
Saurabh Singh
Department of Mycology and Plant Pathology, Institute of Agricultural Sciences, Banaras Hindu University, 221 005, Varanasi, India
Asha Sinha
Department of Mycology and Plant Pathology, Institute of Agricultural Sciences, Banaras Hindu University, 221 005, Varanasi, India
Richa Raaj
Department of Mycology and Plant Pathology, Institute of Agricultural Sciences, Banaras Hindu University, 221 005, Varanasi, India
International Journal of Plant Pathology
Year: 2014 | Volume: 5 | Issue: 1 | Page No.: 21-27







ABSTRACT
Mungbean seeds genotype HUM-4 and HUM-12 were infested with four dominant fungi viz., Alternaria alternata, Aspergillus niger and Penicillium citrum and Fusarium moniliformae as well as fresh seeds were used to test germination ability of mungbean seeds by using different methods viz., Blotter methods, Multi-pot trey methods, Earthen pot method and Rolled paper towel method. Result showed that all infested seeds were found to be significantly reduced the germination of seeds. In Blotter condition, maximum germination was recorded (93.30%) in genotype HUM-12 followed by (90.35%) in Genotype HUM-4. In paper towel methods, freshly harvested mungbean seeds showed maximum seed germination (90.00%) in genotype HUM-12 followed by (88.67%) in Genotype HUM-4. In multipot trey method, freshly harvested mungbean seed showed maximum seed germination (84.12%) in genotype HUM-12 followed by (83.33%) in Genotype HUM-4. In earthen pot methods freshly harvested mungbean seed showed maximum seed germination (87.30%) in genotype HUM-12 followed by (85.23%) in Genotype HUM-4. Aspergillus niger infested seeds showed lowest germination followed by Penicillium rubrum infested seeds.
PDF Abstract XML References Citation
Received: October 24, 2013;
Accepted: January 06, 2014;
Published: April 11, 2014
How to cite this article
Saurabh Singh, Asha Sinha and Richa Raaj, 2014. Determination of Seed Germination of Fresh and Infested Stored Mungbean
Vigna radiata (L.) Wilczek Seeds by using Different Method. International Journal of Plant Pathology, 5: 21-27.
DOI: 10.3923/ijpp.2014.21.27
URL: https://scialert.net/abstract/?doi=ijpp.2014.21.27
DOI: 10.3923/ijpp.2014.21.27
URL: https://scialert.net/abstract/?doi=ijpp.2014.21.27
INTRODUCTION
Pulses are rich source of vegetative protein and play and important role in nutritional security of majority of vegetarian population in India. The country is the largest producer and consumer of pulses occupying 33% of the world’s area and 22% of the production (FAO, 2008). Pulse production in the country has fluctuated widely between 13 and 15 million tonnes (mt) with no significant growth trend between 1991 and 2010. The latest estimate indicates that the present production of pulses has reached 14.7 million tons (mt) with productivity of 637 kg ha-1 although the projected pulse requirement by the year 2030 (32 mt) is estimated to be more than double the current production level (Anonymous, 2011). Seed health testing for the presence of seed borne pathogens is an important step in the management of crop diseases. Seed-borne pathogens have been involved in seed rots during germination and seedling mortality leading to poor crop stand (Khalid et al., 2001); reduction in plant growth and productivity of crops (Williams and McDonald, 1983; Kubiak and Korbas, 1999; Dawson and Bateman, 2001). The seed-borne pathogens associated with seeds externally or internally may cause seed abortion, seed rot, seed necrosis, reduction or elimination of germination capacity, as well as seedling damage resulting in development of disease at later stages of plant growth by systemic or local infection (Bateman and Kwasna, 1999; Ijaz et al., 2001; Khanzada et al., 2002). The present study was conducted to identify and investigate the association of seed borne mycoflora with two genotype of mungbean seeds and their effects on seed germination.
MATERIALS AND METHODS
Determination of seed germination of fresh and infested seeds: Mungbean seeds genotype HUM-4 and HUM-12 were infested with four dominant fungi viz., Alternaria alternata, Aspergillus niger and Penicillium citrum and Fusarium moniliformae as well as fresh seeds were used to test germination ability of mungbean seeds. Seed germination was determined by using three methods which are as follows:
| • | Blotter method: Three pieces of blotting paper were placed in fold in each Petri dish of 9 cm diameter and incubated at 25±2°C. Sixteen seeds were placed in each Petri dish and three replicates were prepared. The percent germination of mungbean seed were calculated after one week (De Tempe, 1953) |
| • | Multi-pot tray method: Plastic multi-pot trays were used to test the germination of mungbean seeds in glass house. The small pots were filled with the pre-sterilized (95°C for 30 min in autoclave) uniform soil mixture containing 4 parts peat with essential amount of fertilizer and seeds were sown in them. The trays were covered with a polyethylene envelope so as to reduce water loss through evaporation (Khare, 1996) |
| • | Earthen pot method: In this method earthen pots were used to test for determination of percent germination of mungbean seeds in glass house. The earthen pot filled with the pre-sterilized (95°C for 30 min in autoclave) uniform soil mixture containing 4 parts peat with essential amount of fertilizer and seeds sown in them the pots were covered with a polypropylene envelope so as to reduce water loss through evaporation. The germination was counted 5-7 days after sowing |
| • | Rolled paper towel method: Sterilized water is used for moisten of the towel for 6 h for removed water soluble toxin from the paper towels and seeds placed over the towel paper. The towel is then wrapped in a sheet of polyethylene, at ten seeds per row to check surface. Hundred seeds of each treatment was replicated thrice and tested by employing rolled towel method and incubated at 25± 2°C under 12 h light and 12 h darkness as described earlier. After seven days of incubation, percent germination was recorded. The first count for germinated seeds was taken 5 days and the second 7 days after plating, respectively |
RESULTS
Determination of seed germination of fresh and stored mungbean seed by using rolled paper towel method: Under accelerated ageing environment seed germination percentage progressively decreased with the duration of ageing. Effect of selected dominant seed borne mycoflora on germination of mungbean seeds genotype on rolled paper towel method is presented in Table 1. The effect significantly differed from genotype to genotype. Result revealed that seed germination of mungbean seed was decreased due to infestation of selected dominant seed mycoflora viz., Alternaria alternata, Aspergillus niger, Fusarium moniliformae and Penicillium rubrum.
| Table 1: | Determination of germination of mungbean seed by using rolled paper towel method |
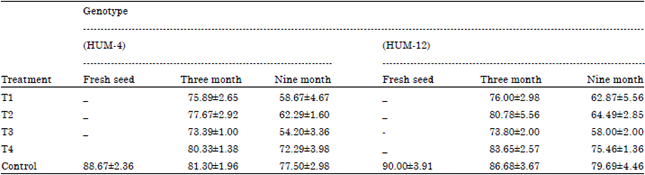 | |
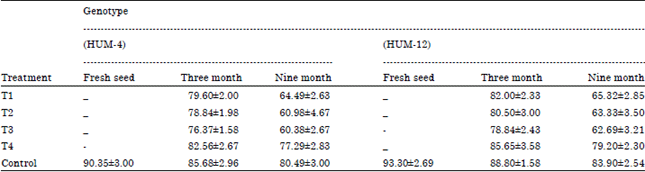
| Table 2: | Determination of germination of mungbean seed by using rolled paper towel method |
 | |
Percent germination was declined from 180-360 days of storage. Freshly harvested mungbean seed showed maximum seed germination (90.00%) in genotype HUM-12 followed by (88.67%) in Genotype HUM-4. After 180 days period of storage maximum seed germination was observed in control (86.68%) in Genotype HUM-12 followed by (81.30%) in Genotype HUM-4, Fusarium moniliformae infested seed showed (83.65%) in genotype HUM-12 and (80.33%) in Genotype HUM-4. Minimum germination was recorded in Aspergillus niger infested seeds (73.39%) in genotype HUM-4 followed by (73.80%) in Genotype HUM-12. After 360 days period of storage maximum seed germination was observed in control (79.69%) in Genotype HUM-12 followed by (77.60%) in Genotype HUM-4 and Fusarium moniliformae infested seed (75.46%) in genotype HUM-12 followed by (72.29%) in Genotype HUM-4. Lowest germination was showed in Aspergillus niger infested seeds (54.20%) in genotype HUM-4 followed by (58.00%) in Genotype HUM-12 The results showed that among all tested fungi Aspergillus niger infected seed reduced the maximum inhibition of germination followed by Penicillium rubrum.
Determination of germination of fresh and stored mungbean seed by using earthen pot method: Effect of the fungi on germination of mungbean seeds of two genotype on earthen pot method is presented in the (Table 2). The effect significantly differed from genotype to genotype. Result revealed that seed germination of mungbean seed was decreased due to infestation of selected dominant seed mycoflora viz., Alternaria alternata, Aspergillus niger, Fusarium moniliformae and Penicillium rubrum. Freshly harvested mungbean seed showed maximum seed germination (87.30%) in genotype HUM-12 followed by (85.23%) in Genotype HUM-4.
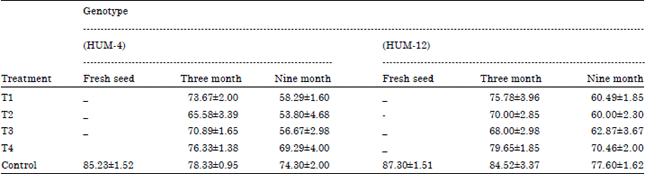
| Table 3: | Determination of germination in mungbean seed by using blotter paper methods |
 | |
After 180 days period of storage maximum seed germination was observed in control (84.52%) in Genotype HUM-12 followed by (78.33%) in Genotype HUM-4, Fusarium moniliformae infested seed showed (79.65%) in Genotype HUM-12 followed by (76.33%) in Genotype HUM-4 and minimum germination was recorded in Penicillium rubrum infested seeds (65.58%) in Genotype HUM-4 followed by (70.00%) in Genotype HUM-12. After 360 days period of storage maximum seed germination was observed in control (77.60%) in Genotype HUM-12 followed by (74.30%) in Genotype HUM-12, Fusarium moniliformae infested seed showed maximum germination (70.46%) in genotype HUM-12 followed by (69.29%) in Genotype HUM-12. Lowest germination was showed in Penicillium rubrum infested seeds (53.80%) in genotype HUM-4 followed by (60.00%) in Genotype HUM-12. The results showed that among all tested fungi Penicillium rubrum infected seed showed maximum inhibition of germination followed by Aspergillus niger.
Determination of germination of fresh and stored in mungbean seed by using blotter methods: Effect of the predominant seed borne fungi on germination of mungbean seeds of two genotype on earthen pot method is presented in Table 3. The effect significantly differed from genotype to genotype. Result revealed that seed germination of mungbean seed was decreased due to infestation of selected dominant seed mycoflora viz., Alternaria alternata, Aspergillus niger, Fusarium moniliformae and Penicillium rubrum. Freshly harvested mungbean seed showed maximum seed germination (93.30%) in genotype HUM-12 followed by (90.35%) in Genotype HUM-4. After 180 days period of storage maximum seed germination was observed in control (88.80%) in Genotype HUM-12 followed by (85.68%) in Genotype HUM-4, Fusarium moniliformae infested seed showed (85.65%) in genotype HUM-12 followed by (82.65%) in Genotype HUM-4 and minimum germination was recorded in Aspergillus niger infested seeds (76.37%) in Genotype HUM-4 followed by (78.84%) in Genotype HUM-12. After 360 days period of storage maximum seed germination was observed in control (83.90%) in Genotype HUM-12 followed by (80.49%) in Genotype HUM-4, Fusarium moniliformae infested seed showed maximum germination (79.20%) in genotype HUM-12 followed by (77.29%) in Genotype HUM-12. Lowest germination was showed in Aspergillus niger infested seeds (60.38%) in genotype HUM-4 followed by (62.69%) in Genotype HUM-12. The results showed that among all tested fungi Aspergillus niger infected seed reduced the maximum inhibition of germination followed by Penicillium rubrum.
Determination of germination fresh and stored in mungbean seed by using multipot trey method: Effect of the fungi on germination of mungbean seeds of two genotype on multipot trey method is presented in Table 4. Data presented in revealed that seed germination of Mungbean seed was decreased due to infestation of selected dominant seed mycoflora viz., Alternaria alternata, Aspergillus niger, Fusarium moniliformae and Penicillium rubrum.
| Table 4: | Determination of germination of mungbean seed by using multipot trey method |
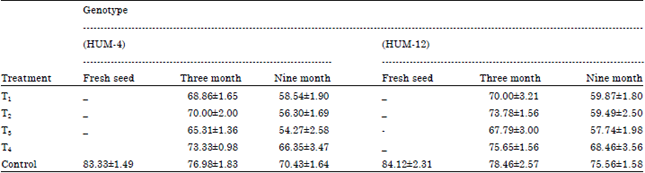 | |
Freshly harvested mungbean seed showed maximum seed germination (84.12%) in Genotype HUM-12 followed by (83.33%) in Genotype HUM-4. After 180 days period of storage maximum seed germination was observed in control (78.46%) in Genotype HUM-12 followed by (76.98%) in Genotype HUM-4, Fusarium moniliformae infested seed showed (75.65%) in Genotype HUM-12 followed by (73.33%) in Genotype HUM-4 and minimum germination was recorded in Aspergillus niger infested seeds (65.31%) in Genotype HUM-4 followed by (67.79%) in Genotype HUM-12. After 360 days period of storage maximum seed germination was observed in control (75.56%) in Genotype HUM-12 followed by (70.43%) in Genotype HUM-4, Fusarium moniliformae infested seed showed maximum germination (68.46%) in genotype HUM-12 followed by (66.35%) in Genotype HUM-4. Lowest germination was showed in Aspergillus niger infested seeds (54.27%) in genotype HUM-4 followed by (57.74%) in Genotype HUM-12. The results showed that among all tested fungi Aspergillus niger infected seed reduced the maximum inhibition of germination followed by Penicillium rubrum.
DISCUSSION
Result revealed that increasing in fungal incidence on seed reduces the viability and discoloration of seed caused germination failure. Among all tested fungi Aspergillus niger showed maximum inhibition of germination which is most harmful storage fungi and recorded to be one of the dominant genera on mungbean. Nik and Mamat (1982) also reported that when mungbean seed inoculated with Aspergillus flavus, A. niger, A. fumigatus and A. parasiticus showed a decline in germination of mungbean seed after 12th week of storage at 95% RH. Similar finding of result were reported by BARI (1981) in mungbean seed. The adverse effect of Aspergillus on the germination of cereals has been reported in the recent years (Kanujia and Singh, 1975). Aspergilllus sp. is most dominant fungus of Rice and also known as surface contaminate. Its produce aflatoxin during storage period that reduce the germination of seed (Imolehin, 1987). There are several reports that infections by storage fungi are a primary cause of loss of germination (Barton, 1961; Harrington, 1963; Mills and Frydman, 1980; Reddy et al., 2001; Tariq et al., 2005). Utobo et al. (2011) reported that Fusarium invade the seed coat, endosperm and embryo resulting in failure of germination. Fusariums are known to produce phytotoxins which probably interfere with germination (Ellis, 1971; Neergaard, 1977; Suryanarayana, 1978; Kanapathipillai and Hashim, 1982). Misra et al. (1975) reported that the fungi viz., Fusarium moniliforme, Rhizopus nigricans and Penicillium oxalicum caused marked reduction in germination of seed. Effect of A. alternata on seed germination was reported by McGee et al. (1980). Fusarium moniliforme and A. alternata reduced germination and induced seedling blight (Konde et al., 1980; Karim, 2005). The association of fungal pathogens with millet seeds demonstrates that the seeds are a major source of transmission of pathogens, which might have adverse effects at seedling and adult stage of plants. Finding has demonstrated that associated fungal pathogens reduce the germination ability of seeds, which causes poor crop stand, a major constraint of low harvested crop yield.
REFERENCES
- Bateman, G.L. and H. Kwasna, 1999. Effects of number of winter wheat crops grown successively on fungal communities on wheat roots. Applied Soil Ecol., 13: 271-282.
CrossRefDirect Link - Dawson, W.A.J.M. and G.L. Bateman, 2001. Fungal communities on roots of wheat and barley and effects of seed treatments containing fluquinconazole applied to control take-all. Plant Pathol., 50: 75-82.
CrossRefDirect Link - Khanzada, K.A., M.A. Rajput, G.S. Shah, A.M. Lodhi and F. Mehboob, 2002. Effect of seed dressing fungicides for the control of seedborne mycoflora of wheat. Asian J. Plant Sci., 1: 441-444.
CrossRefDirect Link - Konde, B.K., B.V. Dhage and B.B. More, 1980. Seed borne fungi of some pearl millet cultivars. Seed Res., 8: 59-63.
Direct Link - McGee, D.C., C.L. Brandt and J.S. Burris, 1980. Seed mycoflora of soybeans relative to fungal interactions, seedling emergence and carry over of pathogens to subsequent crops. Phytopathology, 70: 615-617.
CrossRefDirect Link - Mills, J.T. and C. Frydman, 1980. Mycoflora and condition of grains from overwintered fields in Manitoba, 1977-78. Can. Plant Dis. Survey, 60: 1-7.
Direct Link - Nik, Z.W. and H. Mamat, 1982. Storage of mungbean seed (Vigna radiate (L) Wilczek) inoculated with four species of Aspergilli. Pertanika, 5: 212-218.
Direct Link - Williams, R.J. and D. McDonald, 1983. Grain molds in the tropics: Problems and importance. Annu. Rev. Phytopathol., 21: 153-178.
CrossRef