
Nermeen M. Ismail
Department of Newcastle Disease, Veterinary Serum and Vaccine Research Institute, Abbassia, Cairo, Egypt
Ayman H. El-Deeb
Department of Virology, Faculty of Veterinary Medicine, Cairo University, Giza, Egypt
Mohamed M. Emara
Department of Virology, Faculty of Veterinary Medicine, Cairo University, Giza, Egypt
Hoda I. Tawfik
Department of Newcastle Disease, Veterinary Serum and Vaccine Research Institute, Abbassia, Cairo, Egypt
Nabil Abdel Wanis
Department of Newcastle Disease, Veterinary Serum and Vaccine Research Institute, Abbassia, Cairo, Egypt
Hussein A. Hussein
Department of Virology, Faculty of Veterinary Medicine, Cairo University, Giza, Egypt
International Journal of Poultry Science
Year: 2018 | Volume: 17 | Issue: 4 | Page No.: 167-174







ABSTRACT
Background and Objective: Avian influenza and Newcastle disease viruses are a continuous threat to poultry industry. Vaccines are increasingly used to control these viral infections. This study aimed to develop efficacious vaccines and vaccination strategies against avian influenza and Newcastle disease viruses. Methodology: Two formulations of bivalent vaccines for avian influenza and Newcastle disease viruses were prepared based on the use of IMS1313-nanoparticles (mucosal vaccine) and Montanide ISA71 (parental vaccine) adjuvants. The prepared vaccines were delivered in specific pathogen free (SPF) chickens with different vaccination protocols. Cell mediated and humoral immune response (cytokine expression levels including IFN-γ and IL-6, lymphocyte proliferation, antibodies titers against both H5N1 and NDV) were measured. Challenge trial was carried out to determine the protection percent and shedding pattern of the challenged viruses. Results: Results of the present study revealed a significant increase of IFN-γ and IL-6 genes expression and lymphocytes proliferation in the vaccinated groups compared to the unvaccinated group. Two applications of the mucosal vaccine demonstrated higher hemagglutination inhibition (HI) titers and protection percentage ranged from 40-50% with different levels of virus shedding as measured by qRT-PCR assay. However, when the vaccines were applied in a prime-boost protocol (mucosal-parental, respectively), protection reached 90 and 100% against avian influenza virus (AIV) and Newcastle disease virus (NDV), respectively. No shedding of the NDV-challenge virus was detected whereas, AIV-challenge virus was detected in the samples of the 3rd day post-challenge. Conclusion: Indeed, the use of mucosal-parental vaccines in a prime-boost vaccination protocol demonstrated the potentiality of such approach.
PDF Abstract XML References Citation
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Nermeen M. Ismail, Ayman H. El-Deeb, Mohamed M. Emara, Hoda I. Tawfik, Nabil Abdel Wanis and Hussein A. Hussein, 2018. IMS 1313-nanoparticle Mucosal Vaccine Enhances Immunity Against Avian Influenza and Newcastle Disease Viruses. International Journal of Poultry Science, 17: 167-174.
DOI: 10.3923/ijps.2018.167.174
URL: https://scialert.net/abstract/?doi=ijps.2018.167.174
DOI: 10.3923/ijps.2018.167.174
URL: https://scialert.net/abstract/?doi=ijps.2018.167.174
INTRODUCTION
Avian influenza virus (AIV) and Newcastle disease virus (NDV) both cause illness in birds (e.g., chickens), leading to severe economic losses in the poultry industry. The epidemiology of AIV infections has changed over the past two decades due to the spread of highly pathogenic AIVs, such as H5N1, in domestic poultry together with the spread of the Chinese VIId form of NDV in the Middle East and Egypt1,2. Therefore, control of highly pathogenic AIVs and NDV in poultry is now a global issue. In Egypt, vaccination is a major AIV and NDV control strategy, with several commercial inactivated vaccines being licensed for both diseases3. Most of the available vaccines are based on the use of adjuvant(s) with whole inactivated virions prepared either from wild-type or reverse-genetics viruses4. However, presently, more than 95% of AIV vaccines are adjuvant-based, inactivated products5 that provide incomplete protection in the field. Furthermore, widespread and prolonged use of inactivated AIV vaccines promotes emergence of antigenic variants against which vaccines are ineffective6,7. Therefore, the efficacy of inactivated AIV vaccines requires enhancement.
An effective way to target antigens before infection is to induce mucosal immunity at the replication site of a variety of viruses, including AIV and NDV8,9. In contrast to systemic influenza, mucosal vaccination induces both mucosal and serum antibodies, which in most cases, cross-react with antigenically different influenza viruses10-12. In addition, the local mucosal immune response plays an important role in developing protection against NDV by inhibiting virus multiplication at the viruses portal of entry13,14. Vaccination with an inactivated mucosal NDV and AIV vaccine is of particular interest because it has the advantage of inducing both secretory immunoglobulin A (IgA) in mucosal compartments and immunoglobulin G (IgG) in serum, while parenteral vaccines only induce serum IgG1,15. Nevertheless, mucosal immunity induced by an inactivated virus is ineffective since the inactivated virus loses its ability to replicate in the epithelium of the respiratory tract and, therefore, its ability to induce an effective immune response16.
Consequently, use of adjuvants is necessary with mucosal vaccines in order to be efficient17. As efficient mucosal adjuvants have been developed to induce mucosal immunity18, current efforts are focused on mucosal vaccination strategies that induce protective immunity in both compartments19. Water-in-oil emulsions may not be compatible with vaccinations targeted to diverse mucosal epithelial as the continuous phase of these vaccine formulations is oil. However, the adjuvant MontanideTM IMS 1313 N VGPR (IMS 1313) consists of a water-dispersed liquid nanoparticle combined with an immuno-stimulating compound. Because this adjuvant has an aqueous phase, it is suitable as a mucosal delivery vehicle and can also be used for mass vaccination in the poultry industry20,21.
The current study aimed to assess the immune response(s) following ocular vaccination of bivalent AIV and NDV in combination with the adjuvant IMS 1313. Vaccine efficacy was evaluated by measuring different cytokines [interferon (IFN)-γ and interleukin (IL)-6], lymphocyte proliferation and antibody titers in serum post-vaccination (p.v.). Chicken survival rates and viral shedding in tracheal secretions of vaccinated chickens were assessed post-challenge.
MATERIALS AND METHODS
Viruses: Reassortant AIV strain A/Chicken/Egypt/Q1995D/ 2010(H5N1) was provided by the National Research Center (Cairo, Egypt) and used for vaccine preparation. Influenza A virus [A/chicken/Egypt/VSVRI/2009(H5N1)] was used as a challenge virus and kindly provided by the Veterinary Serum and Vaccine Research Institute (VSVRI, Cairo, Egypt). A NDV (NDV-B7-RLQP-CH-EG-12) was also used for vaccine preparation and as a challenge virus. All of the viruses were propagated, titrated and inactivated with binary ethylamine according to King22.
Adjuvants and vaccines: The IMS 1313 and ISA 71 adjuvants provided by SEPPIC (Puteaux, France) were used for vaccine formulation as per manufacturer instructions.
Experimental design: A total of 125, 1 day old SPF chicks were purchased from the specific pathogen-free egg project of Kom Oshim (El Fayoum Governorate, Egypt). Chickens were randomly divided into five groups (n = 25 birds/group). Group 1 chicks received 0.2 mL of the IMS 1313-based bivalent AIV and NDV vaccine intraocularly. Group 2 chicks were primed and boosted intraocularly with the same dose of the mucosal IMS 1313 AIV/NDV vaccine at 10 and 20 days old. Group 3 chicks were primed intraocularly with 0.2 mL of the mucosal IMS 1313 AIV/NDV vaccine at 10 days old and subcutaneously boosted with 0.5 mL of the inactivated ISA 71 oil-based AIV/NDV vaccine at 20 days old. Group 4 chicks were subcutaneously vaccinated with 0.5 mL of the inactivated ISA 71 oil-based AIV/NDV vaccine and group 5 chicks were kept unvaccinated as control.
Twenty-one days p.v. chickens were challenged via oculonasal route with a 106.0 egg infectious dose. On days 3, 7, 10 and 14 post-challenge, tracheal swabs were collected to quantify viral shedding by quantitative Reverse Transcriptase-Polymerase Chain Reaction (qRT-PCR) using specific primer and probe sequences for AIV and NDV23,24. Survival of challenged chickens was monitored for 14 days to determine the percentage of protection elicited by each vaccination regimen. Serum samples were collected on a weekly basis for 12 weeks p.v. from unchallenged birds for detection of serum antibodies by hemagglutination inhibition (HI) assay. Heparinized blood samples from chickens of each group were drawn for quantification of cytokines IFN-γ and IL-6, mRNA expression by qRT-PCR and measurement of lymphocyte proliferation.
Hemagglutination inhibition assay: Serum samples were tested weekly by hemagglutination inhibition assay according to standard procedures of the Office International des Epizooties (OIE)2.
qRT-PCR: The RNA extraction was done with Qiagen RNA Purification kits (Qiagen Inc., Valencia, CA, USA) and qRT-PCR runs were performed using Stratagene MX3000 thermocyclers. The PCR cycling profile for cytokine measurements was completed according to Suzuki et al.23 and viral RNA amplification PCR was done according to Londt et al.24 and Wise et al.25.
Lymphocyte proliferation assay: Lymphocyte proliferation assay was carried out using Cell Proliferation Kit II (XTT) that was purchased from Sigma-Aldrich. The assay was conducted according to the manufacturer’s instructions.
Statistical analysis: Cellular immune responses were analyzed statistically using a Fischer exact probability test at p<0.05, while humoral immune responses were statistically analyzed using a Duncan multiple range test at p<0.05 using SPSS version 22 software.
RESULTS
Hemagglutination inhibition titers in sera of vaccinated chickens: Antibody titers against NDV showed a significant increase 2 weeks p.v. in group 2 chickens compared to group 1. However, specific NDV antibody titers in group 3 chickens were significantly higher (8.2 log 2) than those in the other vaccinated groups (Fig. 1). Antibody titers against H5N1 in sera samples collected from group 3 showed significantly enhanced H5N1 antibody titers 2 weeks p.v. compared to the other vaccinated groups (6.4 log 2, Fig. 2).
IFN-γ> expression levels: The qRT-PCR was used to determine IFN-γ mRNA expression levels in chickens from peripheral blood mononuclear cells (PBMC) 1 and 2 weeks p.v. Seven days p.v., group 3 chickens demonstrated significantly higher IFN-γ expression than the other groups at all tested time points (Fig. 3).
IL-6 expression levels: The results of IL-6 expression levels as measured by qRT-PCR revealed that vaccinated groups showed a significant increase in IL-6 expression compared to the unvaccinated group. Among the results of vaccinated groups, group 3 had the highest IL-6 expression, with 10.27- and 17.15-fold changes at 1 and 2 weeks p.v., respectively (Fig. 4).
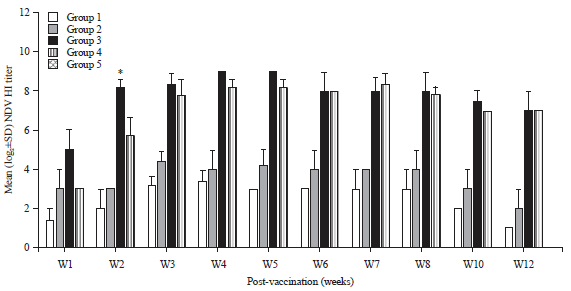 | |
| Fig. 1: | Mean serum antibody titers to Newcastle disease virus (NDV) in different chicken groups |
*Significant p-value<0.05 | |
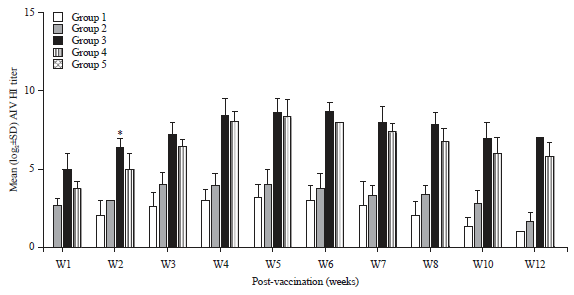 | |
| Fig. 2: | Mean serum antibody titers to avian influenza virus (AI) in different chicken groups |
*Significant p-value<0.05 | |
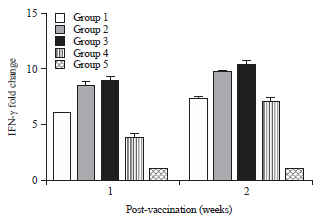 | |
| Fig. 3: | IFN-γ mRNA expression levels in chicken groups at 1 and 2 weeks post vaccination measurement was done by qRT-PCR |
Data shows fold change compared with uninfected control chickens. *Significant p-value<0.05 | |
Lymphocyte proliferation: Significant increases in lymphocyte proliferation were found 3 days p.v. in all vaccinated groups compared to the control unvaccinated group (Fig. 5). Fourteen days p.v., group 3 showed significantly higher (1.7-fold) results relative to other vaccinated groups. The current study revealed that lymphocyte proliferation significantly increased 3 days p.v. in all vaccinated groups compared to the unvaccinated controls.
Protection percent: Velogenic NDV genotype VIId and highly pathogenic H5N1 viral challenges revealed discrepancies in the percentage of vaccination protection, ranging from 20-50% after NDV-challenge and 0-40% after AIV-challenge when the mucosal vaccine was delivered intraocularly in one or two doses, respectively.
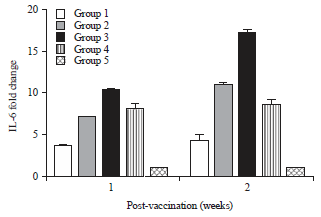 | |
| Fig. 4: | IL-6 mRNA expression levels in chicken groups at 1 and 2 weeks post vaccination measurement was done by qRT-PCR |
Data shows fold change compared with uninfected control chickens. *Significant p-value<0.05 | |
The parental vaccine (group 4) alone induced 80% protection against both viral challenges. However, the protection reached 90 and 100% against H5N1 and NDV challenges, respectively, when the prepared vaccines were used as a mucosal prime followed by parental booster (Fig. 6a, b).
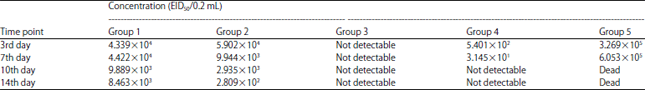
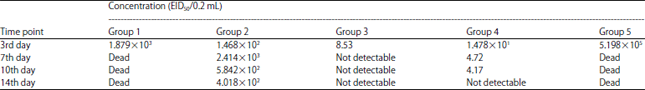
H5N1 and NDV shedding post-challenge: No NDV shedding was detected in samples from group 3. Meanwhile, group 2 results revealed 1 and 2 log reductions in viral shedding compared to the unvaccinated group (Table 1). No H5N1 viral shedding was found in group 3 samples 7 days post-challenge and reduced shedding was found in samples collected from group 2 compared to the unvaccinated group (Table 2).
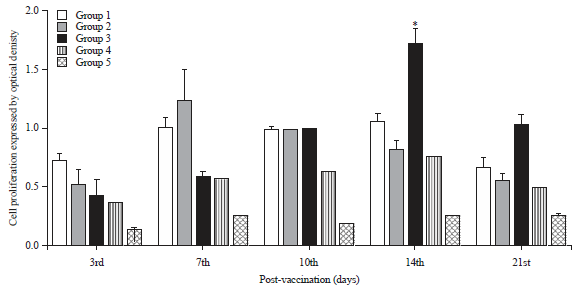 | |
| Fig. 5: | Mean optical density of lymphocytes proliferation, isolated from peripheral blood of chickens in each group |
*Significant at p<0.05 | |
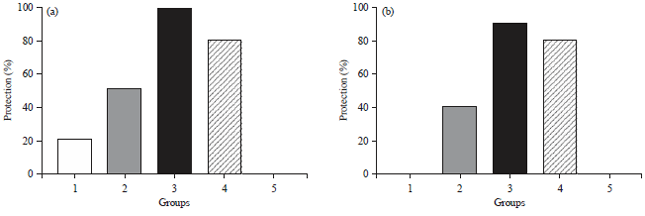 | |
| Fig. 6(a-b): | Protection percentage of the vaccinated and challenged chicken groups (a) Against the velogenic NDV-genotype VIId and (b) Against the A/chicken/Egypt/VSVRI/2009(H5N1) |
| Table 1: | Shedding amount post-challenge with NDV |
 | |
| Table 2: | Shedding amount post challenge with H5N1 |
 | |
DISCUSSION
In the present study, IMS 1313-nanoparticles (mucosal) and ISA 71 (parental) adjuvants were used as immuno-potentiators to enhance the immune response. Current hemagglutination inhibition assay results showed that two intraocular mucosal doses of the IMS 1313 based-vaccine significantly enhanced serum antibody responses in chickens against H5N1 and NDV 4 weeks p.v. relative to single mucosal dose and control groups. These results are consistent with those obtained by Hikono et al.1. On the other hand, the parental inactivated bivalent vaccine based on ISA 71 as an adjuvant induced protective antibody responses after a single subcutaneous injection while at least two intraocular vaccinations were needed to obtain 50% protection. The obtained results are in agreement with the previously published study of Sasaki et al.26. However, combining the mucosal-parental vaccines in a prime-boost vaccination regime induced the highest levels of specific antibody titers against H5N1 and NDV.
The IgG production in responding lymphoid tissues was weaker following mucosal vaccination compared to parental vaccination, which may be attributed to the greater efficiency of antigen delivery to the immune system and tissue trauma associated with needle injection27. Meanwhile, the response to mucosal vaccination includes a substantial IgA component, which was essentially absent in the subcutaneous vaccination response. The Harderian gland is an essential organ for the development of immune responses in poultry. Thus, intraocular vaccinations result in significant increases in plasma cells in sections of the Harderian gland, resulting in production of necessary local antibodies. Specific antibodies against AIV have been previously reported by others in chicken tears after intraocular immunization28,29.
Furthermore, Harderian glands affect antibody-mediated immune responses in other mucosal sites as Harderian gland-derived IgA-B cells have been detected in cecal tonsils30. Additionally, secretory IgA can reduce viral adhesion and capture viruses that invade the mucosa31. In the present study, the percentage of protection with two doses of the mucosal vaccine ranged from 40-50% against AIV and NDV challenges, respectively. However, 80% protection was obtained when the parental ISA 71 vaccine was given alone. Nevertheless, protection reached 90% against the AIV challenge and 100% against NDV, with no shedding of either viruses in chickens that were initially vaccinated mucosally and then given a parental booster. These findings are in accordance with Lee et al.32.
Cytokines, such as proinflammatory IL-6 and IFN-γ, released by lymphocytes after antigen stimulation can regulate immune responses and play important roles in mucosal immuninty33,34. The IFN-γ plays a vital role in macrophage activation and modulation of the cellular immune system in addition to its antiviral activity35. Measuring IFN-γ expression in the present study revealed significant upregulation of IFN-γ in all vaccinated groups compared to unvaccinated chickens. However, mucosal prime-parental boost vaccinated chickens expressed the highest levels of IFN-γ compared to other vaccinated groups.
Herein, the highest levels of proinflammatory IL-6 expression belonged to the chickens that received the mucosal-parental vaccine regimen compared to other groups. These results support those reported by others previously36,37. Enhanced IL-6 expression has been shown to induce proliferation of B cells and promote secretion of IgA16. The role of cell-mediated immunity in protecting against AIV and NDV is limited. T-Lymphocytes are the most important cells mediating the cellular immune response and T-cell subpopulations with diverse functions have been identified in chickens38.
CONCLUSION
Taken together, implementation of the current mucosal prime-parental boost vaccination strategy is able to confer broader immunity against both AIV and NDV in chickens and should be considered in other poultry.
SIGNIFICANCE STATEMENT
This study reports the use of mucosal nanoparticle immune-stimulating complex in the formulation of bivalent reassortant avian influenza and Newcastle disease vaccine in a prime boost strategy. Also, the manuscript presents the efficacy of one and two doses of the mucosal vaccine. Results of prime boost strategy proved the beneficial effects of such application in controlling infections by both viruses specially the induction of immunity at the site of replication. This study will help the researchers to extend the researches on the mucosal vaccine formulation using different adjuvants to control the spread of such viruses at the site of replication post infection. Thus, the manuscript recommend the wide application of prime boost vaccination strategy based on mucosal followed by parental vaccines.
ACKNOWLEDGMENT
The authors would like to thank the Veterinary Serum and Vaccine Research Institute (VSVRI) Cairo, Egypt for all kinds of support.
REFERENCES
- Hikono, H., M. Mase, A. Matsuu, M. Nakayama and T. Saito, 2013. Intraocular vaccination with an inactivated highly pathogenic avian influenza virus induces protective antibody responses in chickens. Vet. Immunol. Immunopathol., 151: 83-89.
CrossRefDirect Link - OIE., 2015. Avian Influenza. In: Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, OIE (Eds.). Chapter 2.3.4, Office International des Epizooties, Paris, France, pp: 1-23.
Direct Link - Khan, T.A., C.A. Rue, S.F. Rehmani, A. Ahmed, J.L. Wasilenko, P.J. Miller and C.L. Afonso, 2010. Phylogenetic and biological characterization of Newcastle disease virus isolates from Pakistan. J. Clin. Microbiol., 48: 1892-1894.
CrossRefDirect Link - Kandeil, A., Y. Moatasim, M.R. Gomaa, M.M. Shehata and R. El-Shesheny et al., 2016. Generation of a reassortant avian influenza virus H5N2 vaccine strain capable of protecting chickens against infection with Egyptian H5N1 and H9N2 viruses. Vaccine, 34: 218-224.
CrossRefDirect Link - Lone, N.A., E. Spackman and D. Kapczynski, 2017. Immunologic evaluation of 10 different adjuvants for use in vaccines for chickens against highly pathogenic avian influenza virus. Vaccine, 35: 3401-3408.
CrossRefDirect Link - Kim, J.K., G. Kayali, D. Walker, H.L. Forrest and A.H. Ellebedy et al., 2010. Puzzling inefficiency of H5N1 influenza vaccines in Egyptian poultry. Proc. Natl. Acad. Sci. USA., 107: 11044-11049.
CrossRefDirect Link - Cattoli, G., L. Susta, C. Terregino and C. Brown, 2011. Newcastle disease: A review of field recognition and current methods of laboratory detection. J. Vet. Diagn. Invest., 23: 637-656.
CrossRefPubMedDirect Link - Zhang, L., M. Zhang, J. Li, T. Cao, X. Tian and F. Zhou, 2008. Enhancement of mucosal immune responses by intranasal co-delivery of Newcastle disease vaccine plus CpG oligonucleotide in SPF chickens in vivo. Res. Vet. Sci., 85: 495-502.
CrossRefDirect Link - Seo, K.Y., S.J. Han, H.R. Cha, S.U. Seo, J.H. Song, S.H. Chung and M.N. Kweon, 2010. Eye mucosa: An efficient vaccine delivery route for inducing protective immunity. J. Immunol., 185: 3610-3619.
CrossRefDirect Link - Yoshikawa, T., K. Matsuo, K. Matsuo, Y. Suzuki and A. Nomoto et al., 2004. Total viral genome copies and virus-Ig complexes after infection with influenza virus in the nasal secretions of immunized mice. J. Gen. Virol., 85: 2339-2346.
CrossRefDirect Link - Hasegawa, H., T. Ichinohe, S.I. Tamura and T. Kurata, 2007. Development of a mucosal vaccine for influenza viruses: Preparation for a potential influenza pandemic. Expert Rev. Vaccines, 6: 193-201.
CrossRefDirect Link - Hong, S.H., Y.H. Byun, C.T. Nguyen, S.Y. Kim and B.L. Seong et al., 2012. Intranasal administration of a flagellin-adjuvanted inactivated influenza vaccine enhances mucosal immune responses to protect mice against lethal infection. Vaccine, 30: 466-474.
CrossRefPubMedDirect Link - Al-Garib, S.O., A.L.J. Gielkens, E. Gruys and G. Kochi, 2003. Review of Newcastle disease virus with particular references to immunity and vaccination. World's Poult. Sci. J., 59: 185-200.
CrossRefDirect Link - Zhao, K., G. Chen, X.M. Shi, T.T. Gao and W. Li et al., 2012. Preparation and efficacy of a live Newcastle disease virus vaccine encapsulated in chitosan nanoparticles. PLoS ONE, Vol. 7.
CrossRefDirect Link - Chen, K. and A. Cerutti, 2010. Vaccination strategies to promote mucosal antibody responses. Immunity, 33: 479-491.
CrossRefDirect Link - Kang, H., H. Wang, Q. Yu and Q. Yang, 2012. Effect of intranasal immunization with inactivated avian influenza virus on local and systemic immune responses in ducks. Poult. Sci., 91: 1074-1080.
CrossRefDirect Link - Savelkoul, H.F.J., V.A. Ferro, M.M. Strioga and V.E.J.C. Schijns, 2015. Choice and design of adjuvants for parenteral and mucosal vaccines. Vaccines, 3: 148-171.
CrossRefDirect Link - Pichichero, M.E., 2008. Improving vaccine delivery using novel adjuvant systems. Hum. Vaccines, 4: 262-270.
CrossRefDirect Link - Park, K.S., J. Lee, S.S. Ahn, Y.H. Byun and B.L. Seong et al., 2009. Mucosal immunity induced by adenovirus-based H5N1 HPAI vaccine confers protection against a lethal H5N2 avian influenza virus challenge. Virology, 395: 182-189.
CrossRefPubMedDirect Link - Jang, S.I., H.S. Lillehoj, S.H Lee, K.W. Lee and E.P. Lillehoj et al., 2011. Montanide™ IMS 1313 N VG PR nanoparticle adjuvant enhances antigen-specific immune responses to profilin following mucosal vaccination against Eimeria acervulina. Vet. Parasitol., 182: 163-170.
CrossRefDirect Link - Riffault, S., G. Meyer, M. Deplanche, C. Dubuquoy and G. Durand et al., 2010. A new subunit vaccine based on nucleoprotein nanoparticles confers partial clinical and virological protection in calves against bovine respiratory syncytial virus. Vaccine, 28: 3722-3734.
CrossRefDirect Link - King, D.J., 1991. Evaluation of different methods of inactivation of Newcastle disease virus and avian influenza virus in egg fluids and serum. Avian Dis., 35: 505-514.
CrossRefDirect Link - Suzuki, K., H. Okada, T. Itoh, T. Tada and M. Mase et al., 2009. Association of increased pathogenicity of Asian H5N1 highly pathogenic avian influenza viruses in chickens with highly efficient viral replication accompanied by early destruction of innate immune responses. J. Virol., 83: 7475-7486.
CrossRefDirect Link - Londt, B.Z., A. Nunez, J. Banks, H. Nili, L.K. Johnson and D.J. Alexander, 2008. Pathogenesis of highly pathogenic avian influenza A/turkey/Turkey/1/2005 H5N1 in Pekin ducks (Anas platyrhynchos) infected experimentally. Avian Pathol., 37: 619-627.
CrossRefDirect Link - Wise, M.G., D.L. Suarez, B.S. Seal, J.C. Pedersen and D.A. Senne et al., 2004. Development of a real-time reverse-transcription PCR for detection of Newcastle disease virus RNA in clinical samples. J. Clin. Microbiol., 42: 329-338.
CrossRefDirect Link - Sasaki, T., N. Isoda, K. Soda, R. Sakamoto and K. Saijo et al., 2009. Evaluation of the potency, optimal antigen level and lasting immunity of inactivated avian influenza vaccine prepared from H5N1 virus. Jpn. J. Vet. Res., 56: 189-198.
CrossRefPubMedDirect Link - Joo, H.M., Y. He, A. Sundararajan, L. Huan and M.Y. Sangster, 2010. Quantitative analysis of influenza virus-specific B cell memory generated by different routes of inactivated virus vaccination. Vaccine, 28: 2186-2194.
CrossRefDirect Link - Smialek, M., B. Tykalowski, T. Stenzel and A. Koncicki, 2011. Local immunity of the respiratory mucosal system in chickens and turkeys. Polish J. Vet. Sci., 14: 291-297.
CrossRefDirect Link - Van Ginkel, F.W., D.C. Tang, S.L. Gulley and H. Toro, 2009. Induction of mucosal immunity in the avian Harderian gland with a replication-deficient Ad5 vector expressing avian influenza H5 hemagglutinin. Dev. Comp. Immunol., 33: 28-34.
CrossRefDirect Link - Macpherson, A.J., K.D. McCoy, F.E. Johansen and P. Brandtzaeg, 2008. The immune geography of IgA induction and function. Mucosal Immunol., 1: 11-22.
CrossRefDirect Link - Lee, D.H., J.K. Park, J.H. Kwon, S.S. Yuk and T.O. Erdene-Ochir et al., 2013. Efficacy of single dose of a bivalent vaccine containing inactivated Newcastle disease virus and reassortant highly pathogenic avian influenza H5N1 virus against lethal HPAI and NDV infection in chickens. PLoS ONE, Vol. 8.
CrossRefDirect Link - Kryczek, I., S. Wei, L. Vatan, J. Escara-Wilke, W. Szeliga, E.T. Keller and W. Zou, 2007. Cutting edge: Opposite effects of IL-1 and IL-2 on the regulation of IL-17+ T cell pool IL-1 subverts IL-2-mediated suppression. J. Immunol., 179: 1423-1426.
CrossRefDirect Link - Rauw, F., Y. Gardin, V. Palya, S. Anbari and M. Gonze et al., 2010. The positive adjuvant effect of chitosan on antigen-specific cell-mediated immunity after chickens vaccination with live Newcastle disease vaccine. Vet. Immunol. Immunopathol., 134: 249-258.
CrossRefDirect Link - Cardenas-Garcia, S., R.P. Dunwoody, V. Marcano, D.G. Diel and R.J. Williams et al., 2016. Effects of chicken interferon Gamma on Newcastle disease virus vaccine immunogenicity. PLoS ONE, Vol. 11.
CrossRefDirect Link - Julkunen, I., K. Melen, M. Nyqvist, J. Pirhonen, T. Sareneva and S. Matikainen, 2000. Inflammatory responses in influenza A virus infection. Vaccine, 19: S32-S37.
CrossRefDirect Link - Osterlund, P., V. Veckman, J. Siren, K.M. Klucher, J. Hiscott, S. Matikainen and I. Julkunen, 2005. Gene expression and antiviral activity of alpha/beta interferons and interleukin-29 in virus-infected human myeloid dendritic cells. J. Virol., 79: 9608-9617.
CrossRefPubMedDirect Link