
A.A.J. Mofunanya
Department of Botany, University of Calabar, PMB 1115, Calabar, Nigeria
A.T. Owolabi
Department of Botany, University of Calabar, PMB 1115, Calabar, Nigeria
A. Nkang
Department of Botany, University of Calabar, PMB 1115, Calabar, Nigeria
International Journal of Plant Pathology
Year: 2014 | Volume: 5 | Issue: 2 | Page No.: 54-62







ABSTRACT
Fluted pumpkin (Telfairia occidentalis) is the foremost and most popular leafy vegetable in the diet of the inhabitants of Southeastern Nigeria, where its cultivation forms the economic mainstay of several families. Infection of the vegetable by Telfairia mosaic virus (TeMV) engendered severe mosaic, leaf distortion and malformation, rendering it unmarketable. In this study, the effects of TeMV on the chlorophyll content and photosynthetic capability of two ecotypes of T. occidentalis (herein referred to as ecotypes A and B) using nitrogen content as indicator were investigated. Chlorophyll contents were determined at 14, 30, 60, 90 and 120 days post inoculation (dpi) after extraction in 80% acetone, the supernatant filtered and values read spectrophotometrically. Nitrogen content was evaluated using the standard micro-Kjeldahl technique. Regarding the chlorophyll content, the results indicated that chlorophylls a, b and total chlorophyll were significantly reduced (p≤0.05) in both ecotypes A and B when compared to the controls at all growth stages. The results also showed a greater reduction of chl a than chl b in both ecotypes. The Nitrogen (N) content in both ecotypes were significantly reduced (p≤0.05) at all growth stages when compared to healthy controls. There were greater reductions in both chlorophyll and N contents in ecotype A than in B. The implication of the result is the inevitable loss in photosynthetic capabilities of both ecotypes with resultant poor vegetative growth and less economic returns.
PDF Abstract XML References Citation
Received: August 15, 2013;
Accepted: March 21, 2014;
Published: August 18, 2014
How to cite this article
A.A.J. Mofunanya, A.T. Owolabi and A. Nkang, 2014. Effects of Telfairia mosaic virus (TeMV) on the Chlorophyll Content
and Photosynthetic Capabilities of Two Ecotypes of Telfairia occidentalis
Hook. f. (Fluted Pumpkin). International Journal of Plant Pathology, 5: 54-62.
DOI: 10.3923/ijpp.2014.54.62
URL: https://scialert.net/abstract/?doi=ijpp.2014.54.62
DOI: 10.3923/ijpp.2014.54.62
URL: https://scialert.net/abstract/?doi=ijpp.2014.54.62
INTRODUCTION
Fluted pumpkin, Telfairia occidentalis (Hook), a member of the Cucurbitaceae family, grows fast in the warm humid tropics and produces leaves for most of the rainy season of 6-10 months (Akoroda, 1990). Although, mostly grown as annual, the plant is a leafy perennial climber which attaches itself to other plants or objects by means of tendrils (Tindall, 1983) and can grow to about 10 m in length (Herklots, 1972). Fluted pumpkin is said to be widely cultivated for its edible succulent shoots and leaves (Akoroda, 1990). The leaves which are compound, palmately lobed and usually 3-5 foliolate are used in the preparation of the much relished edikaikong soup in the southern eastern states of Nigeria.
According to Nono-Womdim (2003), viruses constitute a major limiting factor to vegetable production in most African countries. The production of T. occidentalis is reported to be limited by a number of diseases, some of which are of viral aetiology (Anno-Nyako, 1988). Symptoms such as mosaic, severe leaf distortion and malformation and poor growth are characteristic of TeMV infection of T. occidentalis. Virus infection has been reported to induce, among others, changes in host plant metabolic processes including photosynthesis (Havelda and Maule, 2000). Loss of photosynthetic activity is evident in the appearance of macroscopic symptoms such as yellow/green mosaic patterns or total loss of chlorophyll (chlorosis). Mosaic diseases are known to be commonly associated with decreased photosynthesis, as a result of chlorophyll loss and consequently less organic carbon in the infected leaves (Matthews, 1991).
The literature is replete with reports of virus infection on the amount of pigments in several plant-virus combinations. In sorghum infected with Sorghum red stripe virus (SRSV), Pawar et al. (1990) reported a reduction in the chlorophyll content at all stages of growth. El-Fahaam et al. (1990) also reported significant decrease in the chlorophyll content of lettuce infected with Lettuce mosaic virus (LMV). Muqit et al. (2007) observed a decrease in chlorophyll content in ash gourd due to infection by Bottle gourd mosaic virus (BgMV), Watermelon mosaic virus-2 (WMV-2) and Papaya ringspot virus (PRSV). Similarly, reductions in chlorophyll contents have also been reported in several other host plant-virus combinations (Mali et al., 2000; Milavec et al., 2001; Funayama-Noguchi and Terashima, 2006; Singh and Shukla, 2009; Arora et al., 2009; Sinha and Srivastava, 2010).
Photosynthetic rate has been demonstrated to be related to leaf nitrogen content (Ripullone et al., 2003; Bojovic and Markovic, 2009). As noted by Evans (1989), the proteins of the Calvin cycle and thylakoids represent the majority of leaf nitrogen. The thylakoids, otherwise known as the grana, are located inside the chloroplast and have been demonstrated to constitute the site of the light-dependent reactions of photosynthesis. They are believed to consists of at least 335 proteins (Van Wijk, 2004; Friso et al., 2004; Kleffmann et al., 2006), 42% of which are involved in photosynthesis (Friso et al., 2004).
Previous study by Mofunanya et al. (2007) has documented the effect of TeMV on vitamins and amino acid profile of two ecotypes of T. occidentalis. The effect of the virus on the proximate, mineral and anti-nutritive contents of the vegetable has also been reported (Mofunanya et al., 2008). The present study examines the effect of TeMV on the chlorophyll content and photosynthetic capability of two ecotypes of T. occidentalis using nitrogen content as an indicator.
MATERIALS AND METHODS
Virus source and propagation and inoculum preparation: The virus used in this study, was supplied by (Federal Biological Centre for Agriculture and Forestry, Braunschweig, Germany). The virus which had been stored under liquid nitrogen, was reactivated by triturating the infected tissues in disodium phosphate (Na2HPO4) buffer 0.03 M pH 8.0. The inoculum was rubbed onto carborundum (800-mesh) dusted Nicotiana benthamiana. Symptoms developed two weeks later. The virus was mechanically transferred to T. occidentalis and subsequently maintained in the plant in the greenhouse at 25±3°C.
Sources of seed: Two ecotypes of T. occidentalis were used in this study. One batch was sourced from Ikom, a town in Cross River State, located on latitude 5°57'N and longitude 8°44'E herein denoted as ecotype A. The second batch of seed lot was obtained from Eket, a coastal town in Akwa Ibom State, Nigeria located on latitude 4°37’N and longitudes 7°55'E, herein referred to as ecotype B. Both ecotypes could be distinguished by the general morphology of the plant and seed characteristics. The seeds of ecotype A are generally smaller, measuring about 3.0 cm in length, fruit pod of 25 16 cm-1 (length/width) and with large green leaves. The seeds of ecotype B on the average, measures about 3.5 cm (length/width), fruit pod size of 30 17 cm-1 (length/width) with larger and greener leaves compared to ecotype A.
Experimental design: Thirty seedlings of each ecotype were established in plastic pots 16 cm in diameter. The seedlings were arranged in a randomized block design in three tows of ten. Within each block fifteen plants were randomly sap-inoculated two weeks after planting and the remaining fifteen were inoculated with buffer only to serve as controls. The set up was monitored for symptom expression.
Effect of TeMV on chlorophyll content: For chlorophyll content determination, infected and healthy leaves of the two ecotypes were obtained at 14, 30, 60, 90 and 120 days post inoculation. A two gram leaf sample from both infected and healthy leaves of both ecotypes was homogenized separately in mortar using the pestle in 10 mL of 80% acetone. The homogenates were dispensed into tubes and centrifuged at 4500 rpm for 3 min. The supernatants were decanted and used for chlorophyll determination. Blank set a zero with 3 mL of acetone was prepared. To this was added 0.1 mL of the supernatant and absorbance readings were taken at 663 nm wavelength for chlorophyll a (chl a) and 643 nm wavelength for chl b using spectrophotometer (model 722S, England). Readings were obtained from three samples from each preparation and the chlorophyll contents of the samples were determined according to the formula for the estimation of Strickland and Parsons (1972):
Chl a = (11.6 A663-1.3 A643) VX-1
Chl a = (19.6 A643-4.7 A663) VX-1
Chl a+b = (mg g-1 FW)
where, A663 and A643 are the absorbances at 663 and 643 nm, respectively.
| • | A | : | Absorbance |
| • | V | : | Volume (mL) of 80% acetone |
| • | X-1 | : | Fresh weight of samples used |
| • | Mg | : | Milligram |
| • | g | : | Gram |
| • | FW | : | Fresh weight |
Effect of TeMV on photosynthetic rate using nitrogen as indicator: Leaf sample were obtained at 14, 30, 60, 90 and 120 days post inoculation from infected and healthy plant of both ecotypes. Each sample weighing 20 g was oven-dried and pulverized and analyzed for nitrogen content using standard micro-Kjeldahl technique as described by Miller and Houghton (1945).
Statistical analysis: Data obtained presented were means of three determinations and were analyzed using independent t-test and differences between means were determined at 5% probability.
RESULTS
The results of TeMV inoculation on chlorophylls a, b and total chlorophyll (a+b) contents of T. occidentalis are presented in Table 1, 2.
| Table 1: | Effect of Telfairia mosaic virus on chlorophyll content of Telfairia occidentalis ecotype A |
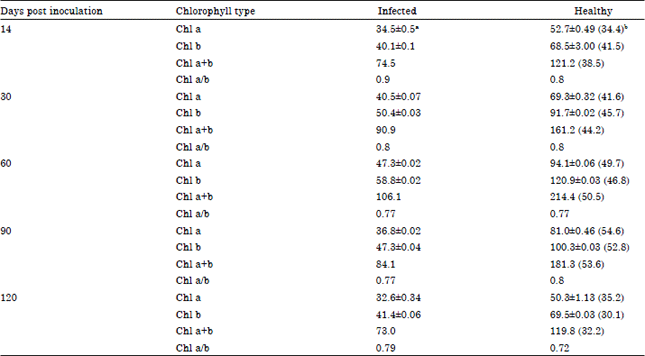 | |
| aValues are means of three readings, bFigures in parentheses are percentage reductions | |
| Table 2: | Effect of Telfairia mosaic virus on chlorophyll content of Telfairia occidentalis ecotype B |
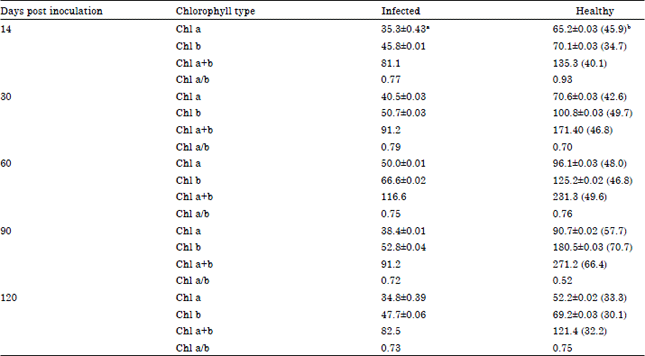 | |
| aValues are means of three readings, bFigures in parentheses are percentage reductions | |
The results show that chlorophylls a, b and total chlorophyll were significantly reduced in both ecotypes A and B when compared to the controls at all growth stages.
The effect of TeMV on the chlorophyll contents showed a gradual loss up to 90 dpi and the effect become less severe as the plant aged. For ecotype A, reduction in chl a content by TeMV was least at 14 days post inoculation (dpi) with a mean value of 34.5±0.5 mg g-1 FW compared to 52.7±0.49 mg g-1 FW for the control, representing a percentage reduction of 34.4%. The highest reduction was recorded at 90 dpi with a mean value of 36.8±0.02 mg g-1 FW compared to the mean value of 81.0±0.46 mg g-1 FW obtained for the corresponding control, representing a percentage reduction of 54.6%. Chl b content was reduced by as much as 46.8 and 52.8% at 60 and 90 dpi, respectively and between 41.5 (14 dpi) and 45.7% (30 dpi) at the other growth stages with the least at 120 dpi (Table 1).
For ecotype B, the results showed greater losses in chls a and b contents as the disease progressed up to 90 dpi. The least reduction for chl a was recorded at 120 dpi with a mean value of 34.8±0.39 mg g-1 FW compared to the healthy with a mean value of 52.2±0.02 mg g-1 FW, representing a percentage reduction of 33.3% while the highest loss was observed at 90 dpi when the mean value for the infected sample was 38.4±0.01 and the control was 90.7±0.02 mg g-1 FW representing 57.7% reduction.
For chl b, the least reduction of 30.1% was recorded at 120 dpi and the highest amounting to 70.7% at 90 dpi when compared to the mean values obtained for the corresponding controls (Table 2). The results also showed significantly losses in total chlorophyll (chl a and b) of both ecotypes as the disease progressed. The greatest losses occurred between 60 and 90 dpi and the least at 120 dpi. Reductions of between 32.2 and 53.6% in the total chlorophyll content of infected leaf samples of ecotype A were obtained while for the ecotype B the losses stood at between 32.2 and 66.4%.
TeMV engendered significant reductions (p≤0.05) in the nitrogen contents in the leaves of both ecotypes A and B at all the stages of growth evaluated when compared to control (Fig. 1). Greater losses were observed as the disease progressed. For ecoype A, percentage reduction in the N content ranged from 28.8 at 14-55.1% at 120 dpi.
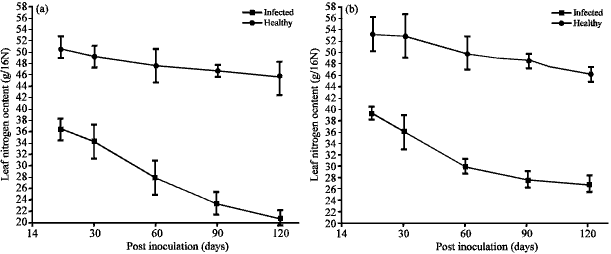 | |
| Fig. 1(a-b): | Photsynthetic capability of Telfairia occidentalis inoculated with Telfairia mosaic virus using nitrogen content as indicztor (a) Ecotype A and (b) Ecotype B |
The mean N values for both growth stages were 35.8 g and 20.5 g/16N, respectively while the corresponding mean values for the healthy samples were 50.3 g and 45.7 g/16N. For ecotype B, percentage reductions ranged from 26.4% at 14 dpi which had a mean N content of 39.9 g/16N, to 42.3% at 120 dpi with a mean N value of 27.5 g/16N. The corresponding values for the healthy controls for both growth stages were 54.2 g and 47.6 g/16N, respectively.
DISCUSSION
The effects of TeMV on the chlorophyll content and the photosynthetic capability, using nitrogen content in the infected leaf tissues as indicator, in two ecotypes of T. occidentalis over a period of 120 days were studied. Infection of the leafy vegetable by the virus led to significant reductions in both chl a, chl b and total chlorophyll (Chl a+b) contents in symptomatic leaves of both ecotypes at all the developmental stages when compared to the healthy control. The results also showed a greater reduction of chl a than chl b in both ecotypes. Similar to the results obtained in this study, reduction in the chlorophyll contents engendered by viruses have been reported in several plant species. Sugarcane infected by Sugarcane yellow leaf virus (SCYLV) (Goncalves et al., 2005) and infection of Eupatorium makinoi by Eupatorium yellow vein virus (EYMV) (Funayama-Noguchi and Terashima, 2006) have been reported to cause significant reduction in the chlorophyll content. Also, ash gourd (Benincasa hispida) singly inoculated with BgMV, Watermelon virus-2 (WMV-2) and PRSV (Muqit et al., 2007), mothbean (Vigna aconitifolia) (Arora et al., 2009) and mungbean (Phaseolus aureus) inoculated with Mungbean yellow mosaic virus (MYMV) (Sinha and Srivastava, 2010) have been reported to cause significant reduction in chlorophyll content of the infected leaf tissues.
Loss of chlorophyll in infected leaves has been attributed to either impaired chlorophyll synthesis (Funayama-Noguchi and Terashima, 2006), structural alterations in the photosynthetic apparatus (Goncalves et al., 2005) or reduced activities of some enzymes associated with electron transport and photosynthesis (Zhou et al., 2004; Guo et al., 2005). Generally, potyviruses have not been reported to affect the structural integrity of chloroplasts of infected leaf tissues. Rather, they have been shown to cause chloroplast aggregation and consequently a reduction in their number (Jin et al., 2007; Huang et al., 2010; Wei et al., 2010, 2013) and inhibition of enzymes associated with carbon assimilation (Zhou et al., 2004; Guo et al., 2005). The significant reduction in the chlorophyll contents of both ecotypes of T. occidentalis by TeMV, a potyvirus, could be said to be due to a reduction in the number of chloroplasts in the infected leaf tissues.
Nitrogen is one of the basic components of nucleic acids, amino acids, proteins, enzymes and chlorophyll. At all the developmental stages of growth for which data were obtained, TeMV significantly reduced the amount of nitrogen in the two ecotypes of T. occidentalis. This result implies a significant loss in the photosynthetic capability of the infected ecotypes. Studies have shown positive correlation between leaf nitrogen content and the photosynthetic capability of leaves (Reich et al., 1995; Ripullone et al., 2003; Bojovic and Markovic, 2009). According to Evans (1989), the proteins of the Calvin cycle and thylakoids represent the majority of leaf nitrogen and to a first approximation thylakoid nitrogen has been shown to be proportional to the chlorophyll content. Lee et al. (2011) have also shown a correlation between leaf nitrogen and chlorophyll content. According to their report, plants with higher N content not only had higher chlorophyll content but also had thicker, flatter and more turgid leaves which enhanced visible light absorption while stimulating near-infrared reflection. By inference, the significant reduction in the nitrogen content by TeMV suggests a significant reduction in the photosynthetic capability of T. occidentalis and poor plant growth found always associated with TeMV.
Since TeMV resulted in significant losses in chlorophyll and leaf nitrogen content and by extension photosynthetic capability of both ecotypes, continued cultivation of these ecotypes where TeMV is endemic may result in poor vegetative growth and concomitant economic losses. The search for resistant varieties and their adoption for cultivation are suggested.
ACKNOWLEDGMENT
The authors are grateful to Mr. S. M. Ekpo of the Department of Biochemistry, University of Calabar, Nigeria for his assistance in executing the micro-Kjeldahl technique.
REFERENCES
- Akoroda, M.O., 1990. Seed production and breeding potential of the fluted pumpkin, Telfairia occidentalis. Euphytica, 49: 25-32.
CrossRefDirect Link - Anno-Nyako, F.O., 1988. Seed transmission of telfairia mosaic virus in fluted pumpkin (Telfairia occidentalis Hook f.) in Nigeria. J. Phytopathol., 121: 85-87.
CrossRefDirect Link - Arora, R., U.N. Joshi, P.P. Gupta and J.V. Singh, 2009. Effect of Yellow mosaic virus on pathogenesis related enzymes and chlorophyll content in mothbean (Vigna aconitifolia). Acta Phytopathologica Entomologica Hungarica, 44: 49-60.
CrossRefDirect Link - Bojovic, B. and A. Markovic, 2009. Correlation between nitrogen and chlorophyll contentin wheat (Triticum aestivum L.). Kragujevac J. Sci., 31: 69-74.
Direct Link - El-Fahaam, Y.M., G.I. Fegla, E.E. Wagih and H.A. El-Karyoni, 1990. Biochemical changes in lettuce plants infected with lettuce mosaic virus. J. King Saud Univ. Agric. Sci., 2: 271-277.
Direct Link - Evans, J.R., 1989. Photosynthesis and nitrogen relationships in leaves of C₃ plants. Oecologia, 78: 9-19.
CrossRefDirect Link - Friso, G., L. Giacomelli, A.J. Ytterberg, J.B. Peltier, A. Rudella, Q. Sun and K.J. Van Wijk, 2004. In-depth analysis of the thylakoid membrane proteome of Arabidopsis thaliana Chloroplasts: New proteins, new functions and a plastid proteome database. Plant Cell, 16: 478-499.
PubMedDirect Link - Funayama-Noguchi, S. and I. Terashima, 2006. Effects of Eupatorium yellow vein virus infection on photosynthetic rate, chlorophyll content and chloroplast structure in leaves of Eupatorium makinoi during leaf development. Functional Plant Biol., 33: 165-175.
CrossRefDirect Link - Goncalves, M.C., J. Vega, J.G. Oliveira and M.M.A. Gomes, 2005. Sugarcane yellow leaf virus infection leads to alterations in photosynthetic efficiency and carbohydrate accumulation in sugarcane leaves. Fitopatol. Bras., 30: 10-16.
CrossRefDirect Link - Guo, Y.P., D.P. Guo, Y. Peng and J.S. Chen, 2005. Photosynthetic responses of radish (Raphanus sativus var. longipinnatus) plants to infection by turnip mosaic virus. Photosynthetica, 43: 457-462.
CrossRefDirect Link - Huang, T.S., T. Wei, J.F. Laliberte and A. Wang, 2010. A host RNA helicase-like protein, AtRH8, interacts with the potyviral genome-linked protein, VPg, associates with the virus accumulation complex and is essential for infection. Physiology, 152: 255-266.
CrossRef - Jin, Y., D. Ma, J. Dong, D. Li, C. Deng, J. Jin and T. Wang, 2007. The HC-Pro protein of Potato virus Y interacts with NtMinD of tobacco. Mol. Plant Microbe Interact., 20: 1505-1511.
CrossRef - Kleffmann, T., M. Hirsch-Hoffmann, W. Gruissem and S. Baginsky, 2006. Plprot: A comprehensive proteome database for different plastid types. Plant Cell Physiol., 47: 432-436.
CrossRef - Lee, Y.J., C.M. Yang, K.W. Chang and Y. Shen, 2011. Effects of nitrogen status on leaf anatomy, chlorophyll content and canopy reflectance of paddy rice. Botanical Stud., 52: 295-303.
Direct Link - Mali, P.C., U. Burman and S. Lodha, 2000. Effect of planting dates and development of yellow mosaic viruson biochemical constituents of moth bean genotypes. Indian J. Phytopathol., 53: 379-383.
Direct Link - Milavec, M., M. Ravnikar and M. Kovac, 2001. Peroxidases and photosynthetic pigments in susceptible potato infected with potato virus YNTN. Plant Physiol. Biochem., 39: 891-898.
CrossRefDirect Link - Miller, L. and J.A. Houghton, 1945. The micro-kjeldahl determination of the nitreogen content of amino acids and proteins. J. Biol. Chem., 159: 373-383.
Direct Link - Muqit, A., A.M. Akanda and K.A. Kader, 2007. Biochemical alteration of cellular components of ash gourd due to infection of three different viruses. Int. J. Sustainable Crop Prod., 2: 40-42.
Direct Link - Reich, P.B., M.B. Walters, B.D. Kloeppel and D.S. Ellsworth, 1995. Different photosynthesis-nitrogen relations in deciduous hardwood and evergreen coniferous tree species. Oecologia, 104: 24-30.
CrossRef - Ripullone, F., G. Grassi, M. Lauteri and M. Borghetti, 2003. Photosynthesis-nitrogen relationships: Interpretation of different patterns between Pseudotsuga menziesii and Populus × euroamericana in a mini-stand experiment. Tree Physiol., 23: 137-144.
CrossRef - Singh, V. and K. Shukla, 2009. Effect of PRSV infection on pigment content and assimilation of carbohydrate in Carica papaya L. Ann. Plant Prot. Sci., 17: 152-156.
Direct Link - Sinha, A. and M. Srivastava, 2010. Biochemical changes in mungbean plants infected by Mungbean yellow mosaic virus. Int. J. Virol., 6: 150-157.
CrossRefDirect Link - Wei, T., T.S. Huang, J. McNeil, J.F. Laliberte, J. Hong, R.S. Nelson and A. Wang, 2010. Sequential recruitment of the endoplasmic reticulum and chloroplasts for plant Potyvirus replication. J. Virol., 84: 799-809.
CrossRefDirect Link - Wei, T., C. Zhang, X. Hou, H. Sanfacon and A. Wang, 2013. The SNARE protein Syp71 is essential for turnip mosaic virus infection by mediating fusion of virus-induced vesicles with chloroplasts. PLoS Pathogens, Vol. 9.
CrossRef - Zhou, Y.H., Y.H. Peng, J.L. Lei, L.Y. Zou, J.H. Zheng and J.Q. Yu, 2004. Effects of potato virus YNTN infection on gas exchange and photosystem 2 function in leaves of Solanum tuberosum L. Photosynthetica, 42: 417-423.
CrossRef - Havelda, Z. and A.J. Maule, 2000. Complex spatial responses to cucumber mosaic virus infection in susceptible Cucurbitapepo cotyledons. Plant Cell, 12: 1975-1985.
Direct Link