methionine and cystine requirements of slow- and fast-feathering broiler males from three to six weeks of age. Poult. Sci., 82: 1428-1437.
CrossRefDirect Link - Ekmay, R.D., M. De Beer, S.J. Mei, M. Manangi and C.N. Coon, 2013. Amino acid requirements of broiler breeders at peak production for egg mass, body weight and fertility. Poult. Sci., 92: 992-1006.
CrossRefDirect Link - Finkelstein, J.D. and J.J. Martin, 1984. Methionine metabolism in mammals. Distribution of homocysteine between competing pathways. J. Biol. Chem., 259: 9508-9513.
PubMed - Mudd, S.H., J.D. Finkelstein, F. Irreverre and L. Laster, 1965. Transsulfuration in mammals: Microassays and tissue distribution of three enzymes of the pathway. J. Biol. Chem., 240: 4382-4392.
Direct Link - Mudd, S.H., H.L. Levy and G. Morrow, 1970. Deranged B12 metabolism: Effects on sulfur amino acid metabolism. Biochem. Med., 4: 193-214.
CrossRefDirect Link - Finkelstein, J.D., W.E. Kyle and B.J. Harris, 1971. Methionine metabolism in mammals. Regulation of homocysteine methyltransferases in rat tissue. Arch. Biochem. Biophys., 146: 84-92.
CrossRefDirect Link - Finkelstein, J.D. and S.H. Mudd, 1967. Transsulfuration in mammals. The methionine sparing effect of cystine. J. Biol. Chem., 242: 873-880.
Direct Link - Cooper, J.D.H. and D.C. Turnell, 1982. Fluorescence detection of cystine by o-phthalaldehyde derivatisation and its separation using high-performance liquid chromatography. J. Chromatogr. B: Biomed. Sci. Applic., 227: 158-161.
CrossRefDirect Link - Harms, R.H. and G.B. Russell, 1996. Evaluation of the cystine requirement of the commercial laying hen. J. Applied Poult. Res., 5: 139-149.
CrossRefDirect Link - Schutte, J.B., E.J.V. Weerden and H.L. Bertram, 1984. Protein and sulphur amino acid nutrition of the hen during the early stage of laying. Arch. Geflugelk, 48: 165-170.
Direct Link - Bertram, H.L., J.B. Schutte and J. de Jong, 1995. Influence of DL-methionine supplements on the performance of laying hens under heat stress. Arch. Geflügelk., 59: 314-318.
Direct Link - Rama Rao, S.V., V. Ravindran, T. Srilatha, A.K. Panda and M.V.L.N. Raju, 2011. Effect of dietary concentrations of energy, crude protein, lysine and methionine on the performance of White Leghorn layers in the tropics. J. Applied Poult. Res., 20: 528-541.
CrossRefDirect Link - Saki, A.A., R.N. Harsini, M.M. Tabatabaei, P. Zamani and M. Haghight, 2012. Estimates of methionine and sulfur amino acid requirements for laying hens using different models. Braz. J. Poult. Sci., 14: 159-232.
CrossRefDirect Link - Harms, R.H., B.L. Damron and P.W. Waldroup, 1966. Evaluation of the sulfur amino acid requirements of commercial egg production type pullets. Poult. Sci., 46: 181-186.
Direct Link - Reid, B.L. and C.W. Weber, 1973. Dietary protein and sulfur amino acid levels for laying hens during heat stress. Poult. Sci., 52: 1335-1343.
CrossRefDirect Link - Schutte, J.B., E.J.V. Weerden and H.L. Bertram, 1983. Sulphur amino acid requirement of laying hens and the effects of excess dietary methionine on laying performance. Br. Poult. Sci., 24: 319-326.
CrossRefDirect Link - Finkelstein, D. and J.J. Martin, 1986. Methionine metabolism in mammals. Adaptation to methionine excess. J. Biol. Chem., 261: 1582-1587.
PubMed - Cao, T., J.T. Weil, P. Maharjan, J. Lu and C.N. Coon, 2020. The diurnal changes of hepatic enzymes and metabolites of methionine metabolism in laying hens. Int. J. Poult. Sci., 19: 210-218.
CrossRefDirect Link